一个灵长类种属特异基因—XAGE-3转基因
2010-09-09向志光陈炜全雄志董伟曹兴水高珊张晓娟张海涛张连峰
向志光,陈炜,全雄志,董伟,曹兴水,高珊,张晓娟,张海涛,张连峰
(中国医学科学院北京协和医学院实验动物研究所,卫生部人类疾病比较医学重点实验室,北京 100021)
研究报告
一个灵长类种属特异基因—XAGE-3转基因
向志光,陈炜,全雄志,董伟,曹兴水,高珊,张晓娟,张海涛,张连峰
(中国医学科学院北京协和医学院实验动物研究所,卫生部人类疾病比较医学重点实验室,北京 100021)
目的分析XAGE-3基因在灵长类和啮齿类动物的基因组中的同源性,通过转基因研究XAGE-3基因在小鼠中的功能及生物学意义。方法根据Homologene及Taxplot数据库,通过Blast比对方法分析XAGE-3在两类动物基因组中的同源性;从人胎盘组织克隆XAGE-3基因,转入真核表达载体pCDNA3.1(+),显微注射方法得到转基因动物,基因组PCR鉴定基因型,反转录PCR分析基因的表达;Brdu标记3周龄动物显示睾丸内细胞的增殖。结果XAGE-3在人、黑猩猩和猕猴中存在高度同源基因,而在小鼠和大鼠中无同源区域;在基因型鉴定阳性的5个首建系中3个品系睾丸组织目的基因表达较高,在传代的两个品系中,在小肠,胸腺,睾丸等组织中目的基因均有表达;睾丸组织Brdu标记显示XAGE-3转基因动物有更多的发育晚期的精细胞被标记。结论XAGE-3作为灵长类种属特有基因在转基因小鼠中影响了精细胞发育。
XAGE-3;种属特异基因;转基因
生物物种的进化本质是遗传物质基因组的进化,包括基因的融合、倒位、易位、基因的丢失、新基因的出现等。这些正是促使物种发生分化的基础。基因组学研究使得我们可以在DNA序列水平比较人类和其他生物的差异,从而找到生命进化的内因。小鼠作为成熟的模式生物是除人类之外基因组研究较为清楚的生物。通过比较,我们发现小鼠和人类都有自己相对独立的一些基因,这些基因在另外的物种基因组中是找不到的。
我们根据Homologene数据库[1]对于小鼠(Musmusculus)、人(Homo sapiens)、大鼠(Rattus norvegicus)、黑猩猩(Pan troglodytes)等的同源基因进行了比较,找出一些差异基因,又对找出的差异基因用Blast程序针对其他基因组进行了比对,还在蛋白水平检索了Taxplot[2]数据库,从而找到了人类与小鼠存在差异的基因,这些基因存在一些共性,在基因的功能上更多的是和生殖、免疫、嗅觉等相关。
XAGE-3基因(X antigen family,member 3)就是通过上面的方法找到的一种灵长类种属特异基因,该基因在人类的近亲黑猩猩和猕猴(Macaca mulatta)存在同源基因,而在小鼠和大鼠中不存在该基因的同源基因。根据EST数据库,XAGE-3基因位于人类的X染色体短臂11区21至22带之间,它主要在人类胎盘和早期胚胎组织中表达。由于存在着变位剪切,该基因有两个转录本(NM_133179,NM_130776),但是两个转录本的蛋白编码序列是相同的。因为和XAGE1D基因(X antigen family,member 1D)位置靠近,被命名为XAGE-3,而实际上该基因在序列上和XAGE1D无相似性。为了研究该基因的可能功能,我们构建了以CMV为启动子的转基因载体,通过显微注射得到XAGE-3转基因小鼠。
1 材料和方法
1.1 材料
实验中使用的动物为C57BL/6J小鼠购自北京康蓝生物技术有限公司【SCXK(京)2005-0013】。实验中涉及动物的操作程序已经得到中国医学科学院医学实验动物研究所实验动物使用与管理委员会的批准(GC-08-2032)。
1.2 数据库的使用
实验使用了NCBI数据库Homologene,限定物种,采用逻辑减法,筛选灵长类种属特异基因,即Homo sapiens-Mus musculus,同时参考两个物种的近缘物种大鼠和黑猩猩的比较信息。也使用了关键词“gene exclusive to homo sapiens”检索Homologene数据库。XAGE3即在这一策略下检索出,又对小鼠基因组在DNA和蛋白水平进行比对,参考TaxPlot数据库,确定基因的种属特异性。
1.3 基因的克隆
