不同迁移方式与不同学习“方式”对知觉运动序列学习的影响——一项ERP研究
2011-01-03胡伟,吕勇
胡 伟,吕 勇
●专题研究
不同迁移方式与不同学习“方式”对知觉运动序列学习的影响
——一项ERP研究
胡 伟,吕 勇
拟采用迁移范式,以事件相关电位(ERP)成分N2、P3作为主要指标,结合不同迁移方式和不同学习“方式”探讨学习者的知识表征方式的差异。结果发现:内隐组和外显组被试在习得了一定的序列知识后,相对于标准序列的反应时,效应器迁移序列的反应时明显变慢;在脑电数据中,以N2的平均波幅作为指标,不管是内隐组还是外显组,标准序列与刺激迁移序列不存在区别,而与效应器迁移序列却差异显著。对于P3的平均波幅的分析得出:在内隐组,标准序列与效应器迁移序列不存在区别;而在外显组,两者却差异显著。该结果说明:效应器水平的编码机制是参与序列知识的学习的;且相对于知觉编码,不管以何种“方式”学习,效应器编码起的作用都要更大。
知觉运动序列;迁移;知识表征方式;事件相关电位
1 问题提出
人类最不平凡的能力之一便是可以学会复杂的顺序规则。人可以学会弹钢琴、打网球、织毛衣、骑自行车说话、书写等,这些都涉及序列学习(sequence learning)能力。而序列学习又可以以不同的方式进行,比如知觉序列学习(例如只通过听或看进行)、动作序列学习(例如只通过手或脚等动作效应器进行)或知觉动作序列学习(例如边看边进行动作)。
上述序列学习的学习方式涉及了序列学习知识表征问题。所谓知识表征,就是学习者在学习过程中到底学到了什么;而知识表征方式,则涉及了学习者在知识获得过程中使用的编码方式。本研究首先则试图探讨知觉运动序列学习(percepto-motor sequence learning)过程中究竟是依存于效应器(effectordependent)还是独立于效应器(effector-independent)。对于知觉动作序列,最明显的序列结构便是刺激物按次序呈现的刺激序列以及效应器做出反应的动作序列。那么,关于序列结构的知识到底是由做出动作的效应器编码的,还是完全与效应器编码系统无关呢?又或者,有没有可能该表征方式是由知觉和动作联合编码的呢?Raibert的研究表明,动作序列的编码是完全独立于效应器的[1]。因为他发现一个人的笔迹,不管他是用手,前臂,手肘,甚至脚或嘴写出来,通过笔记学鉴定结果都是相似的。另外,Margolin以及后来的Hillis和Caramazza对于神经病患者的神经科学研究也表明,编码文字书写顺序的脑区与编码文字的口头拼读的顺序的脑区存在重合[2-3]。Cohen等也对这一问题展开了研究,他们让被试对在不同位置出现的刺激用3个不同的手指反应,在接下来的阶段,转换效应器,让被试只用一个手指反应[4]。结果发现尽管效应器改变,在前一阶段习得的知识仍然发生了迁移。Remillard等人借助双刺激的研究范式,发现无需借助动作反应也可习得刺激序列[5]。“双刺激”范式包括两种反应方式,一种为“反应相关特征”,即被试需要根据刺激做出相应反应;另一种为“反应无关特征”,即被试无需根据该刺激选择相应反应。结果发现,被试在两种情况下都习得了序列规则。作者据此认为不通过动作效应器,单凭学习视觉刺激便能习得序列规则。然后,Keele等并不支持这种观点,他们将这种迁移应用到了不同的效应器系统[6]。研究者让被试在学习阶段按键,而在转换阶段用声音反应,结果发现虽然迁移效应很小但仍然存在,这似乎说明学到的并不是一系列特定的效应器的反应。然而在不同的效应器迁移过程中出现的反应时增加似乎又表明不同序列结构知识的编码对于特定的效应器系统存在一定的依赖性[7-8];Bapi等又考察了在不迁移效应器,只变换刺激的情况,发现对于序列知识的迁移同样发生[9]。这就说明序列学习中可能还包含了非效应器编码的成分,比如对刺激的知觉编码。
另一方面,不同的学习“方式”下,即内隐学习和外显学习中,序列学习者的知识表征方式是否存在不同也是本研究着力于解决的一个问题。自从序列学习被证明可以以内隐的方式进行[10-17],外显序列学习和内隐序列学习的知识表征方式差异便成为该领域研究的重点问题之一[18-21]。Eimer等结合ERP技术使用序列反应时范式的变式来研究序列学习的知识表征方式,并且在不同的学习方式下(内隐和外显)进行研究[18]。首先,让被试学习一个由4个大写字母组成的有规律的序列,序列中的标准字母偶尔被偏差字母代替。偏差字母要求被试用和标准字母不一样的手反应。其次,通过外显测试将被试分为内隐被试和外显被试。