发酵秸秆糖产丁二酸放线杆菌的CO2固定关键酶特性分析
2010-10-25薛培俭姜绍通李兴江杨为华潘丽军
薛培俭, 姜绍通, 李兴江, 杨为华, 潘丽军
(1.安徽丰原集团控股有限公司,安徽 蚌埠 233010;2.合肥工业大学 生物与食品工程学院,安徽 合肥 230009)
丁二酸广泛应用于化工、食品及医药等领域,丁二酸可以合成百种以上的其它化学品,是最为重要的平台化学品之一。目前丁二酸主要依赖石化原料制备,使其广泛应用受到限制[1]。
农作物秸秆是最重要的生物质,以可再生的秸秆糖为原料发酵生产丁二酸日益受到人们的重视,尤其因为其发酵菌种具有固定CO2的特征,不仅摆脱了对石化原料的依赖,更开辟了温室气体CO2利用的新途径,使得丁二酸的微生物转化成为一个最具发展潜力的绿色模式[2]。
丁二酸作为三羧酸循环的中间体,在其发酵过程中,由其前体物质草酰乙酸的两步氢化反应制备得到[3],而草酰乙酸最主要的生成途径是来自于磷酸烯醇式丙酮酸与CO2的固定,这一途径既是该类菌吸收温室气体(CO2)的主要途径,也是其代谢关键酶所在。
文献[4-6]研究表明,作为兼性厌氧产丁二酸菌,丁二酸放线杆菌(Actinobacillus succiniogenes)胞内具有一大类重要的酶,其中磷酸烯醇式丙酮酸羧化激酶充当着最为重要的角色,在发酵过程中该酶保持着相当高的酶活。因此,本文在对菌株该关键酶克隆测序基础上,围绕该关键酶的酶活测定以及各种因素对关键酶酶活力的影响进行了重点分析,以期为发酵工艺的调控提供指导。
1 材料与方法
1.1 材 料
菌株为产丁二酸放线杆菌(Actinobacillus succinogenes S.JST),为本实验室保藏。
发酵培养基为:酵母浸粉 10 g/L,玉米浆(CSL)15 g/L,Na2HPO4◦12H2O 0.7 g/L,NaH2PO4◦2H2O 1.16 g/L,NaCl 1.0 g/L,CaCl20.2 g/L,MnCl2◦6H2O 0.2 g/L,MgCO340 g/L,秸秆糖80 g/L,胰蛋白胨大豆肉汤(TSB)5 g/L。培养条件为在37℃摇瓶CO2厌氧发酵。
1.2 试剂和仪器
PCR反应及DNA提取、回收试剂均购自大连宝生物工程公司;细胞破碎仪为JY92-Ⅱ系统;高效毛细管电泳色谱(HPCE)采用美国Beckman Coulter公司P/ACE MDQ型毛细管电泳仪;厌氧培养系统为CBJS-5B/1。
1.3 Pepck关键酶基因克隆测序
克 隆 引 物 相 同,H1:5′-CCAACTAAT TTAGCCGCT TCC-3′,H2:5′-T TAGGGCT TACCGATGTTAAG-3′。
PCR反应:25 μ L PCR反应体系中,dNTP(2.5 m)为 2 μ L,10 ×缓冲线为 2.5 μ L,H1(20 pmol)为1 μ L,H2(20 pmol)为 1 μ L,Taq 为0.2 μ L,模板DNA 为 1 μ L,用水补足到 25 μ L 。
PCR反应程序为:94℃5 min;94℃40 s,
55℃40 s,72℃100 s,30个循环;72℃10 min。
PCR产物回收、连接、转化及测序:试剂盒纯化 DNA;取 1 μ L回收的 PCR产物,按照pMD18T载体试剂盒操作,16℃连接1 h以上;取连接混合物3.5 μ L加到200 μ L感受态细胞液中,冰浴30 min后热击转化;对阳性克隆株进行PCR鉴定;由大连宝生物工程公司完成序列测定。
1.4 酶活检测
1.4.1 细胞酶液制备
取10 mL发酵培养菌悬液10 000 r/min离心获得菌体,pH值为7.5,0.1 mol/L磷酸盐缓冲液稀释至10 mL,冰浴超声粉碎10 min(超声2 s/间隔5 s),4℃下 10 000 r/min冷冻离心15 min,获取上清液酶液。其中菌液为37℃条件下CO2厌氧气室摇瓶培养24 h所得。之后取上清初酶液进行硫酸铵沉淀纯化,采用35%~55%饱和硫酸铵沉淀制备,即先用35%饱和硫酸铵沉淀,8 000 r/min冷冻离心15 min后,除去杂蛋白,取离心所得的上清液;而后饱和硫酸铵质量分数增至55%,10 000 r/min冷冻离心15 min后,获取沉淀,将沉淀物用0.1 mol/L磷酸盐缓冲液(pH=7.5)稀释至10 mL,再用透析袋在4℃条件下3 h内反复透析6次(透析液为pH=7.5的0.1 mol/L磷酸盐缓冲),透析结束后定容至10 mL,此时得到初步纯化酶液。
1.4.2 磷酸烯醇式丙酮酸羧化激酶测定
加入3 mL pH值为7.5,0.1 mol/L磷酸盐缓冲液进对照比色皿中,测定比色皿中加2.3 mL缓冲液,依次加入5 mmol/L MnCl2、0.5 mmol/L碳酸氢钠、0.5 mmol/L磷酸烯醇式丙酮酸钠、50 mmol/L的GDP(鸟嘌呤核苷二磷酸)、5 mmol/L NADH(还原辅酶Ⅰ)溶液各0.1 mL,置于25℃水浴中保温,分别快速加入0.1 mL的500 U/mL苹果酸脱氢酶纯酶液与适量待测酶液,启动反应;在340 nm处,3 min内每30 s以对照比色皿调零,记录吸光度值,计算一定时间内吸光度的降低值ΔA。
1.4.3酶活的计算
酶活单位计算在本方法相当于每分钟直接或间接消耗1 μ mol NADH量,计算式如下:

