Ⅰ型鸭肝炎病毒内部核糖体进入位点的结构与功能研究
2011-05-21李传峰陈宗艳刘光清
赵 伟,李传峰,陈宗艳,刘光清*
(1.中国农业科学院上海兽医研究所国家家禽工程技术研究中心,上海 200241;2.西北农林科技大学动物医学院,陕西 杨凌 712100)
Ⅰ型鸭病毒性肝炎(Duck Virus Hepatitis,DVH)是由Ⅰ型鸭肝炎病毒(Duck hepatitis virusⅠ,DHV-Ⅰ)引起的,以肝脏出血、肿大和高死亡率为特征的急性接触性传染病。该病主要感染5周龄内的雏鸭,是对养鸭业威胁较大的传染病之一。DVH最早由美国学者Levine报道并分离病原[1]。随后,一些欧洲和亚洲国家和地区也相继报道该病疫情。1958年,中国首次报道DVH疫情,但是直到1984年才完成病原的鉴定[2]。目前该病已遍及中国大部分鸭养殖地区。
根据DHV-Ⅰ的形态学特征,国际分类委员会将其化归于小RNA病毒科的未定属,并于2006年完成对其基因组的测定[3-4]。研究表明,DHV-Ⅰ基因组为单股正链RNA分子,全长约7690 nt,由5'和3'端非编码区(UTR)和一个开放阅读框(ORF)组成,推测DHV-Ⅰ的表达可能是首先翻译出一个多聚蛋白前体,再经自身编码的蛋白酶裂解形成VP1、VP3、VP0、2A1、2A2、2B、2C、3A、3B、3C和3D等成熟蛋白[4-5]。但迄今尚无试验支持该预测结果,而关于DHV-Ⅰ的表达和调控机制的研究也尚属空白。本研究在对DHV-Ⅰ5'UTR进行序列分析的基础上,利用基因重组及定点突变等方法对5'UTR的结构和功能进行分析和研究,为深入开展DHV-Ⅰ的表达和调控机制奠定基础。
1 材料和方法
1.1 重组质粒和细胞 重组质粒pIRES-RED-eGFP、pGEM-CAT-LUC、pGEM-CAT-EMCV-LUC以及含有DHV-Ⅰ全长cDNA的重组质粒pBlDHV均由本实验室构建并保存;BHK-21细胞购自中国科学院典型培养物保藏委员会细胞库。
1.2 主要试剂 Lipofectamine购自Invitrogen公司;TNT®Quick Coupled Transcription/Translation Systems和Transcend Non-Radioactive Translation Detection Systems试剂盒购自Promega公司;鼠源eGFP抗体和HRP标记的羊抗鼠抗体均购自上海威奥生物科技有限公司;定点突变试剂盒购自北京全式金生物技术公司。
1.3 重组质粒的构建 根据DHV-Ⅰ序列(EF382778)设计扩增的引物:5'UTR+和5'UTR-以及扩增DHV-Ⅰ内部核糖体进入位点(IRES)的引物:IRES+和IRES-(表1)。以重组质粒pBlDHV为模板,扩增DHV-Ⅰ5'UTR和360 nt~620 nt片段,并分别克隆入重组质粒pIRES-RED-eGFP中,获得重组质粒pIRES-RED-DHV5'UTR-eGFP和pIRES-RED-5'UTR 260nt-eGFP。
1.4 定点突变及引物设计 以重组质粒pIRESRED-5'UTR260-eGFP为模板,按照定点突变试剂盒说明书将DHV 5'UTR 570 nt~573 nt的GATA突变为AAAA,447 nt~449 nt的TGT突变为CCC,581 nt~582 nt的GG突变为CC。获得的定点突变质粒分别命名为:pIRES-IIIem-eGFP、pIRES-SL1m-eGFP、pIRES-SL2m-eGFP、pIRES-SL1R-eGFP 和 pIRESSL2R-eGFP。针对SL-1和SL-2突变设计4条定点突变引物(表 1)。