试验结果表明,偏差字母在刺激后的240~340 ms引发了一个较标准字母的波幅更大的负波(N2b)。这个成分后面紧跟着一个正成分(P3b),在该成分上偏差字母效应同样存在。这种现象只存在于外显被试中,在内隐被试中则不存在。作者由此得出结论,N2b成分可能反映了被试获得的关于规则序列有意识知识的量。在Russeler等人的系列电生理研究中,他们同样使用了与Eimer相似的范式[19-20]。只不过他们将偏差字母区分为知觉偏差(只违背刺激序列不违背反应序列)和动觉偏差(既违背刺激序列又违背反应序列)。结果发现在外显组知觉偏差和动觉偏差较之标准字母都引发了更大的N2b波幅,比较P3b的波幅时,只有动觉偏差出现了这种情况。内隐组并未发现N2b和P3b成分上的差异。反应时数据发现外显组知觉偏差和动觉偏差较之标准字母的增加,然而内隐组只出现了动觉偏差反应时的增长。作者据此得出外显和内隐学习涉及了不同的知识表征方式。内隐学习者主要习得了反应—反应之间的联系,而外显学习者则可能习得了刺激—刺激、刺激—反应或反应—刺激之间的联系。相同序列的知识表征方式在不同学习类型下存在显著差别,外显的学习方式序列知识倾向于以知觉和动觉同时表征,而内隐的学习方式序列知识则倾向于动觉表征。同样的序列知识出现了不同的表征方式,这同样暗示着序列知识的加工方式并不是一个单一的过程。
由上述研究可以看出,对于序列学习领域的研究已越来越倾向于使用具有高时间分辨率的事件相关电位技术,通过测量与序列学习相关的特定脑电成分而进一步探讨序列学习的知识表征方式。本试验将在人工语法范式中被广泛应用的迁移任务应用于序列反应时任务之中[13,22-23],结合不同的迁移方式和不同的学习“方式”探讨学习者的知识表征方式是否存在不同。此种设计的初衷源于在以前人工语法的研究中,不同字母串间的整体迁移会随着人工语法的表现形式或反应形式的变化导致内隐学习量的减少[24-27];拟采用事件相关电位技术(ERPs),通过ERPs的高时间分辨率,直接探索序列学习知识表征方式的脑机制。总结起来,当前试验是一个2因素混合试验设计,分别为序列类型因素(3水平:标准序列,刺激迁移序列,效应器迁移序列)、学习“方式”因素(2水平:内隐组,外显组)。
2 研究对象与方法
2.1 被 试
38名本科生作为有偿被试参加了本试验。他们被随机地分配到“告知组”和“不告知组”。告知组有19名被试(10男9女),其余为不告知组被试(9男10女)。外显知识测验剔除了14名被试,另有3名被试的数据因脑电伪迹太多而被剔除,故最终有效被试21名,内隐组10名,6男4女,年龄为22.3±0.8岁;外显组11名,5男6女,年龄为22.7±0.5岁。所有被试皆无精神或神经损伤史,均为右利手,视力正常或矫正至正常,无参加序列学习试验的经历。
2.2 材 料
试验材料为8个大写字母组成的序列,分别为D、L、N、R、T、V、X、Z。呈现于21英寸CRT显示器(分辨率1 024×768,刷新率75 Hz)上。通过美国NeuroScan公司的Stim2软件控制刺激呈现,字母呈现于屏幕中央,为白色,背景为黑色。屏幕距被试双目约1 m,字母的视角为0.58°×0.28°。
2.3 试验程序
试验前,将被试随机分配到“告知组”和“不告知组”。告知组告知被试字母序列存在规律,发现规律可以提高任务成绩。不告知组只告知被试为简单的按键反应试验。两组被试都被要求又快又准地对字母做出反应,同时都必须完成指导语中要求的第二任务。
被试坐于隔音且亮度适中的电磁屏蔽室内,对屏幕上出现的相应字母按键。手指与字母的对应规则如下:左手食指对应字母D、T;左手中指对应字母L、V;右手食指对应字母R,Z;右手中指对应字母N、X。按键的手在被试间做了平衡。当字母在屏幕上出现时,被试使用对应的手指按键,字母消失,500 ms后出现下一个字母。如果被试不作反应,字母将保留在屏幕上,直到被试反应为止(见图1)。

图1 试验流程图Fig.1 Experiment procedure