酶活为每毫克酶蛋白所具有的酶活力,单位为U/mg,计算式如下:

其中,V为酶促反应体积,指比色皿中酶液体积,为3 mL;ΔA为340 nm处吸光度的变化值,无量纲;b为比色皿的光程,取1 cm;ε为NADH的表观摩尔消光系数,取6.22×103 L/(mol◦cm);Δ t为时间间隔,取3 min;x为向反应比色皿内加入的待测酶液体积;n为稀释倍数,n=3 000/x;ρ为比色皿中酶蛋白的质量浓度。
2 结果与分析
2.1 菌株的Pepck关键酶基因克隆分析
对菌株Pepck基因分析表明(Accession No.EU253567),该基因有1 617个碱基序列,编码的蛋白酶为裂解酶类中的羧激酶,国际酶学编号为EC4.1.1.32。
基于所获序列及酶催化方式初步构建其蛋白结构如图1所示。

图1 磷酸烯醇式丙酮酸羧化激酶的蛋白质结构
通过生物信息学比较,发现该酶的结构蛋白中除了具有经典的底物结合位点及辅酶结合位点外,分别发现激酶常见的 Mn2+与Mg2+结合位点,在264的异亮氨酸与272位的天冬氨酸分别为Mn2+结合位点,在211位的赖氨酸与235位的缬氨酸分别为Mg2+结合位点。
2.2 酶活分析
在整个发酵36 h期间,每6 h取一次样,对样品进行酶活检测,结果如图2所示。
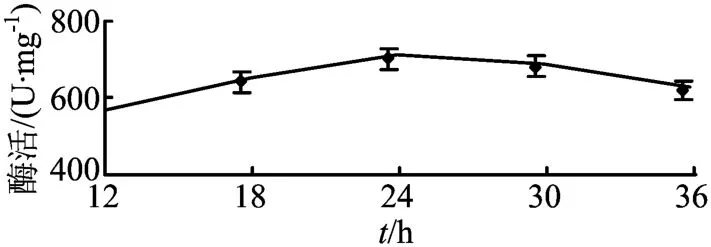
图2 不同发酵阶段磷酸烯醇式丙酮酸羧化激酶的酶活
通过酶活检测分析,发现发酵过程中该酶均维持较高酶活,尤其发酵中期24 h左右,酶活最高,因此后续条件试验均选取此时的酶进行分析。
2.3 pH值对酶活影响
发酵体系中的pH值过低或过高对酶的催化都有不利影响,甚至使酶失活。本文测定了酶液在不同pH值缓冲液中的酶活,结果如图3所示。

图3 不同pH值对磷酸烯醇式丙酮酸羧化激酶的影响
从图3可以看出,菌株的Pepck活性对环境的pH值变化较敏感,pH值为6.5~7.0时酶活较高,因此在发酵过程中应及时添加中和剂,否则生成丁二酸过量则会造成发酵液的pH值过低,会进一步抑制Pepck活性,而关键酶的被抑制对总体发酵是不利的。
2.4 不同离子对酶活的影响
在反应体系中分别加入不同浓度的MgSO4、ZnSO4、MnSO4及 CuSO4,考察离子对 Pepck酶活的影响,其结果如图4所示。