1.5 BHK-21细胞转染及western blot检测绿色荧光蛋白(eGFP)的表达 采用常规方法将重组质粒pIRES-RED-DHV5'UTR-eGFP、pIRES-RED-eGFP和pIRES-RED-5'UTR260nt-eGFP共转染BHK-21细胞,同时设未转染质粒的BHK-21细胞对照组,24 h后荧光显微镜观察。取转染的细胞培养物,采用western blot方法检测eGFP的表达情况。
1.6 定点突变对IRES功能影响的分析
1.6.1 细胞转染试验 分别将重组质粒pIRES-Ⅲem-eGFP、pIRES-SL1m-eGFP、pIRES-SL2m-eGFP、pIRES-SL1R-eGFP和pIRES-SL2R-eGFP转染BHK-21细胞,并检测绿色荧光素酶的表达情况,分析定点突变对DHV-ⅠIRES功能的影响。
1.6.2 无细胞蛋白表达系统 以重组质粒pIRESRED-DHV5'UTR260nt-eGFP、pIRES-Ⅲem-eGFP、pIRESSL1m-eGFP、pIRES-SL2m-eGFP、pIRES-SL1R-eGFP和pIRES-SL2R-eGFP为模板,将其中的IRES和突变的IRES分别连接至原核表达载体pGEM-CAT-LUC中,分别命名为重组质粒pGEM-CAT-LUC、pGEMCAT-DHV5'UTR260nt-LUC、pGEM-CAT-Ⅲem-LUC、pGEM-CAT-SL1m-LUC、 pGEM-CAT-SL2m-LUC、pGEM-CAT-SL1R-LUC 和 pGEM-CAT-SL2R-LUC。并以其为模板,按照TNT®QuickCoupledTranscription/TranslationSystems试剂盒说明书进行体外转录和翻译,以含有猪脑心肌炎病毒(EMCV)IRES元件的重组质粒pGEM-CAT-EMCV-LUC为阳性对照,以不含IRES元件的重组质粒pGEM-CAT-LUC为阴性对照。翻译产物用化学发光方法进行分析。

表1 引物序列Table 1 Primer sequence
2 结果
2.1 重组质粒构建结果 本研究应用常规分子生物学方法构建重组质粒pIRES-RED-DHV5'UTR-eGFP和pIRES-RED-5'UTR260nt-eGFP;应用定点突变技术构建重组质粒 pIRES-Ⅲem-eGFP、pIRES-SL1meGFP、pIRES-SL2m-eGFP、pIRES-SL1R-eGFP 和pIRES-SL2R-eGFP,均经测序鉴定构建正确。
2.2 细胞转染试验结果 将重组质粒pIRES-REDDHV5'UTR-eGFP和pIRES-RED-eGFP分别转染BHK-21细胞24 h后,荧光显微镜下观察可见转染细胞均呈红色荧光(图1 A、B),在蓝色激发光下只有转染pIRES-RED-DHV5'UTR-eGFP的细胞呈现绿色荧光(图1C),而转染pIRES-RED-eGFP的细胞无绿色荧光(图1D),表明DHV-Ⅰ5'UTR可以内部启动绿色荧光蛋白的表达。

2.3 Western blot检测 将分别转染重组质粒pIRES-RED-eGFP和pIRES-RED-DHV5'UTR-eGFP的BHK-21细胞培养48 h后,收集培养物,利用绿色荧光蛋白的特异性抗体进行western blot检测,结果显示在约26 ku处存在蛋白条带,与eGFP的分子量一致,而对照细胞无可见条带(图2),进一步证明DHV 5'UTR可以启动下游绿色荧光蛋白的表达。
2.4 定点突变对DHV-ⅠIRES功能影响的分析
2.4.1 细胞内试验 将分别转染重组质粒pIRESRED-DHV5'UTR260nt-eGFP、pIRES-Ⅲem、pIRESSL1m、pIRES-SL2m、pIRES-SL1R和pIRES-SL2R的BHK-21细胞培养24 h后,荧光显微镜观察eGFP的表达情况,结果显示:转染重组质粒pIRES-Ⅲem、pIRES-SL1m和pIRES-SL2m的细胞仅能检测到红色荧光,而绿色荧光十分微弱(图3B、C、D),表明3个位点突变对IRES的翻译起始功能造成严重影响,但是将针对SL1和SL2突变构建的恢复突变体(pIRES-SL1R和pIRES-SL2R)转染细胞,检测显示均呈现绿色荧光,而后者比前者的荧光弱(图3E、F),表明上述位点是影响IRES的关键性结构域。