试验包括练习和正式试验。练习材料由D、L、N、R、T、V、X、Z随机排列而成,72个字母为一组段。练习至被试一组段中错误数少于6个字母,平均反应时低于1 500 ms时即开始正式试验。正式试验共19个组段(block),包括标准序列和迁移序列。其中第1-6、8-10、12-14、16-18组段为标准序列,其余为迁移序列。标准序列的字母序列顺序为:NLRDLDNRLNDR。每个组段中序列重复6次。第7、19组段为刺激迁移序列,字母序列为XVZTVTXZVXTZ。刺激迁移序列是将标准序列中的N、L、R、D替换为X、V、Z、T,但对应的按键手指不变。其改变了刺激序列,即改变了字母,但效应器的动作序列即按键的手指的顺序没有被破坏。该组段中字母序列重复3次。第11、15组段为效应器迁移序列,该序列的字母顺序与标准序列相同,只是在该序列组段开始之前,以指导语要求被试以对侧手的相同手指对字母作反应,即应该用左手食指反应的,换用右手食指反应,依此类推。该序列改变了反应的效应器,即改变了按键的手,但对外界刺激本身并没有被破坏。该组段中字母序列重复3次。标准序列和迁移序列的每一组段中,每一重复的字母序列的起始字母都是随机选取的,每一个字母都不会连续出现。
在做按键反应的同时,要求被试完成第二任务,即对字母序列中的“L”计数。试验间歇,在指导语的提示下,将从试验开始到指导语出现为止字母“L”出现的总次数记录在纸上。记录之后,开始重新计数,直到该指导语再次出现,再次记录。设置计数字母序列中的“L”的目的是尽量使不告知组被试不能对字母序列规律的进行有意识的探索,以保证序列学习无意识的进行。
序列学习结束后,对被试的外显序列知识进行评估。根据过程分离的方法将生成任务分成包含任务和排除任务。两种任务都是填空任务,包含任务为给出标准序列中任意两个相临字母,然后要求被试填出下一个字母,每填对1个记1分;排除任务则要求填写两个相临字母后面那个字母之外的标准序列中余下的那个字母,如果填对记1分。被试在计算机上完成这两种任务,任务的先后顺序在被试间做了平衡。
2.4 脑电记录
被试佩戴Quick-cap64导Ag/AgCl脑电帽,导联方法采用国际10-20系统,前额发际下1 cm处接地,以左右乳突的平均值为参考,在眼眶周围记录水平眼电(HEOG)和垂直眼电(VEOG)。采样率500 Hz,模拟滤波带通为0.01~100 Hz,50 Hz陷波处理,放大150 000倍,精确度0.084 μV/LSB,头皮电阻小于5 kΩ。
3 数据分析及结果
3.1 内隐被试和外显被试的鉴别
根据被试在生成任务中的表现,对内隐被试和外显被试进行区分。在告知组,被试在包含任务中正确的次数大于排除任务中正确的次数,且大于填空总次数的33.3%为外显被试,其余被试被剔除。在不告知组,被试在生成任务的包含任务中正确的次数小于或等于排除任务中正确的次数,或小于填空总次数的33.3%为内隐被试,其余被试被剔除。24名被试被保留下来,外显组和内隐组各12名。
3.2 行为数据
将各个组段中的错误、重复反应的数据剔除,每个组段中正负3个标准差之外的数据也被剔除。所有被试在计数任务中,都没有偏离正确的“L”的个数的5%。外显组和内隐组在各个组段反应时情况见图2。

图2 外显组和内隐组在各个组段的反应时Fig.2 Reaction time of explicit and implicit group in every block
分别对内隐组和外显组的第1和第6组段的反应时数据进行的配对样本T检验,皆发现显著性差异(内隐组:t(9)=5.253,P<0.01;外显组:t(10)=3.367,P<0.01),这表明两组被试在第6组段(即迁移组段之前)都习得了序列知识。
使用两因素重复测量方差分析检验不同序列间的差异,以及这种差异在两组被试中是否存在不同。主因素为序列类型(迁移方式)(3水平:标准序列,刺激迁移序列,效应器迁移序列)和学习方式类型(2水平:内隐组,外显组),标准序列的反应时由第6、10、14、18组段中的反应时平均获得。统计分析发现:序列类型主效应显著(F(2,38)=60.761,P<0.01),学习方式类型主效应不显著(F(1,19)=0.703,P>0.05),序列类型与学习方式类型的交互作用显著(F(2,38)=6.763,P<0.05)。从第6组段开始到试验结束,内隐组对标准序列的反应显著快于对刺激迁移序列的反应((647±176)ms,(896±228)ms,P<0.05),也显著地快于对效应器迁移序列的反应((1 169±315)ms,P<0.01),刺激迁移序列与效应器迁移序列的反应时也存在显著差异(P<0.05);在外显组中,标准序列的反应时显著地快于刺激迁移序列((562±107)ms,(979±219)ms,P<0.01),也显著地快于效应器迁移序列((967±212)ms,P<0.01),刺激迁移序列与效应器迁移序列并无显著差别(P>0.05)。统计分析还发现,两组被试对3种序列反应的正确率均不存在显著差别(P>0.05)。
3.3 脑电数据
