广东地区汉族人群TLR1基因多态性的测序研究*
2011-03-17肖文娟刘泽寰
张 斌, 肖 林, 肖文娟, 刘泽寰△
(1暨南大学生命与健康工程研究院,广东广州510632;2中山大学附属第五医院,广东珠海519000)
Toll样受体1(toll-like receptor 1,TLR1)基因编码TLR1蛋白,因其与果蝇的Toll蛋白和人类的白细胞介素1(interleukin-1,IL-1)受体的同源性而得到确认。该基因位于人基因组4p14,长度为8 537 bp,有4个外显子区(exon),分别位于 -5 960~-5 923、-5 551~-5 475、-2 117~-2 026和-67~2 576(以翻译起始密码ATG的A为+1)。外显子1、2、3连同外显子4的前67 bp是5'非翻译区(untranslated region,UTR),外显子4尾部的216 bp是3'UTR。
TLR1蛋白是一种1型跨膜蛋白,由胞外区、跨膜区和胞内区组成。该蛋白含有786个氨基酸,前22个氨基酸为信号肽(signal peptide,SP),23~582的氨基酸序列组成其胞外区,583~605是跨膜区,606~786为其胞内区。其中胞内区包含有 TIR (Toll/interleukin-1 receptor)结构域,而胞外区则具有富含亮氨酸的重复序列(leucine-rich repeat,LRR)[1]。LRR序列一般由24个氨基酸组成,其特征是亮氨酸间隔分布于几个固定位点,此结构有利于促进蛋白质间的相互黏附,故可用来识别病原体或其产物;胞内区胞内段由Toll同源结构域(Toll homology domain,THD)和分子羧基端长短不同的短尾肽组成,与人IL-1受体的胞内区具有同源性。在TLR蛋白家族中,TLR1和LTR6存在高度的同源性,分别能与TLR2形成异二聚体,主要功能是与TLR2结合识别革兰氏阳性菌及革兰氏阳性菌其细胞壁成分如脂肽、脂蛋白等。然而TLR1/2和TLR1/6在对微生物的识别上,均有各自的专一性,例如二酰基脂肽和三酰基脂肽[2]。
近年来涉及到TLR1基因多态性导致免疫系统应答和疾病易感性的证据不断出现。例如位于TLR1基因编码的胞外区域11号LRR区域的非同义突变单核苷酸多态性(single nucleotide polymorphism,SNP)位点+944 C/T(Pro315Leu,rs5743613)就与胞外受体的配体识别有关。尽管没有与这个SNP位点直接关联的疾病被报道,但可以肯定的是它对识别细菌细胞壁成分的天然免疫防卫产生着负面的易感性影响[3]。而位于TLR1基因编码的跨膜区域的非同义突变SNP位点+1 805 G/T(Ser602Ile,rs5743618)也被认定是一个与 TLR1功能相关的SNP位点。这个位点的突变改变了细菌脂肽识别,并潜在地影响到天然免疫应答机制和一些相关疾病的易感性[4]。
TLR1在天然免疫应答机制上对于识别外源病原体具有重要作用,但国内对TLR1基因遗传多态性研究几乎处于空白状态,所以对中国汉族人TLR1基因单核苷酸多态性的研究,探究分子水平上的个体及群体差异性是十分必要的。针对这种情况,本研究采用高分辨率的PCR直接测序的方式,随机筛查50例广东地区汉族人TLR1基因的调控区、5'和3'非翻译区以及4个外显子区的功能性多态性位点,初步建立起广东汉族人TLR1基因的功能性多态图谱。这些工作,将会为进一步研究该基因多态性与中国人常见其它自身免疫疾病的相关性研究打下良好基础。
材料和方法
1 样本收集和基因组DNA提取
从采集到的广东地区汉族无亲缘关系、健康个体的抗凝外周血样本中随机抽取50例作为研究样本,采血对象年龄在19~48岁间,男女性别比例为1∶1.2。用E.Z.N.A.®血液DNA抽提试剂盒(Omega)提取基因组DNA。本研究获得暨南大学附属第二医院医学伦理委员会批准,所有采血对象均签署知情同意书。
2 目的片段的扩增和测序