2.4.2 体外试验分析结果 分别以pGEM-CATLUC、pGEM-CAT-DHV5'UTR260nt-LUC、pGEMCAT-Ⅲ em-LUC、pGEM-CAT-SL1m-LUC、pGEMCAT-SL2m-LUC、pGEM-CAT-SL1R-LUC、pGEMCAT-SL2R-LUC和pGEM-CAT-EMCV-LUC为模板进行体外翻译,采用化学发光方法检测CAT和LUC蛋白的表达情况。结果显示,所有重组质粒均能够正常表达CAT蛋白,而LUC基因的表达则因其上游插入的IRES元件发生突变而表现出不同的翻译结果,如pGEM-CAT-Ⅲem-LUC、pGEM-CAT-SL1m-LUC和pGEM-CAT-SL2m-LUC 3个突变质粒均不能翻译出LUC蛋白,而回复突变的重组质粒则可翻译出 LUC蛋白(图4)。
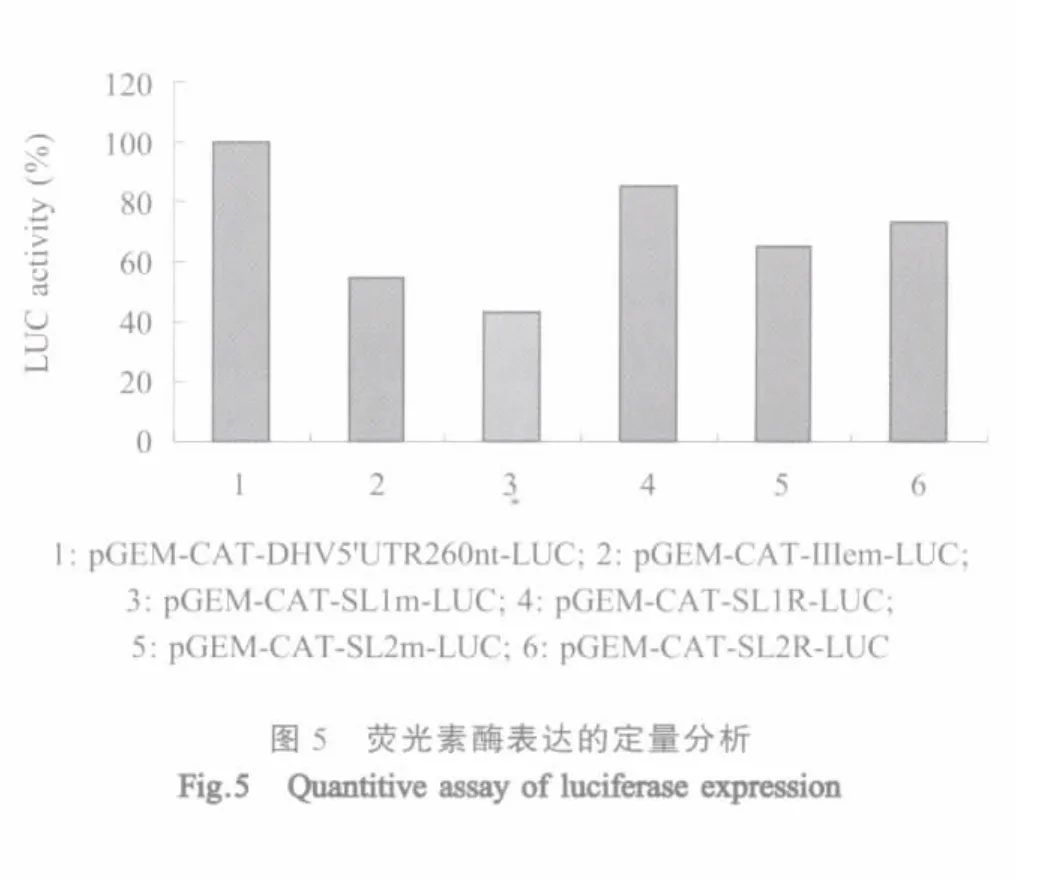
2.5 荧光素酶表达的定量分析 利用双荧光素酶检测系统对体外翻译产物中的LUC进行定量检测,结果表明:分别以重组质粒pGEM-CAT-Ⅲem-LUC、pGEM-CAT-SL1m-LUC和pGEM-CAT-SL2m-LUC为模板进行的翻译产物中,LUC的产量均下降50%左右,而以pGEM-CAT-SL1R-LUC和pGEM-CATSL2R-LUC为模板进行的翻译,LUC的产量可以基本得到恢复(图5)。


3 讨论
由于DHV基因组5'末端缺乏帽子结构,不能够采用经典的帽子结构依赖性翻译机制表达蛋白,因此研究人员推测DHV-Ⅰ可能是利用内部起始翻译的机制表达病毒的蛋白质。本研究根据预测结果,以试验证实在DHV-Ⅰ5'UTR的260 nt~620 nt处含有典型IRES元件。体内和体外试验结果均表明DHV-Ⅰ5'UTR可以启动下游报告基因(绿色荧光蛋白和荧光素酶)的表达。本研究首次以试验证明DHV-Ⅰ基因组的翻译起始方式与口蹄疫病毒相似,是一种高度依赖IRES元件的内部翻译起始机制。
目前,已经发现很多依赖IRES元件进行翻译起始的RNA病毒,如口蹄疫病毒、丙型肝炎病毒、肠道病毒和猪脑心肌炎病毒等。根据IRES的二级结构和功能,可将其分为四大类:Ⅰ类主要包括心病毒属和口疮病毒属的成员;Ⅱ类主要存在于肠道病毒属和鼻病毒属;Ⅲ类多见于肝病毒属;Ⅳ类存在于黄病毒属成员中[6-9]。通过比较和分析,我们发现DHV-ⅠIRES的二级结构与Ⅳ类IRES的结构相近。由于IRES是通过形成复杂而稳定的RNA结构在反式作用因子存在的情况下富集核糖体来启动翻译的,因此,研究IRES的结构及其与IRES功能之间的关系对于研究DHV-Ⅰ的翻译调控机制具有重要意义。预测和分析显示DHV-ⅠIRES元件中有三个保守性的二级结构(Stem1、Stem2和Ⅲe),同样存在于HCV和禽脑脊髓炎病毒的IRES元件中,该结构是决定IRES功能的关键性结构域[10-11]。本研究通过定点突变和体内、体外试验分析,证明这些结构域同样也是影响DHV-ⅠIRES的关键区域。破坏任何一个结构域,IRES起始翻译的功能均将丧失。本研究为进一步探索DHV-Ⅰ的翻译起始和调控机制奠定了基础。
[1]Levine P P,Hofstad M S.Duck disease investigation[M].Annu Rep New York State Vet Coll,Ithaca,1945,55-56.