脑电数据统计分析因素包括序列类型因素(3水平:标准序列,刺激迁移序列,效应器迁移序列)、学习类型因素(2水平:内隐组,外显组),脑区位置共两因素,分别为前后脑区位置(4水平)和左右脑区位置(3水平)。
脑电数据经过与行为数据的融合,去眼电之后,以刺激开始前的200 ms和刺激呈现后的800 ms为起止时间进行分段,以刺激前200 ms作为标准进行基线校正。删除反应错误,反应重复,反应时快于100 ms或慢于1 500 ms的脑电分段。波幅在± 100 μV之外脑电的分段被认为存在伪迹而剔除。内隐组和外显组共有3名(内隐组2名,外显组1名)被试因为脑电伪迹过多而被剔除,其行为数据亦未被统计。
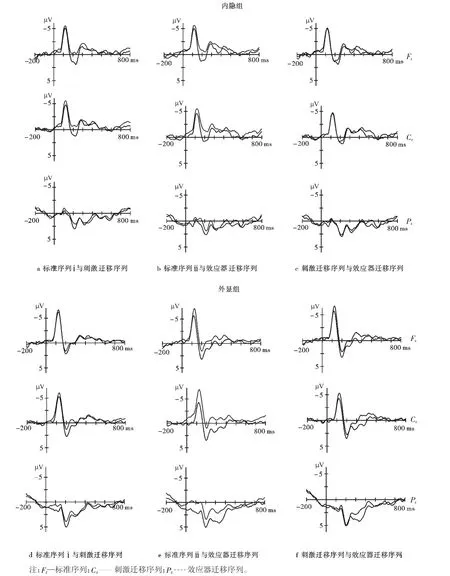
图3 内隐组和外显组被试3种不同序列在Fz、Cz、Pz点的总平均波形(标准序列ⅰ为组段6、18的平均值,标准序列ⅱ为组段10、14的平均值)Fig.3 Group average waveform on electrode of Fz,Cz,Pzin three different sequences ofexplicit and implicit groups
从ERP波形(见图3)也可以看出内隐和外显被试对于标准序列和迁移序列诱发的波形有显著差异。不管是内隐被试还是外显被试,其ERP总时程内都出现了显著的N1成分(80~160 ms,头皮前中部明显)、P2成分(160~220 ms)、N2成分(220~300 ms)、P3成分(250~420 ms,头皮中后部明显)。两组被试的脑电数据按标准序列、刺激迁移序列和效应器迁移序列分别叠加。根据既往研究,对于序列学习知识表征的研究主要应用N2、P3作为指标,所以我们只研究这两个成分的变化。根据总平均图与参考文献确定ERP成分及其时间窗口。N2的时间窗口为刺激呈现后的220~300 ms,P3为250~420 ms。
使用重复测量方差分析来检验不同迁移类型对于学习结果的影响,以及这种影响在外显组和内隐组是否存在不同。方差分析的P值皆用Greenhouse-Geisser法校正。对N2成分进行方差分析的4个因素为序列类型(3水平:标准序列,刺激迁移序列,效应器迁移序列)、学习类型(2水平:内隐组,外显组),左右电极位置(3水平:左(F3,C3,P3,Po7)、中线(Fz,Cz,Pz,Oz),右(F4,C4,P4,Po8))、前后电极位置(4水平:前(F3,Fz,F4)、前中(C3,Cz,C4),后中(P3,Pz,P4),后(Po7,Oz,Po8))。对于P3成分的分析也采用同样的统计方法,不过选取的电极有所不同,左右电极位置因素的3个水平为左(C3,P3,Po7)、中线(Cz,Pz,Oz)、右(C4,P4,Po8),前后因素的3个水平为前(C3,Cz,C4),中(P3,Pz,P4)、后(Po7,Oz,Po8))。
研究并没有将标准序列的所有组段进行平均,而是将刺激迁移序列和标准序列ⅰ(组段6、18的平均值)以及效应器迁移序列和标准序列ⅱ(组段10、14的平均值)以两组重复测量方差分析分别加以比较,选取的标准序列组段均为迁移序列的前一组(例如,刺激迁移序列为组段7、19,那么与之相比较的标准序列ⅰ则为组段6、18;另一组亦同)。这样是为了更好地考察特定的干扰对于已习得规律的影响,同时也为了防止由于平均带来的对试验数据的污染。
对于N2的波幅值,将刺激迁移序列和标准序列ⅰ(组段6、18)以及效应器迁移序列和标准序列ⅱ(组段10、14)两组分别加以分析。在对刺激迁移序列和标准序列ⅰ(组段6、18)的分析中,发现学习类型主效应不显著,学习类型与序列类型、与前后电极位置、与左右电极位置间的交互作用均不显著。序列类型主效应不显著,且与前后电极位置不存在交互作用,但与左右电极位置存在显著的交互作用(F(2,38)=3.944,P<0.05)。进一步简单效应分析发现,在头皮的左、中线、右3个水平上刺激迁移序列与标准序列均不存在差别(均为P>0.05)。
对于效应器迁移序列和标准序列ⅱ(组段10、14)分析发现学习类型主效应显著(F(1,19)=5.056,P<0.05),然而学习类型与序列类型、与前后电极位置、与左右电极位置间的交互作用均不显著。序列类型主效应不显著,与左右电极位置不存在显著的交互作用,但与前后电极位置交互作用显著(F(3,57)=4.778,P<0.05)。进一步分析发现,在前后电极位置因素的前中位置,效应器迁移序列和标准序列存在显著差异((-0.885±0.509)μV,-2.185±0.637 μV,P<0.05)。