根据GenBank公布的人类TLR1基因信息(Accession:NC_000004;Reglon:38797876~38806412; Versoon:NC_000004.11;GI:224589816),用Oligo 6.0软件(Molecular Biology Insight Inc.)设计引物,对随机抽取的50例广东汉族人TLR1基因的启动子区、5'和3'非翻译区、4个外显子区进行PCR扩增,扩增引物见表1。PCR扩增条件为:94℃变性5 min;之后35个循环:95℃变性30 s,退火30 s(各片段退火温度见表1),72℃延伸90 s;最后72℃延伸10 min。PCR产物用3S Spin PCR产物纯化试剂盒(上海申能博彩公司)纯化后,用ABI PRISM 377自动测序仪测序(英韦创津公司),测序引物与扩增引物相同,每个扩增片段均进行正、反向2次测序。
3 统计学处理
参照GenBank参考序列,用 ClustalX 1.81软件[5]比对测序结果获得TLR1基因多态性位点信息及其频率分布;用SPSS 13.0软件进行卡方检验以验证Hardy-Weinberg平衡;用DnaSP 5.10软件[6]计算Tajima's D、Fu&Li's D值和频谱分析;用软件Haploview 4.1[7]进行连锁不平衡分析。

表1 PCR扩增及测序引物Table 1 .Primers for PCR amplication and sequencing
结果
通过对50例广东汉族人TLR1基因启动子区、5'和3'非翻译区以及4个外显子区等功能性区域进行分段扩增,分别得到了与5种预期目的片段长度一致的扩增结果,见图1。在此基础上针对这些扩增片段构成的总长为5 813 bp的区域进行测序,共发现17个SNPs位点和2个插入/缺失(insertions and deletions)多态,见表2,分别为:-6 876 C/T、-6 847 A/G、-6 737 A/T、-6 399 C/T、-6 375 C/T、-6 010 C/T、-5 531 A/G、-5 507 A/G、-5 490 C/ G、-2076C/T、+130C/T、+743A/G、+936C/T、 +1 378 A/G、+1 482 A/G、+1 518 A/G、+1 937 C/T、-6 912 C/TA和-5 984-/CT。所有多态性位点均符合Hardy-Weinberg平衡(P>0.05)。其中,前8个位于启动子调控区,中间4个位于5'非翻译区,后7个位于编码区。对比美国国立生物信息中心(National Center for Biotechnology Information,NCBI)db-SNP数据库的已有数据发现,其中SNPs位点+936 C/T和+1 378 A/G属于首次发现,见图2,且位于编码区的新SNP位点+1 378 A/G为非同义突变位点,能导致460位丝氨酸(Ser)残基替换为甘氨酸(Gly)残基,并且这个氨基酸残基的替换处于TLR1胞外区的LRR结构域中,从而有可能影响蛋白的识别功能。
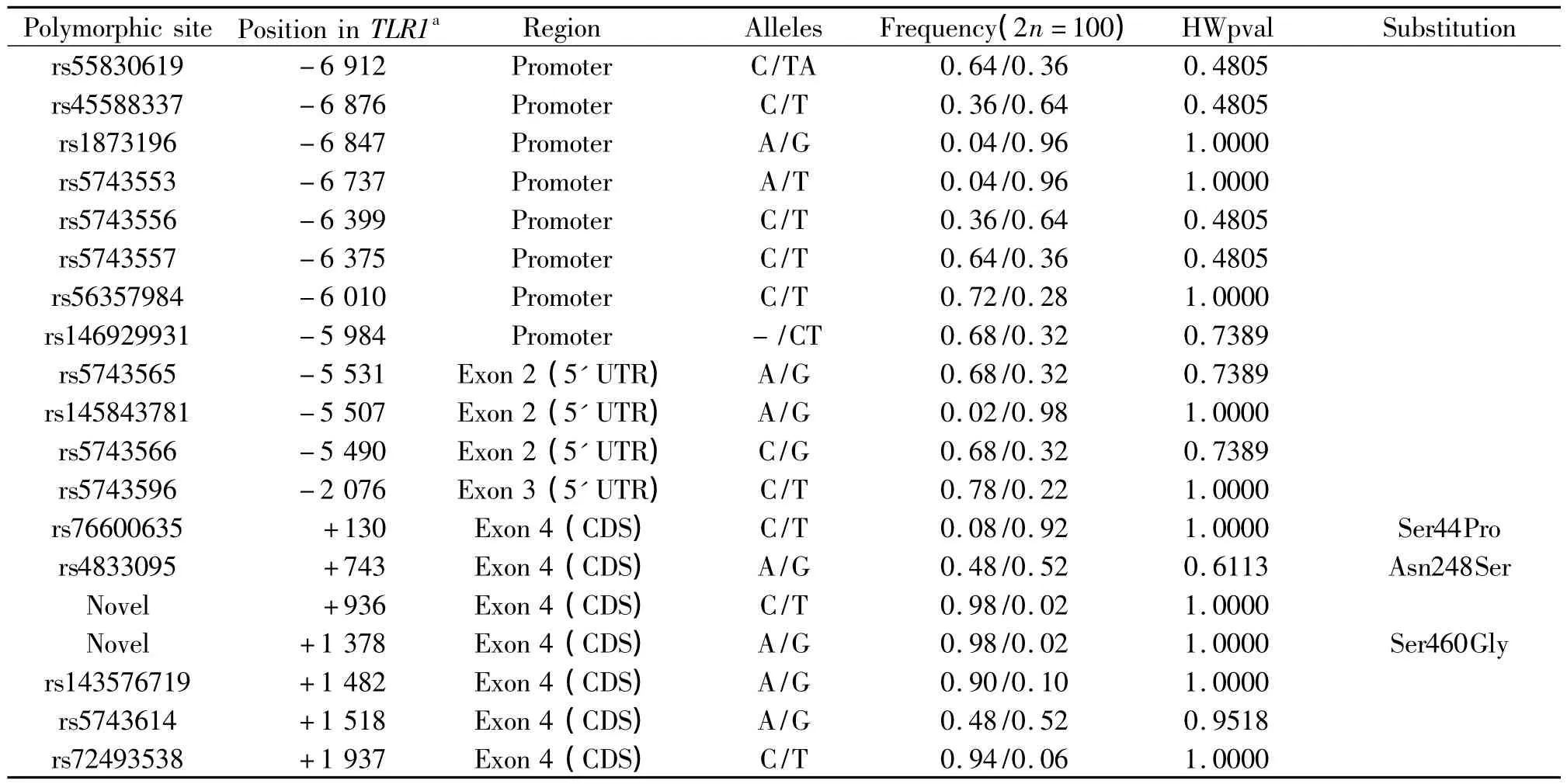
表2 广东汉族人群TLR1基因功能性多态性位点的相关信息Table 2 .Characterization of functional genetic polymorphisms in TLR1 locus in Cantonese Han population

Figure 1.Electrophoresis analysis of amplified DNA fragments from TLR1 gene.A:including promoter,exon 1,intron 1 and exon 2 region(-7 038~-5 286);B:exon 3(-2 339~-1 959);C:partial exon 4(-281~1 100);D:partial exon 4 (731~2 386);E:partial exon 4(2 148~2 789)M:DNA marker.1:PCR products.图1 TLR1基因扩增片段电泳图