使用Trizol(Invitrogen)提取人胎盘组织总RNA,用Superscript III反转录试剂盒(Invitrogen)以OligodT合成cDNA第一链,以此为模板扩增目的基因。上下游引物分别为5′-ttggtaccgcagctgtgtgaaatatg- 3′;5′-gcgaattccttcatttaaacctgtgg-3′。经94℃3 min预变性,后扩增30循环,依次为变性94℃30 s,复性58℃30 s,延伸72℃1 m in。最后72℃延伸10 m in。
1.4 载体的构建和显微注射
扩增的目的基因两端带有酶切位点Kpn I和EcoR I,插入真核表达载体pCDNA3.1(+),转化感受态细胞DH5a,挑选克隆,提取质粒,酶切鉴定,测序正确后扩增质粒。线性化质粒,保留表达元件。用显微注射法制备转基因小鼠。
1.5 鼠尾基因组的提取和基因型的鉴定
剪0.5 cm鼠尾加入500 μL裂解液(Tris-Cl,pH 8.0,EDTA 0.005 mol/L,SDS 0.1%,Protease K 20 μg/m L),55℃摇动过夜,加入300 μL饱和NaCl,颠倒混匀置冰上10 m in,10 000 g离心10 m in取上清600 μL,加等量异丙醇轻柔混匀,看到沉淀,离心,70%乙醇洗涤DNA沉淀,100 μL TE溶解DNA。取1 μg DNA进行PCR鉴定。所用反应条件与之前扩增目的基因相同。
1.6 RT-PCR
Trizol提取组织RNA,RNA经DNase I(RNase free,New England Biolab)消化处理,用相同条件行半定量PCR。用管家基因GAPDH为内对照,引物为5′-tccaccaccctgttgctgta-3′;5′-accacagtccatgccatcac-3′。调整PCR循环数,GAPDH为20,XAGE-3的扩增为28。
1.7 Brdu标记实验
使用Zymed公司Brdu标记试剂和检测试剂盒,按照试剂盒方法腹腔注射BrdU入3周龄雄性小鼠。XAGE-3转基因组和对照组各3只。2 h后处死动物取睾丸,中性甲醛固定72 h以上,石蜡包埋切片。按照试剂盒方法进行BrdU抗体染色。
2 结果
2.1 XAGE-3基因是灵长类种属特异基因
Taxp lot是基于不同物种蛋白序列的比对结果分析某一基因在不同物种同源性的一个数据库工具。图1A(彩插1)为将人类基因组(Homo sapiens,ID9606)同小鼠基因组(Mus musculus,ID10090)和大鼠基因组(Rattus norvegicus,ID10116)的比较图。每一点代表某一基因的同源性,坐标值为相似度。XAGE-3基因的位点为红点所示,在图1A位于0点附近(箭头a所指处),确定为人类的XAGE-3基因在小鼠和大鼠的基因组中的同源性较低。图1B(彩插1)为人类基因组和近亲黑猩猩基因组(Pantroglodytes,ID9598)猕猴基因组(Macaca mulatta,ID9544)比较,XAGE-3的相似度在500以上(箭头b所指处)。这说明XAGE-3基因在灵长类存在同源性基因,而在啮齿类中并不存在,和啮齿类相比是灵长类的特有基因。
为了确证XAGE-3基因的同源性,我们在核酸序列和蛋白序列水平使用NCBI中的程序Blastn和Blastp分别比对了小鼠的基因组,无同源信息。就目前的基因组学信息和比较方法,灵长类种属特异基因XAGE-3在小鼠的基因组中无同源性序列。
2.2 目的基因的克隆和载体的构建
我们从胎盘组织、正常人外周血、人肿瘤细胞系扩增目的基因,只在正常胎盘组织中扩出目的基因,这一点和EST数据库中的信息相吻合。扩出的XAGE-3基因的开放读框通过Kpn I和EcoR I酶切位点连入真核表达载体pCDNA3.1(+),经Bgl II线性化后,显微注射制备转基因鼠。见图2。
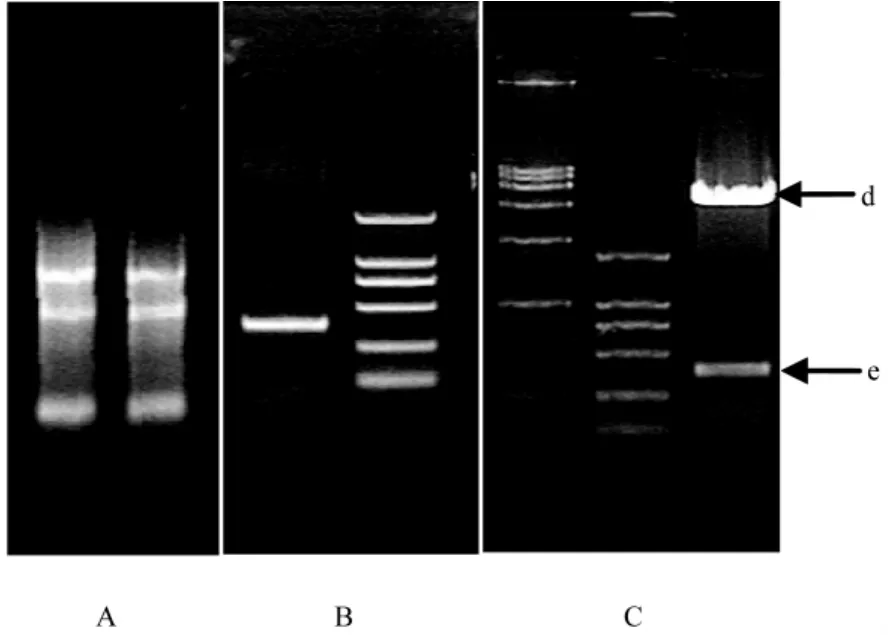