在对于刺激迁移序列和效应器迁移序列的方差分析中,只有前后电极位置,左右电极位置与序列类型的三因素交互作用显著(F(6,114)=2.778,P<0.05),但进一步的简单效应分析发现在每一个位置上,两种序列都不存在差异(P>0.05)。
对于P3的波幅值,将刺激迁移序列和标准序列(组段6、18)以及效应器迁移序列和标准序列ⅱ(组段10、14)也分别加以分析。对于刺激迁移序列和标准序列ⅰ的分析中,学习类型主效应不显著,且学习类型与序列类型、与前后电极位置间的交互作用均不显著,与左右电极位置(F(2,38)=1.382,P<0.05)间的交互作用显著,进一步的简单效应分析并未发现差别(P>0.05)。
对于效应器迁移序列和标准序列ⅱ(组段10、14)分析发现,发现学习类型因素不显著,序列类型与电极位置因素不存在交互作用;学习类型与前后电极位置和左右电极位置同样不存在交互作用,但学习类型和序列类型之间交互作用显著(F(1,19)= 5.53,P<0.05)。进一步的简单效应分析发现,外显组t(9)=4.298,P<0.05,内隐组t(10)=0.589,P>0.05。
对于刺激迁移序列和效应器迁移序列的方差分析发现学习类型与电极位置因素交互作用皆不显著,但与序列类型却存在显著的交互作用F(1,19)=8.29,P<0.05),进一步分析发现只有外显组的两种序列存在差别(t(10=-2.83,P<0.05)。序列类型与前后电极位置的交互作用不显著,而与左右电极位置则交互作用显著(F(2,38)=3.675,P<0.05),简单效应分析发现两种序列类型在3个水平上均未出现差异(P>0.05)。
4讨 论
4.1 效应器水平编码的存在
随着序列学习研究的逐步深入,越来越多的研究者开始关注序列知识的表征问题[28-29]。那么,知觉动作序列知识的学习是否可以独立于执行这些动作的效应器系统呢?有人认为既然动作序列的知识可以在不同效应器之间自由的迁移,那么对于该序列的编码过程应该不涉及效应器水平[18,30]。这种观点显而易见的问题是既然存在迁移代价(如转移至不同的效应器以后导致反应时延长),就不能认为序列学习与效应器无关。在本试验中,被试通过对前6个组段标准序列的学习,已经学会了一定的序列知识,随后在效应器迁移序列,内隐组和外显组的被试的成绩均表现出一定程度的下降,这说明,序列知识的表征并不能完全独立于效应器水平。同时,在脑电数据中的N2和P3成分的波幅上,效应器迁移序列较之标准序列和刺激迁移序列都出现了显著的差异,这也说明了效应器迁移对于正常的序列学习进程出现了干扰。
4.2 不同学习“方式”对于序列学习知识表征的影响
本研究使用的试验材料为知觉动作序列,那么在学习进程中,被学习的序列则主要为知觉序列和动作序列。那么,对知觉序列的干扰形成了刺激迁移序列,而对运动序列的干扰形成了效应器迁移序列。从得到的行为数据看来,内隐组被试在由标准序列向刺激迁移序列和效应器迁移序列迁移的时候,都表现为行为效率下降,但内隐组与外显组的具体情况却有所不同。内隐组在效应器迁移序列中的反应时长于刺激迁移序列,而在外显组中被试在这两种迁移序列中的反应时并不存在明显差异。这表明,虽然两组学习者都受到了效应器迁移的影响,但较之外显组,内隐组学习者受到的影响更多。这意味着在内隐学习过程中序列知识的编码过程更多地涉及动觉加工。
在本试验得到的ERP数据中,被试习得一定的序列知识以后,外显组被试迁移至效应器迁移序列时出现N2和P3波幅的改变,但在迁移至刺激迁移序列时没有相应的变化;内隐组在迁移至效应器迁移序列的时候出现了N2波幅的改变,但P3波幅没有显著变化。在这里内隐序列学习和外显序列学习出现了分离,证实内隐序列学习和外显序列学习的学习机制存在不同,另外,动觉加工对于外显和内隐被试序列知识的表征可能都是必须的。
具体来说,通过行为数据的结果,我们可以看出:在内隐组,在从标准序列向改变刺激序列的刺激迁移组段和改变效应器的效应器迁移组段迁移的过程中都出现了显著的反应时的变慢(正确率保持不变的情况下),这可能说明了在知识迁移的过程中不管是外界刺激还是效应器的变化都会对其造成影响。不过,在对两种迁移结果的比较当中,发现不同于内隐组的差异显著,外显组的两种迁移结果并不存在差别,这可能说明两种学习类型在序列学习的过程中对于不同序列知识的编码方式可能存在不同,同时也说明了迁移并不是一种只局限于外显知识的现象。然而,在脑电数据中,在N2的平均波幅作为指标的情况下,发现在因素D的a2水平即中央区脑区,标准序列与刺激迁移序列不存在区别,而与效应器迁移序列却差异显著,这种情况在外显组和内隐组并不存在差别。同时,对于P3的平均波幅的分析得出这样的结果:序列类型因学习类型的不同而出现了差别。在内隐组,标准序列与效应器迁移序列不存在区别,而在外显组,两者却差异显著。这似乎说明了在特定的学习类型下,效应器编码对于序列学习的表征起了一定的作用。相对于知觉编码,效应器编码起的作用更大。