Figure 2.Two novel SNPs found in TLR1 gene.A:promoter(+936,C/T genotype);B:promoter(+1378,A/G genotype).The arrows represent mutation sites.图2 TLR1基因中新发现的2个SNPs位点
将广东汉族人群TLR1基因的多态性位点频率数据与欧洲人群、日本人群和非洲人群进行了比较,见表3。广东汉族人和欧洲人在绝大多数多态性位点的频率分布上差异显著;与日本人相比,在rs55830619、rs45588337、 rs5743556、 rs5743557、rs56357984、rs5743565、rs5743566、rs5743596 及rs4833095等的频率分布上也有较大的差异;而非洲人群则由于数据不全,因此可比性不强,但在仅有的多态 性 位 点 rs1873196、rs5743553、rs5743557、rs5743565、rs5743596及rs4833095的频率分布上差别还是非常显著的,2个在广东汉族人中低频的位点在非洲人群中却频率较高,而3个在广东汉族人中呈现高频的位点在非洲人群中却完全观察不到。至于位点rs146929931、rs145843781及rs143576719则在其他人群中的频率至今尚未见报道。

表3 广东汉族人群TLR1基因功能性多态性位点的等位基因频率与其他人群的比较Table 3 .Functional genetic polymorphisms and their minor allele frequencies of TLR1 in different populations
在以上测序结果和SNPs频率分布数据的基础上,本研究还分析了TLR1基因全部功能区的核苷酸多样性水平(nucleotide diversity,π),为1.5×10-3,高于人类基因组的平均水平(0.8×10-3)[8]。中性检验结果显示,TLR1基因功能区的Tajima's D值和Fu&Li's D值分别为1.38062和1.73756,前者不显著(P>0.1),后者有较强的显著性(P<0.02),说明该基因一定程度地背离了中性进化,很可能受到平衡选择的作用。频谱分析也发现,TLR1基因多态性位点的频谱分布不符合中性进化,见图3,中高频突变过多,很可能是平衡选择造成的。

Figure 3.The frequency spectrum analysis of the functional regions of TLR1 gene.The curve represents expected values.Histogram represents the actual observed values.图3 TLR1基因功能区的频谱分析
用Haploview 4.1对TLR1基因19个多态性位点进行连锁不平衡分析发现,多态性位点-6 912 C/ TA、-6 876 C/T、-6 399 C/T、-6 375 C/T之间(Block 1),-6 847 A/G和-6 737 A/T之间,以及-5 984-/CT、-5 531 A/G和-5 490 C/G之间(Block 2)完全连锁(r2=1);位点-6 010 C/T和Block 2之间,+743 A/G和+1 518 A/G之间,以及Block 1和Block 2之间紧密连锁(r2>0.8),见图4。
讨论
本研究新发现的SNP位点+1 378 A/G为非同义突变位点,能导致460位Ser残基替换为Gly残基,并且这个氨基酸残基的替换位于TLR1胞外区的LRR结构域中。有报道指出,LRR序列被认为与病原体模式分子的识别有关[9],而处于LRR序列中的非同义SNP就很有可能影响到蛋白对于病原体的识别。比如处于TLR1基因LRR序列上的非同义SNP位点Pro315Leu(rs5743613)就与TLR1蛋白识别病原体的免疫防卫机制有关[10]。