[2]刘兆宇,程安春.雏鸭病毒性肝炎的研究进展[J].中国家禽,2002,24(10):6-9.
[3]Kim M C,Kwon Y K,Joh S J,et al.Molecular analysis of duck hepatitis virus type 1 reveals a novel lineage close to the genus Parechovirusin the family Picornaviridae[J].J Gen Virol,2006,87:3307-3316.
[4]Tseng C H,Knowles N J,Tsai H J.Molecular analysis of duck hepatitis virus type 1 indicates that it should be assigned to a new genus[J].Virus Res,2007,123:190-203.
[5]Ding C,Zhang D.Molecular analysis of duck hepatitis virus 1[J].Virology,2007,361:9-17.
[6]Palmenberg A C,Sgro J Y.Topological organization of picornaviral genomes:statistical prediction of RNA structural signals[J].Semin Virol,1997,8(3):231-241.
[7]Dorner A J,Semler B L,Jackson R J,et al.In vitrotranslation of poliovirus RNA:utilization of internal initiation sites in reticulocyte lysate[J].J Virol,1984,50(2):507-514.
[8]Glass M J,Summers D F.Identification of a trans-acting activity from liver that stimulates hepatitis A virus translationin vitro[J].Virology,1993,193(2):1047-1050.
[9]Chard L S,Kaku Y,Jones B,et al.Functional analyses of RNA structures shared between the internal ribosome entry sites of hepatitis C virus and the picornavirus porcine teschovirus 1 talfan[J].J Virol,2006,80(3):1271-1279.
[10]Pisarev A V,Chard L S,Kaku Y,et al.Functional and structural similarities between the internal ribosome entry sites of hepatitis C virus and porcine teschovirus,a picornavirus[J].J Virol,2004,78(9):4487-4497.
[11]Bakhshesh M,Groppelli E,Willcocks M M,et al.The picornavirus avian encephalomyelitis virus possesses a hepatitis C virus-like internal ribosome entry site(IRES)element[J].J Virol,2008,82:1993-2003.