4.3 动觉加工所涉及的内部机制
以往关于序列反应时任务中知识表征方式的观点主要有3种,分别认为是S-R(刺激-反应)表征、S-S(刺激-刺激)表征和R-R(反应-反应)表征[20,31-34]。我们认为,将序列学习知识的表征仅仅归结为刺激和反应两种水平可能过于简单化。在序列学习试验中,被试完成的任务类似于选择反应时任务,起码要经历刺激辨别—反应抉择—动作发出3个阶段。这3个阶段都有可以因为学习而改善,从而导致学习效应的产生。此外,在此类任务中一个经常被忽视的可能的学习机制是动觉反馈。按键的手指在做出反应动作的同时,也会向中枢发出感觉信号,这种信号可以和在屏幕上呈现的字母序列一样,也可以成为学习的对象。因此,序列学习的内部过程或许可以用图4表示出来。

图4 序列学习的内部过程Fig.4 Inner process of sequence learning
如果序列学习过程正如图4所示的那样,序列学习过程就不像过去人们认为的那样,仅仅涉及到学习者序列刺激物辨别能力的改善和动作能力的改善。本试验倾向于用上面的模型解释得到的结果。在本试验中,无论是内隐学习组还是外显学习组,当被试向效应器迁移序列迁移的时候,N2成分都出现波幅的变化。目前,N2被认为是一个与朝向反应、刺激辨别、目标选择、任务要求、反应抑制等多种过程相关的ERP成分[33]。近来有些研究者认为它与认知控制和刺激的失匹配有关[34]。N2成分上出现的效应器迁移序列相对于刺激迁移序列的更大的干扰效应有可能除了动觉本身的编码机制之外还反映了动觉的反馈作用。
4.4 试验中需要进一步讨论的问题
根据本试验的设计中采用整体进行迁移进行干扰的特点,有如下两个问题似乎需要进一步讨论。第一,虽然证明效应器迁移序列和标准序列在表征方式上存在差异,但是从N2的波幅上来看,结果并不是迁移后的波幅值比标准序列更负[19-20],而是出现了相反地结果。我们分析原因可能来自于成组进行迁移可以判断干扰的方式主要是建立在对于长时存储表征进行干扰的基础上的,而完全不同于既往研究中随机的单个插入偏差的情况,该种情况则完全是对于短时记忆表征的破坏。根据Eimer[18]同类研究的结果所提示的,N2b的波幅可能反映了被试所习得的关于刺激序列的有意识的知识的量。那么效应器迁移组N2b波幅的下降,则极有可能是因为在迁移过程中关于序列知识的外显部分减少造成的。第二,随机的单个插入偏差的研究方法由于需要对许多单个偏差刺激进行叠加平均,这增加了该类刺激的内部不一致性,不恰当的平均可能带来对试验数据的污染。
另外,在对于P3的分析中,根据既往文献,P300反映了被试对刺激材料的一部分主观估计,Wakraud和Wolfgar[37]发现在P300上的任务评估机制对于目标刺激概率更为敏感(相对于其他的同样反映预期估计的脑电指标,如CNV)。在本研究中,按照预想N2波幅在标准序列向迁移序列改变的过程出现的变化趋势也应该同样出现在P3的波幅变化上,但是,试验数据波形并未出现非常明显的正向成分。导致这种情况出现的原因可能是,标准刺激和偏差刺激都是作为靶刺激出现的,而P3效应一般出现在一系列非靶刺激中的偏差靶刺激的情况下。Courchesne,Courchesne,和Hillyard的研究中偏差刺激既可以作为靶刺激又可以作为非靶刺激[38]。这种情况下非靶刺激偏差引起的波幅明显小于靶刺激的。在我们的试验中,可能也是因为所有的刺激都是任务相关的(靶刺激),所以削弱了P3的波幅。
5 结论
本研究表明,序列学习的知识表征方式首先并不完全是独立于效应器的,在某些水平,效应器水平的编码机制是参与对于序列知识的学习的,并且相对于外界刺激,效应器编码起的作用更大。另外,动觉加工的表征方式对于外显和内隐学习者序列知识的表征可能都是必须的。
[1]Raibert M H.Motor control and learning by the state space model[M]. Cambridge,MA:Unpublished DoctoralDissertation,MassachusettsInstitute ofTechnology,1997.