人类先天免疫相关基因的多态性水平通常较低[11]。但在本研究中却发现,广东汉族人群TLR1基因功能区的核苷酸多样性明显高于人类基因组平均水平。这说明,广东汉族人群TLR1基因在进化过程中可能受到不寻常因素的影响。通过Tajima's、Fu&Li's检验以及频谱分析我们发现,TLR1基因多态性明显背离中性进化理论。为此,我们进一步将TLR1功能区序列分为调控序列和外显子序列两部分来进行中性检验,发现外显子区没有背离中性进化,而是调控区可能受到了平衡选择的作用,Tajima's D值为2.09685(P<0.05)。这说明TLR1基因的调控区可能具有重要的生物学功能。当然,这种结果还有可能由搭便车效应、群体瓶颈效应或者群体结构改变造成。

Figure 4.Linkage disequilibrium(LD)plot of the functional polymorphisms in TLR1 gene.The r2(×100)values were depicted in the diamonds.White:r2=0;grey:0<r2<1;black:r2=1.图4 广东汉族人群TLR1基因功能性多态性位点的连锁不平衡分析
人类基因多态性在不同种群、地区间的分布具有明显差异。在本研究中,广东汉族人与欧洲人、非洲人在TLR1基因多态性频率分布上差异明显。比如,在广东汉族人样本中,没有发现在欧洲和非洲人群中高频且与疾病易感的非同义SNPs位点+1 805 G/T(Ile602Ser,rs5743618),有证据显示Ile602Ser这个位于TLR1跨膜区的SNP位点能调节细菌脂肽的识别,从而影响着天然免疫应答和对一些病原体的临床易感性[12]。还有报道指出,在节段性回肠炎患者中,602Ser突变的纯合子个体是最早表现出回肠末端炎症反应的[13]。同样,前面提到的与TLR1蛋白识别病原体的免疫防卫机制有关的Pro315Leu也未在广东汉族人样本中检测到。这些均可能是选择压力、遗传漂变与迁徙等因素导致不同地区人群的等位基因频率发生分化的结果。
本研究发现的19个多态性位点如进一步用于TLR1基因的疾病相关性研究或临床检验,可以根据连锁不平衡分析的结果进行精简,筛选出较有代表性tSNP(tag SNP)后再进行检验。其中要注意的是,本来应从内部完全连锁的Block 1、Block 2、-6 847 A/G和-6 737 A/T之间各选择一个tSNP位点,但由于 Block 1和 Block 2之间以及 -6 010 C/T (rs56357984)和Block 2之间紧密连锁,因此可合并选择一个tSNP,原则上应在调控区的启动子和转录调控位点附近选择,因为之前的分析已指出,调控区受到平衡选择,相对于选择中性的外显子2可能在功能上更加重要;另外,紧密连锁的 +743 A/G (rs4833095)和+1 518 A/G(rs5743614)之间也可以选择一个作为tSNP,应首选引起氨基酸改变的非同义突变rs4833095;此外,在不连锁的其余位点中,处于编码区的同义突变 +936 C/T、+1 482 A/G (rs143576719)以及+1 937 C/T(rs72493538)没有太大的研究价值,建议可以省略。
由于TLR1在天然免疫中具有重要的作用,其基因多态性与疾病的易感关系已越来越成为研究热点。但是,由于人群之间等位基因频率分布的差异较大,直接选择其他人种、民族人群中已发现的高风险位点在中华民族人群中进行研究是不妥当的。为此,本研究首次系统、全面地建立了广东汉族人群TLR1基因功能区的多态性图谱,它将为今后在汉族人群中开展TLR1蛋白功能、疾病易感性、致病机理乃至人群进化研究提供重要基础。
[1] Xiao WJ,Li N,Gong YX,et al.Single nucleotide polymorphisms of TLR4 locus in Chinese Cantonese population[J].中国病理生理杂志,2010,26(6):1193-1197.
[2] 伍模鑫,肖文娟,刘泽寰.广东汉族人群TLR2基因的多态性研究[J].中国病理生理杂志,2010,26(12): 2373-2377.