注:A.胎盘组织提取的总RNA;B.为扩增的目的基因,左为目的条带,大小为373 bp,左为DNA分子量标准DL2000 (Takara);C.为酶切结果,箭头d指向切出的载体,e箭头指向插入片段。两个分子量标准分别为DL15000和DL2000(Takara)图2 XAGE-3分子克隆和载体构建Note:A Shows the total RNA from placenta;B shows the amplified XAGE-3 at 373 bp,the marker is DL2000(Takara); C shows the identification of constructed vector,arrow d shows the linearized vector,arrow e shows the DNA inserted fragment,The markers are DL15000 and DL2000(Takara).Fig.2 Cloning of XAGE-3 and expression vector construction
2.3 动物基因型的确定
XAGE-3转基因鼠鼠尾DNA通过PCR基因型鉴定,共得到5个首建鼠,趾号1、13、16、25、34。确定为F0代。F0代动物和野生型C57BL/6J杂交,得到F1代动物,鼠尾DNA经PCR鉴定,5个品系均有阳性,阳性比率约为50%,部分动物传代至F2,F3。见图3。
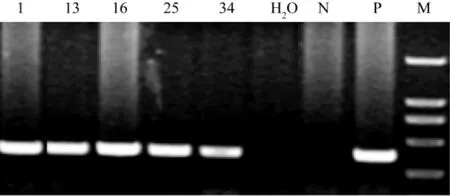
注:N为野生动物对照,P为阳性对照。H2O作扩增系统对照。DNA分子量标准为DL2000(Takara)图3首建鼠基因型PCR鉴定Note:N is wild type control,P is positive control,H is water as system control.The marker is DL2000(Takara).Fig.3 PCR genotyping of the founder mice
2.4 基因表达的分析
取5个品系F1代1月龄雄性动物睾丸组织行反转录PCR,图4A中可以看出在13,16,34中目的基因有较高表达,选取13、16号动物品系分析目的基因在其他组织中的表达(图4B、C),目的基因在所选组织均有表达,表达丰度在两个品系不同组织间存在一些差异,但是在两个品系中睾丸组织目的基因的高表达一致。见图4。
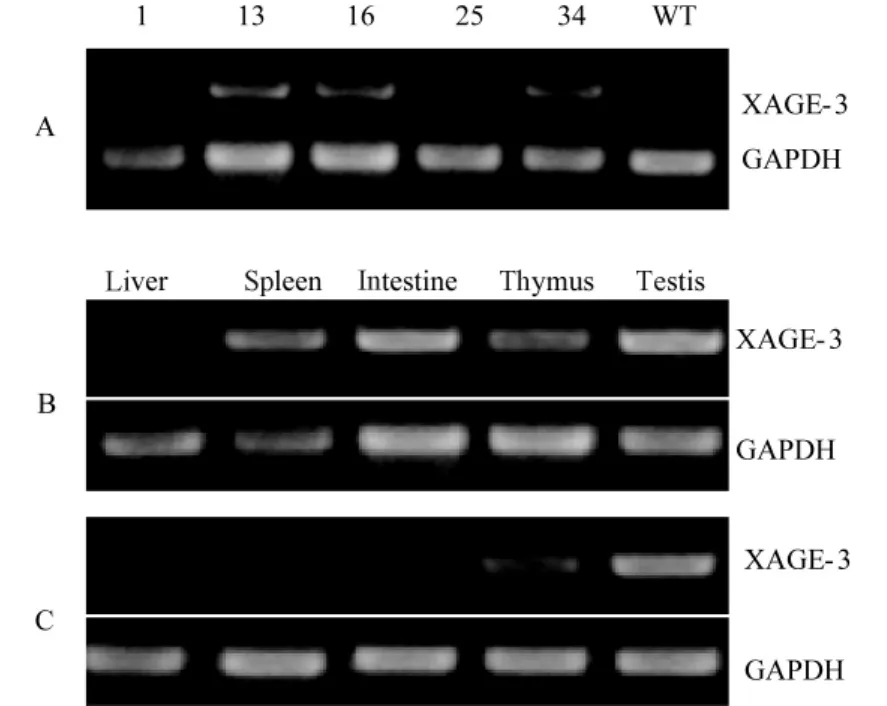
注:A.不同首建鼠睾丸组织XAGE-3表达丰度的检测,WT为野生小鼠睾丸组织样品,选择13和16号founder进行后续实验;B、C.13和16号品系XAGE-3基因不同组织表达的检测。图4 RT-PCR检测XAGE-3表达Note:A shows XAGE-3 expression in the testis in different lines,WT is wild type mouse,line 13 and line 16 were chosen for subsequent analysis;B and C show the expression pattern in different tissues in line 13 and line 16.Fig.4 XAGE-3 expression detected by RT-PCR analysis
2.5 对精母细胞的影响
在对于3周龄雄性小鼠进行Brdu标记2 h后,我们对小鼠睾丸取材,剪开包膜固定3 d以上,脱水石蜡包埋切片,切片脱水,用Brdu标记试剂盒检测被标记细胞,在图5(彩插1)中可以看出,XAGE-3转基因动物被标记的细胞有较多的处于初级精母细胞阶段(图5B,箭头所示),而非转基因对照动物被标记的细胞多为未发生减数分裂的精原细胞(图5A,箭头所示)。
3 讨论
物种的进化是一个漫长的过程。人类和小鼠作为哺乳动物他们有共同的祖先[3]。人类属于灵长类动物,而小鼠属于啮齿类,他们的生活环境不同,生活习性也存在差异。比如小鼠的繁殖方式为一胎多仔,由于生活环境较阴暗,他们进化了高度发达的嗅觉系统。而在基因水平灵长类物种的嗅觉基因有更多的处于失活状态[4]。人类和小鼠的基因组测序完整程度较高,通过两个物种的比较,人们发现,两个物种在免疫系统、生殖系统、嗅觉等方面存在更多的物种特异基因[5]。这一情况在考虑到物种生活习性等方面的差异后就容易理解了。
