产AmpC酶大肠埃希菌中整合子的研究
2010-09-06陆思静管希周王睿梁志欣刘又宁辽宁医学院附属第一医院呼吸科锦州市11001解放军总医院军医进修学院北京市100853
陆思静,管希周,王睿,梁志欣,刘又宁(1.辽宁医学院附属第一医院呼吸科,锦州市 11001;.解放军总医院军医进修学院,北京市 100853)
产AmpC酶大肠埃希菌中整合子的研究
陆思静1*,管希周2,王睿2,梁志欣2,刘又宁2(1.辽宁医学院附属第一医院呼吸科,锦州市 121001;2.解放军总医院军医进修学院,北京市 100853)
目的:观察25株产AmpC酶大肠埃希菌中整合子的分类、结构及其在介导AmpC酶基因转移中的作用。方法:采用微量稀释法测定20种抗生素对试验菌株的敏感性。利用多重聚合酶链反应(PCR)方法检测整合酶基因(intI)及其定位,对其阳性菌株可变区(Int)扩增产物进行测序分析。结果:这25株菌对多种抗生素耐药。20株Ⅰ类整合酶基因阳性(80%);所携带的耐药基因盒绝大多数为aadA5和dfr 17;未发现携带AmpC基因盒的整合子。结论:Ⅰ类整合子广泛地存在于产AmpC酶的大肠杆菌中;耐药基因盒是整合子阳性菌株对氨基糖苷类、磺胺类药物及氯霉素耐药的主要原因,但对介导AmpC酶基因转移,不起主要作用。
整合子;产AmpC酶大肠埃希菌;基因盒;聚合酶链反应
大肠杆菌是引起临床感染最常见的细菌之一,随着多重耐药菌株的日渐增多,治疗该菌引起的感染变得越来越困难。其耐药机制非常复杂。研究表明,整合子因其不仅可介导细菌耐药性群聚,导致多重耐药性的产生,而且可以在不同遗传物质和菌种间转移,引起耐药基因高效快速转移而已经成为研究热点[1]。但国内这方面的研究鲜见。整合子由5′和3′保守末端及中间的可变序列组成,5′保守末端携带编码整合酶的基因(intI)、整合酶基因重组位点(attI)和一个启动子的基因片段。中间的可变区域携带有耐药基因的基因盒[2]。现已确认至少有9种整合子,但在耐药方面起最主要作用的为第1类整合子[3]。以解放军总医院临床连续收集的25株产AmpC酶大肠埃希菌为对象,对整合子分布及其结构特征、整合子在介导耐药转移中的作用进行研究。
1 资料与方法
1.1 菌株来源
解放军总医院微生物科从2001~2002年各种临床标本中分离得到的全部大肠杆菌719株。用三维试验、等电聚焦电泳等方法进一步筛选高产AmpC酶的菌株25株[4]。
1.2 主要试剂与仪器
DL 15 000 DNA Marker为日本TaKaRa公司产品;pGEMT载体、DNA聚合酶采用美国Promega公司产品;包被20种抗生素的96孔药敏板(天津金章医用新技术研究所);DNA Thermal Cycler 2400为美国Perkin Elmer公司产品。
1.3 药敏试验方法
采用微量稀释法测定产AmpC酶大肠埃希菌对以下20种抗生素的最低抑菌浓度(MIC):磺胺甲基异唑、庆大霉素、卡那霉素、阿米卡星、妥布霉素、奈替米星、氯霉素、四环素、亚胺培南、环丙沙星、加替沙星、左氧氟沙星、头孢噻肟、头孢他啶、头孢吡肟、头孢西丁、头孢哌酮、头孢哌酮/舒巴坦、替卡西林、替卡西林/克拉维酸。质控菌株为大肠杆菌ATCC25922。药敏判断标准按美国临床实验室标准化委员会(NCCLS)2004[5]年版的规定执行。数据统计时将中敏归于耐药分析。
1.4 整合酶基因的PCR检测
引物序列为IntIF:5′-TGC GGG TYA ARG ATB TKG ATT-3′,IntIB:5′-CAR CAC ATG CGT RTA RAT-3′(其中,Y代表C或T;R代表A或G;B代表C,G或T;K代表G或T),目的片段长度491 bp。PCR反应条件:94℃预变性10 min,然后进入循环94℃1 min,51℃30 s,72℃45 s,循环40次,最后72℃延伸10 min,在紫外灯下观察扩增产物在1.0%的琼脂糖凝胶电泳中的分析结果,出现明显亮带的为初筛阳性。阳性条带行切胶纯化后测序,用Blast程序分析。