[2]Margolin D I.The neuropsychology of writing and spelling:Semantic,phonological,motor,and perceptual processes[J].Quarterly Journal of Experimental Psychology,1984,36A:459-489.
[3]Hillis A E,Caramazza A.The graphemic buffer and attentional mechanisms[A].Baltimore,MD:Johns Hopkins University,1998.
[4]Cohen A,Ivry R I,Keele S.Attention and structure in sequence learning [J].Journal of Experimental Psychology:Learning,Memory,and Cognition,1990,16(1):17-30.
[5]Remillard G.Pure perceptual-based sequence learning[J].Journal ofExperimental Psychology:Learning,Memory,and Cognition,2003,29:581-597.
[6]Keele S,Jennings P,Jones S,et al.On the modularity of sequence representation[J].Journal of Motor Behavior,1995,27:17-30.
[7]Stadler M A.On learning complex procedural knowledge[J].Journal of Experimental Psychology:Learning,Memory,and Cognition,1989,21:674-685.
[8]Keele S,Davidson M,Hayes A.Sequential representation and the neural basis of motor skill[M].Eugene:Institute of Cognitive and Decision Sciences,UniversityofOregon,1996.
[9]Bapi R S,Doya K,Harner A M.Evidence for effector independent and dependent representations and their differential time course of acquisition during motor sequence[J].Experimental Brain Research,2000,132:149-162.
[10]Nissen MJ,Bullemer P.Attentional requirements of learning:evidence fromperformance measures[J].Cognitive Psychology,1987,19:1-32.
[11]Jiménez L,Méndez C.Implicit sequence learning with competing explicit cues[J].Quarterly Journal of Experimental Psychology,2001,54A:345-369
[12]Perruchet P,Amorim M A.Conscious knowledge and changes in performance in sequence learning:Evidence against dissociation[J]. 1992,18:785-800.
[13]Shanks D R,Johnstone T,Staggs L.Abstraction process in artificial grammar learning[J].Quarterly Journal of Experimental Psychology,1997,50A:216-252.
[14]张卫.内隐知识表征的性质与研究 [J].心理学探新,1999,72(4):26-30
[15]付秋芳,傅小兰.内隐学习中表征与意识的关系[J].心理科学进展,2006,14(1):18-22.
[16]郭秀艳.内隐学习和外显学习的关系评述 [J].心理科学进展,2004,12:185-192
[17]Fu QF,Liu YF,Fu XL.The effects oftype and feature ofknowledge on implicit sequence learning[J].Journal of Chinese Psychology Acta Psychologica Sinica,2004,36(5):525-533.
[18]Eimer M,Goschke T,Schlaghecken F,et al.Explicit and implicit learning ofeventsequences:Evidence from event-related brain potentials[J].Journal of Experimental Psychology:Learning,Memory,and Cognition,1996,22(4):970-987.
[19]Russeler J,Rosler F.Implicit and explicit learning of event-sequences:evidence for distinct coding of perceptual and motor representations[J].Acta Psycho(lAmst),2000,104:45-67.
[20]Jascha Russeler,Erwin Hennighausenb,Thomas F,et al.Differences in incidental and intentional learning of sensorimotor sequences as revealed byevent-related brain potentials[J].Cognitive Brain Research,2003,15:116-126.
[21]Jiménez L,Vázquez1 G A.Sequence learning under dual-task conditions:Alternatives to a resource-based account[J].Psychological Research,2005,69(5/6):352-368.
[22]Altmann G T M,Diebes Z,Goode A.On the modality independence of implicitly learned grammatical knowledge[J].Journal of Experimental Psychology:Learning,Memory,and Cognition,1995,21:899-912.