我们利用了NCBI的同源基因数据库Homologene对人类和灵长类物种特异基因筛查,该数据库将不同物种的基因按照同源性进行聚类,虽然这种同源性包括某一基因在同一物种内结构相似的一些基因,也有物种之间的同源。将数据库的信息按照物种(organism)先聚类,这一物种所有通过聚类分析的基因都在这一类。通过逻辑减法,将不同的物种聚类进行比较,可以找到这样的基因,它只属于被减的物种,而在另一物种该聚类基因不存在。关键词的检索也被使用。Gene exclusive to H.sapiens作为关键词检索数据库也得到一些基因,这种检索方法得到的信息不是非常严格,所有的方法得到的结果均需验证。基因组的比对(blast)是第一轮的验证。将被选基因和小鼠的基因组比对,我们研究的目的基因XAGE-3在小鼠基因组没有同源区域(结果没有显示),之后又在Taxplot平台进行分析,结果可以看出XAGE-3因在人类灵长类近亲物种同源参数大于500,而和啮齿类动物大鼠、小鼠的同源参数接近于零。这一点可以说明就目前的测序结果进行的生物信息学分析XAGE-3基因在啮齿类动物中没有发现同源序列,而在灵长类动物的3个物种该基因同源性较高。
XAGE-3因属于XAGE基因家族,因为坐落在X染色体上,在结构上和GAGE基因簇相似,故而名为XAGE。XAGE基因家族的成员XAGE1b和小细胞肺癌的发生相关[6],这一点和GAGE基因家族相似。但是XAGE-3和XAGE1b在结构上并没有太多共同点,只是因为坐落在同一染色体区,这一区域的基因被命名为XAGE-1,2,3,5等。XAGE基因家族和MAGE基因家族相似,是作为肿瘤相关基因而被关注的。这类基因主要表达于人类肿瘤组织和一些胚胎期组织以及生殖相关组织。由于其表达谱的状况,该基因也属于CT(cancer-testis)antigens家族。而这些灵长类特异基因实际上正处在高度进化的状态[7](cancer testis evolution)。在研究之初我们试图从人类肿瘤细胞系中克隆该基因,在5种不同来源的人类细胞系中并不能扩出该基因(结果未展示)。而EST数据显示该基因表达于人类胎盘组织。随后我们从志愿者的胎盘组织中扩出该基因,证实该基因在生殖系统胎盘中的表达。了解这些基因的生物学意义有助于我们对物种进化的理解。
我们制备的灵长类特异基因XAGE-3转基因小鼠实现了该基因在小鼠特别是睾丸组织的高表达。分析睾丸组织细胞分裂增殖情况,结果表明小鼠表达XAGE-3后,在3周龄时转基因小鼠有更多的细胞发育至初级精母细胞,而在非转基因动物的睾丸中更多的生精细胞处在精母细胞之前精原细胞或生殖干细胞的状态。这说明XAGE-3的表达加速了精子细胞的发生,但是这其中的机制目前我们了解甚少,对于该基因在生物体的真正意义我们尚需进一步的研究。
(本文图1、图5见彩插1。)
[1]Sayers EW,Barrett T,Benson DA,et al.Database resources of the National Center for Biotechnology Information[J].Nucleic Acids Res,2009,37(Database issue):D5-15.
[2]David AR,Garry SA,Jacques R.Visualization of comparative genomic analyses by BLAST score ratio[J].BMC Bioinformatics.2005,6:2.
[3]Ji Q,Luo ZX,Yuan CX,et al.The earliest known eutherian mammal[J].Nature,2002,416:816-822.
[4]Yoav G,Orna M,Svante P,et al.Human specific loss of olfactory receptor genes[J].Proc Natl Acad Sci U S A,2003,100:3324-3327.
[5]Waterston RH,Lindblad-Toh K,Birney E,et al.Initial sequencing and comparative analysis of the mouse genome[J]. Nature.2002,420:520-562.
[6]Nakagawa K,Noguchi Y,Uenaka A,vet al.XAGE-1 expression in non-small cell lung cancer and antibody response in patients[J].Clin Cancer Res,2005,11:5496-5503.