1.5 可变区基因盒PCR检测及DNA测序分析
引物序列为Int-F:CGG ATG AAG GCA GCAACC CA,Int-R:AAG CAG ACT TGA CCT GAT AG。反应参数:94℃预变性5 min,94℃变性1 min、55℃退火1 min、72℃延伸3 min,35个循环,最后1个循环72℃延伸7 min,4℃保护。在紫外灯下观察扩增产物在0.8%的琼脂糖凝胶电泳中的分析结果,阳性条带行切胶纯化,TA克隆,转化感受态细胞DH5a,送北京奥科生物技术有限责任公司及上海基康生物技术有限公司进行测序。在Genbank比对。
2 结果
2.1 大肠杆菌中整合酶基因PCR检测及测序结果
20株产AmpC酶大肠埃希菌均在500 bp左右出现了阳性条带,提示上述菌株携带了整合酶基因,阳性率为20/25(80%),结果见图1。随机抽取的2个样品ECO03、ECO34测序,结果在Genbank上Blast为Ⅰ类整合酶。与AY463797比较序列符合率(identities)为100%。
2.2 Ⅰ类整合子可变区相关基因盒扩增结果
对20株Ⅰ类整合阳性菌株用可变区基因盒特异引物,扩增出的20个阳性PCR产物,根据片段大小分成3组:16株出现接近1 700 bp大小条带的为A组;1株出现接近2 000 bp大小条带的为B组;3株出现大于2 500 bp条带为C组。整合子可变区特异性引物PCR扩增结果见图2。在A组中随机抽取4株,B组1株、C组3株分别进行测序和序列分析。A组片段大小为1 664 bp,其含有2个开放阅读框,大小分别为474 bp和789 bp,与耐药基因盒aadA5和dfr17都为100%同源,分别对氨基糖苷类抗生素壮观霉素、链霉素和磺胺类药物甲氧苄啶产生耐药,这4株所带基因盒完全一致,Genbank登陆号分别为AY748452和AY828551。B组PCR产物测序结果表明,其长度为1 913 bp,内部插入dhfrⅫ基因(编码二氢叶酸还原酶,对磺胺类耐药)和aadA2基因(编码氨基酰腺苷酰基转移酶,对氨基糖苷类耐药),序列符合率100%,还有一编码功能不清楚的ORF5,Genbank登陆号AY748453。C组PCR产物经多次测序失败,仍在研究中。
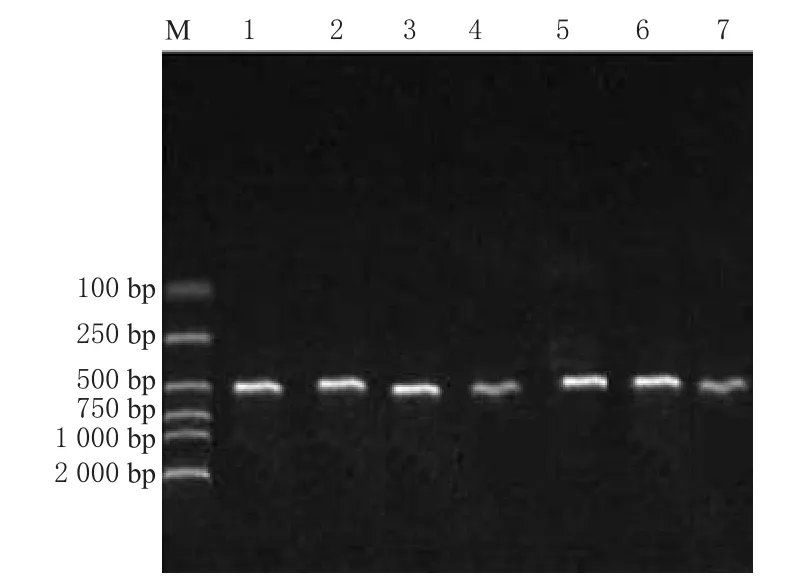
图1 整合酶基因PCR结果电泳图M.2 000 bp DNAMarker;1~7均为试验菌株Fig 1 Electrophoretogram of PCR product of intIM.2 000 bp DNAMarker;1~7.test strains
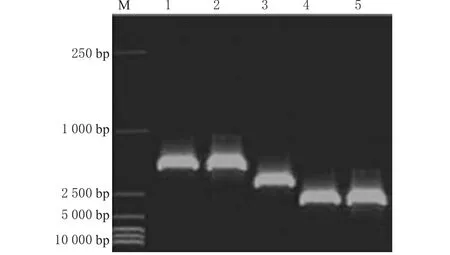
图2 整合子可变区特异性引物PCR扩增结果电泳图M.15 000 bp DNAMarker;1~2为A组;3为B组;4~5为C组Fig 2 Electrophoretogram for PCR product of specific primer in variable regions of integronsM.15 000 bp DNAMarker;1~2.groupA;3.group B;4~5.group C
2.3 20种抗生素对25株产AmpC酶大肠埃希菌的体外抗菌活性
本研究的25株大肠埃希菌对20种抗生素的敏感率由高到低的顺序依次为碳青霉烯类中的亚胺培南100%;氨基糖苷类中的奈替米星91%、阿米卡星87%、卡那霉素74%、妥布霉素70%,而头孢类仅头孢吡肟65%,其余均在50%或以下,结果见表1(表中S、I、R分别代表细菌敏感率、中介率和耐药率,其中,R=耐药菌株数/受试菌总株数,I、R的计算方法类同)。
由表1可见,大肠埃希菌对左氧氟沙星、头孢西丁的耐药率高达100%,环丙沙星95%、、四环素91%、替卡西林86%,磺胺甲基异唑、加替沙星、替卡西林/克拉维酸耐药率均达80%。
3 讨论