图4 不同离子对磷酸烯醇式丙酮酸羧化激酶的影响
由图4可以看出,Mn2+与Mg2+对磷酸烯醇式丙酮酸羧化激酶具有明显的激活作用,Mn2+的浓度为0.1~0.2 mmol/L时就几乎将酶活潜力完全激发了,而Mg2+的浓度要达到0.5 mmol/L以后方可检测到酶的高活力。Zn2+对该酶没有明显影响,而Cu2+则对酶活具有明显的抑制作用。因此在发酵过程中应尽量减少Cu2+的存在,并提供一定的Mg2+浓度。同时在对2.1节中获得的酶基因进行保守序列分析时也发现,该类酶具有不止一处的Mg2+与Mn2+激活位点。
2.5 底物的影响
该酶在体系中催化的反应为:
磷酸烯醇式丙酮酸+GDP+CO2(溶液体系中以HCO3-)=草酰乙酸+GT P。
分别以不同浓度的底物磷酸烯醇式丙酮酸、鸟嘌呤核苷二磷酸(GDP)及HCO3-进行反应,测定相应酶活,根据酶促反应动力学的 Hanes-Woolf法,求得该酶对以上底物的 Km依次为3.0×10-4mol/L、8.0×10-5mol/L及 7.0×10-2mol/L,由此可见,该磷酸烯醇式丙酮羧化激酶对HCO3-的亲和力最弱(H2CO3/HCO3-的电离常数为4.3×10-7,在缓冲体系中性的情况下,HCO3-转化为CO2不是限制因素,所以该酶对HCO3-的亲和力可以近似等效于对CO2的亲和力),因此若要保证发酵过程中产物的高效生成必须持续提供高浓度的HCO3-。
3 结束语
Actinobacillussuccinogenes是一种从牛瘤胃中分离产丁二酸菌[7],由于牛瘤胃具有高CO2含量的厌氧特性,使得所筛菌株具有固定CO2的能力,同时这也是该类菌株最显著的特征,因此围绕其关键酶(磷酸烯醇式丙酮酸羧化激酶)开展研究具有重要意义。
本文在对关键酶克隆及酶活测定的基础上,重点分析了pH值、离子及底物对酶特性影响。其中pH值影响分析表明,必须持续添加中和剂,保证发酵过程中的产物丁二酸持续地累积,否则过酸的环境将对关键酶产生明显的酶活抑制。不同离子的影响表明,有的离子具有明显的抑制作用,如Cu2+,有的离子具有明显的激活作用,如Mn2+与Mg2+,这就要求发酵过程中控制恰当的离子浓度,否则关键酶的抑制可能影响总体发酵。
酶促反应动力学分析表明,该酶对HCO3-亲和力较弱,该酶的这种特性要求发酵过程中必须有足够的HCO3-。因此在发酵过程中,维持发酵体系中添加过量的MgCO3作为中和剂,既可以维持发酵体系的 pH值中性,又可以提供Mg2+,同时又能保证溶液中一定量的HCO3-,这只是针对磷酸烯醇式羧化激酶独立而言,就整个发酵体系而言,菌株可能还有其它更严格的要求,如极端的CO2厌氧条件等[8]。
[1] Chotani G,Dodge T,Hsu A,et al.The commercial production of chemicals using pathway engineering[J].Biochimica et Biophy sica Acta,2000,1543(2):434-455.
[2] Lee S Y,Hong S H,Lee S H,et al.Fermentative production of chemicals that can be used for polymer synthesis[J].M acromol Biosci,2004,4(3):157-164.
[3] 王庆昭,赵学明.产丁二酸菌种研究进展[J].生物工程学报,2007,23(4):570-576.
[4] M cKinlay J B,Shachar H Y,Zeikus J G,et al.Determining Actinobacillus succinogenes metabolic pathways and fluxes by NM R and GC-MS analyses of C-13-labeled metabolic product isotopomers [J].M etabolic Engineering,2007,9(2):177-192.
[5] McKinlay J B,Vieille C.13C-metabolic flux analysis of Actinobacillus succinogenes fermentative metabolism at different NaHCO3and H2concentrations[J].M etabolic Engineering,2008,10(1):55-68.
[6] Cotelesage J J H,Prasad L,Zeikus J G,et al.Crystal structure of Anaerobiospirillum succiniciproducens PEP carboxy kinase reveals an impo rtant active site loop[J].InternationalJournal of Biochemistry and Cell Biology,2005,37(9):1829-1837.
[7] 刘宇鹏,郑 璞,倪 晔,等.抗氟乙酸突变株的选育及其代谢流量分析[J].生物工程学报,2008,24(3):460-467.
[8] 谢 鑫,陈可泉,刘忠敏,等.产丁二酸工程菌的构建及其厌氧发酵[J].生物工程学报,2008,24(1):101-105.