[7]Brian JS,Christian I,Sumir P,et al.Rapid evolution of cancer/ testis genes on the X chromosome[J].BMC Genomics,2007,8:129.
XAGE-3,A Primate Specific Gene Transgenic Research
XIANG Zhi-guang,CHEN Wei,QUAN Xiong-zhi,DONG Wei,CAO Xin-shui,GAO Shan,ZHANG Xiao-juan,ZHANG Hai-tao,Zhang Lian-feng
(Institute of Laboratory Animal Science,Chinese Academy of Medical Science and Peking Union Medical College,Key Laboratory of Human Disease Comparative Medicine,Ministry of Health,Beijing 100021,China)
ObjectiveTo assess the XAGE-3 homologous characteristics in primate and rodent species,and to analyze the XAGE-3 biological function in transgenic mice.M ethods Based on the databases Homologene and Taxplot,to compare the homologous characteristics in different genomes,to clone XAGE-3 gene from human placenta,to construct the eukaryotic expression vector pCDNA3.1(+),to create transgenic mice by microinjection,to identify the genotyping and the expression of transformed gene by genomic PCR and reverse transcription PCR,and to check the proliferation of spermatocytes in transgenic m ice and control ones by BrdU labeling.ResultsThe XAGE-3 gene had homologs in Homo sapiens,Pan troglodytes and Macaca mulatta,not in Mus musculus and Rattus norvegicus.XAGE-3 gene expression was detected in the testis in 3 of 5 transgenic lines,and in 2 lines chosen for phenotype analysis.The expression was detected in the intestine,thymus and testis.More spermatocytes were labeled by BrdU in transgenic mice compared with that in the negative control.ConlusionPrimate species specific gene XAGE-3 may affect the proliferation of spermatacytes.
XAGE-3;Species specific gene;Transgenic mice
R-394.11
A
1005-4847(2010)02-0096-04
2009-12-17
卫生部项目,实验动物和人类疾病动物模型资源扩展(200802036);十一五新药专项支持(2009ZX09501-026)。
向志光(1980-),男,博士研究生,研究方向:遗传工程。E-mail:xiangzhg@yahoo.com
张连峰,Tel:87778442,E-mail:Zhanglf@cnilas.org