试验显示,25株产AmpC酶大肠埃希菌中整合子检出率为80%,均为Ⅰ类整合子,高于其他国家或地区的平均水平43%~76%左右[6],可能因为:(1)实验样本均为分离自患者的药敏测试中具有多重耐药表型的高耐药菌;(2)由于有的第Ⅰ类整合子具有不完整的3′-保守端,在试验设计上,应用了简并引物,可同时检测Ⅰ、Ⅱ、Ⅲ类整合子的分布,防止以3′-保守端扩增结果作为整合子阳性菌株的检出方法所造成漏检。
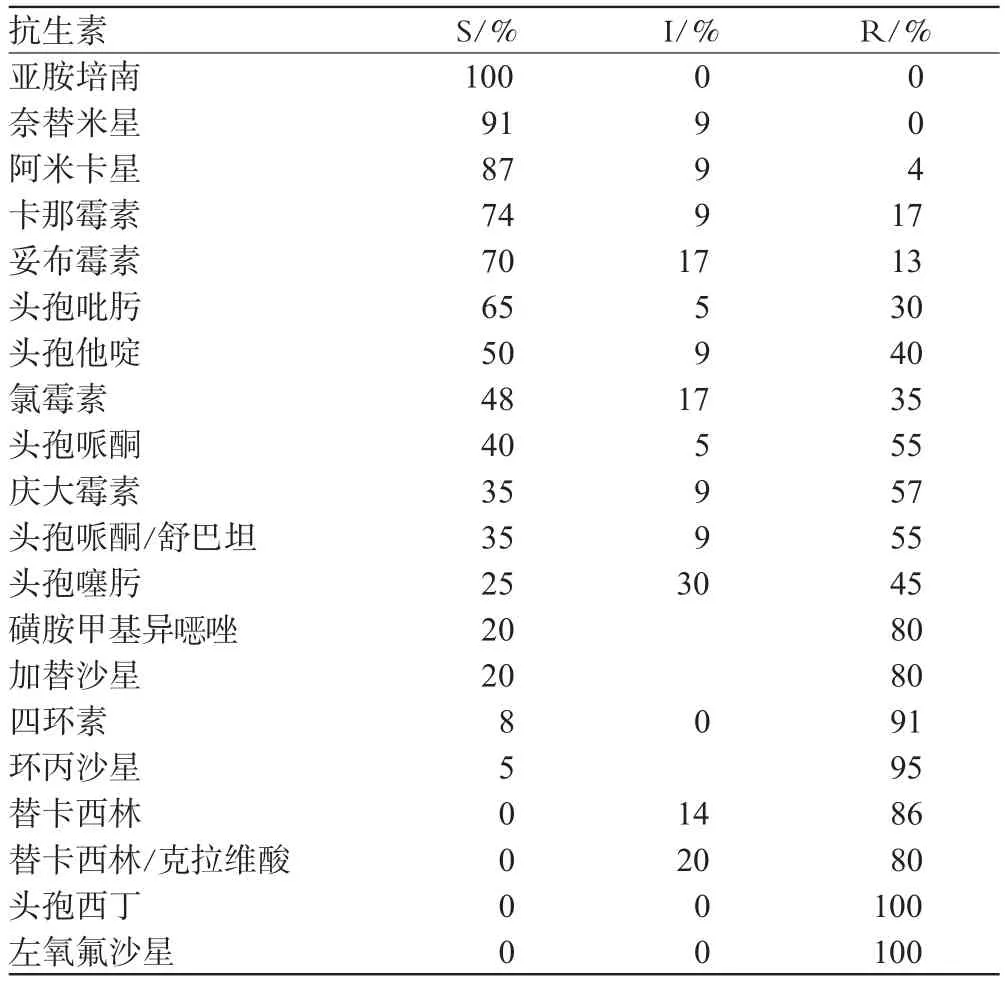
表1 大肠埃希菌对20种抗生素的药敏试验结果Tab 1 Results of susceptibility test of 20 kinds of antibiotics to Escherichia coli
研究表明,整合子所携带与耐药基因盒主要是aadA5和dfr17,aadA2和dhfrXII基因,未检测到AmpC酶基因和介导喹诺酮类耐药的基因。尽管有大量报道β-内酰胺类基因盒是整合子最常携带的基因盒之一,但本研究的25株产AmpC酶大肠埃希菌在前期试验中均发现携带产AmpC酶基因[4],至少有17株细菌整合子未携带产AmpC酶耐药基因盒,与报道不一致[7,8]。提示,在本研究的25株大肠杆菌中,耐药基因盒在介导产AmpC酶耐药转移方面,不起主要作用。
尽管这25株大肠埃希菌对包括喹诺酮类、青霉素类及酶抑制剂合剂、四环素、第3代头孢菌素及酶抑制剂合剂、磺胺甲基异唑、庆大霉素等十余种抗生素耐药,但对碳青霉烯类中的亚胺培南,敏感率仍高达100%,与有些报道[9]一致;对氨基糖苷类的阿米卡星和奈替米星仍然保持了较高的敏感率。对高产AmpC酶的大肠埃希菌引起的感染,除碳青霉烯类外,氨基糖苷类抗生素是可以选择的对象,但对治疗整合子阳性的耐药菌株,部分氨基糖苷类抗生素则无效。
值得注意的是,Verdet等[10]首次报道AmpC酶DHA-1编码基因定位于3′-保守末端整合子,此后尽管报道很少,也应引起足够重视。细菌自身染色体上的整合子-耐药基因盒可通过复制传递给下一代,即垂直传播;而整合在质粒上的则可在同菌属甚至不同菌属间水平传播。
[1]Mary GK,Jane WM,David LP,et al.Integron-mediated multidrug resistance in a global collection of nontyphoidal salmonella enterica isolates[J].Emerg Infect Dis,2009,15(3):388.
[2]Dubois V,Parizano MP,Arpin C,et al.High genetic stability of integrons in clinical isolates of shigella spp.of worldwide origin[J].Antimicrob Agents Chemother,2007,51(4):1 333.
[3]Yang CM,Lin MF,Lin CH,et al.Characterization of antimicrobial resistance patterns and integrons in human fecal escherichia coli in taiwan[J].Jpn Infect Dis,2009,62(3):177.
[4]管希周,刘又宁,罗燕萍,等.新CMY型头孢菌素酶在大肠埃希菌中的流行[J].中华医学杂志,2004,84(22):1 872.
[5]Clinical and Laboratory Standards Institute(CLSI).Performance Standards for Antimicrobial Susceptibility Testing;Sixteenth Informational Supplement[S].USA:CLSI,2005:112.