[23]Knowlton B J,Squire L R.Artificial grammar learning depends on implicit acquisition of both abstract and exemplar-specific information [J].Journal of Experimental Psychology:Learning,Memory,and Cognition,1996,22:169-181.
[24]Mathews R C,Buss R R,Stanley W B,et al.Role of implicit and explicit processes in learning from examples:A synergistic effect[J].Journal of Experimental Psychology:Learning,Memory,and Cognition,1989,15(6):1 083-1 100.
[25]Musen G,Squire L R.Implicit learning of color-word associations using a stroop paradigm[J].Journal of Experimental Psychology:Learning,Memory,and Cognition,1993,19(4):789-798.
[26]Reber A.Transfer of syntactic structure in synthetic languages[J].Journal of Experimental Psychology,1969,81(1):115-119.
[27]Willingham D B,Wells L A,Farrell J M.Implicit motor sequence learning is represented in response locations[J].Memory&Cognition,2000,28(3):366-375.
[28]王树明,章建成.知觉运动技能训练的国外研究进展[J].上海体育学院学报,2005,29(3):60-64.
[29]陈耕春.高水平运动技能训练中的运动知觉心理训练取向 [J].西安体育学院学报,2007,24(3):9-12.
[30]ZieBler M.The impact of motor responses on serial-pattern learning[J].Psychological Research,1994,57:30-41.
[31]Willingham D B,Nissen M J,Bullemer P.On the development of procedural knowledge [J].JournalofExperimentalPsychology:Learning,Memory,and Cognition,1989,15:1 047-1 060.
[32]Howard Jr J H,Mutter S A,Howard D V.Serial pattern learning by event observation[J].Journal of Experimental Psychology:Learning,Memory,and Cognition,1992,18(5):1 029-1 039.
[33]Mayr U.Spatial attention and implicit sequence learning:evidence for independent learning of spatial and nonspatial sequences[J].Journal of Experimental Psychology:Learning,Memory,and Cognition,1996,22(2):350-364.
[34]Nattkemper D,Prinz W.Stimulus and response anticipation in a serial reaction task.Psychological Research:Sequence learning(special issue)[J].Phenomena and Models,1997(2):98-112.
[35]Alexandra P,Fonaryova Key,Guy O,et al.Linking Brainwaves to the Brain:An ERP Primer[J].Developmental Neuropsychology,2005,27(2):183-215.
[36]Jonathan R Folstein,Cyma VAN Petten.Influence of cognitive control and mismatch on the N2 component of the ERP:A review[J].Psychophysiology,2007,44:1-19.
[37]Wahraud,Wolf gar,Viviane,et al.Differential effects of the stimulus sequence on CNVand P300[J].Brain Research,2006,1 123:157-167.
[38]Courchesne E,Hillyard S A,Courchesne R Y.P3 waves to the discrimination of targets in homogeneous and heterogeneous stimulus sequences[J].Psychophysiology,1997,14:590-597.
Influence of Different Transfer and Different Learning“Strategy”on the Epresentations of Sequence Knowledge:An ERP Study
HU Wei,LÜ Yong
(Academy of Psychology and Behavior,Tianjin Normal University,Tianjin 300074,China)
The present study investigated implicit and explicit learning representations'difference in a modified serial reaction time task,using N2b,P3b as index.38 subjects were grouped randomly to intentional or incidental group,respectively.Subjects had to learn a 12-letter-long repeating sequence that was irregularly disrupted by deviating sequence,including perceptual deviant and motor deviant.After the task,all participants were asked to complete the generation task.Then explicit and implicit learners were separated with the scores.The result suggested that representations of sequence knowledge in sequence learning were not totally independent of effectors.To some extent,effectors coding were involved in learning of sequence knowledge;Different type sequence knowledge may connect with specific independent mechanism,but not co-represent.The way to represent sequence knowledge between implicit and explicit learner was different,the motor representations(effectors-related)played a more important role than the perceptual way(stimulus-related)in both way to sequence learning.The study revealed that different sequence involved different representations in the different stage,and the process of forming representations is related to different mechanism of our learning system,also proved that implicit and explicit learning representations are different.
perceptual-motor sequence;transfer;knowledge representations style;ERPs(Event-Related Potentials)
G 804.8
A
1005-0000(2011)04-0277-07
2011-03-25;
2011-05-09;录用日期:2011-05-15
教育部人文社会科学研究基地重大项目(项目编号:05JJDXLX003);全国高校优秀博士论文作者专项资金项目(项目编号:200708);天津师范大学哲学社会科学研究基金项目(项目编号:52WM73)
胡 伟(1980-),男,天津市人,助理实验师,研究方向为认知神经科学。通讯作者:吕 勇(1963-),男,天津市人,博士,教授,研究方向为认知神经科学。
天津师范大学心理与行为研究院,天津300074。