[3] Omueti KO,Mazur DJ,Thompson KS,et al.The polymorphism P315L of human TLR1 impairs innate immune sensing of microbial cell wall components[J].J Immunol,2007,178(10):6387-6394.
[4] Hawn TR,Misch EA,Dunstan SJ,et al.A common human TLR1 polymorphism regulates the innate immune response to lipopeptides[J].Eur J Immunol,2007,37 (8):2280-2289.
[5] Thompson JD,Gibson TJ,Plewniak F,et al.The CLUSTAL-X windows interface:flexible strategies for multiple sequence alignment aided by quality analysis tools[J].Nucleic Acids Res,1997,25(24):4876-4882.
[6] Librado P,Rozas J.DnaSP v5:a software for comprehensive analysis of DNA polymorphism data[J].Bioinformatics,2009,25(11):1451-1452.
[7] Barrett JC,Fry B,Maller J,et al.Haploview:analysis and visualization of LD and haplotype maps[J].Bioinformatics,2005,21(2):263-265.
[8] Reich DE,Schaffner SF,Daly MJ,et al.Human genome sequence variation and the influence of gene history,mutation and recombination[J].Nat Genet,2002,32(1): 135-142.
[9] Fitzgerald KA,Chen ZJ.Sorting out Toll signals[J].Cell,2006,125(5):834-836.
[10] Omueti KO,Mazur DJ,Thompson KS,et al.The polymorphism P315L of human Toll-like receptor 1 impairs innate immune sensing of microbial cell wall components[J].J Immunol,2007,178(10):6387-6394.
[11] Ferrer A,Bosch E,Sikora M,et al.The evolution of parasite recognition genes in the innate immune system:purifying selection on drosophila melanogaster peptidoglycan recognition proteins[J].J Mol Evol,2003,57(5):598-605.
[12] Schumann RR,Tapping RI.Genomic variants of TLR1-it takes(TLR-)two to tango[J].Eur J Immunol,2007,37(8):2059-2062.
[13] Pierik M,Joossens S,van Steen K,et al.Toll-like receptor-1,-2,and-6 polymorphisms influence disease extension in inflammatory bowel diseases[J].Inflamm Bowel Dis,2006,12(1):1-8.