[6]Van Essen-Zandbergen A,Smith H,Veldmanl K,et al.Occurrence and characteristics of class1,2 and 3 integrons in Escherichia coli,Salmonella and Campylobacter spp.in the Netherlands[J].J Antimicrob Chemother,2007,59(4):746.
[7]Poeta P,Radhouani H,Igrejas G,et al.Seagulls of the berlengas natural reserve of Portugal as carriers of fecal Escherichia coli harboring CTX-M and TEM extended-spectrum beta-lactamases[J].Appl Environ Microbiol,2008,74(23):7 439.
[8]杨立军,朱立华,徐国宾,等.产ESBLs肠杆菌科细菌中整合子参与多重耐药机制研究[J].中国抗生素杂志,2002,27(12):722.
[9]罗宇芬,赖伟华,郭昭伟.院内产超广谱β-内酰胺酶的大肠埃希菌、肺炎克雷伯菌感染临床用药分析[J].中国药房,2008,19(26):2 036.
[10]Verdet C,Arlet G,Barnaud G,et al.A novel integron in salmonella enterica serovar enteritidis,carrying the bla-DHA-1 gene and its regulator gene ampR,originated from morganella morganii[J].Antimicrob Agents Chemother,2000,44(1):222.
Study on Integron of AmpC Enzyme-producing Escherichia Coli
LU Si-jing(Dept.of Respiratory,The First Affiliated Hospital of Liaoning Medical University,Jinzhou 121001,China)
GUAN Xi-zhou,WANG Rui,LIANG Zhi-xin,LIU You-ning(Chinese PLA Postgraduate Medical School,Beijing 100853,China)
OBJECTIVE:To investigate the classification,structure and integron-mediated AmpC enzyme gene transfer of integron in 25 strains of AmpC enzyme-producing Escherichia coli.METHODS:Sensitivity of test strains to 20 kinds of antibiotics was tested by microdilution method.PCR and sequencing were performed on test strains to identify integrase gene(intI)and its location,the product of variable region of positive integrons(Int)respectively.RESULTS:ClassⅠintI were indentified in 20 strains(80%)of 25 strains Escherichia coli.The most drug resistance box cassettes were aadA5 and dfr17 and integory encoding AmpC gene cassette was not observed.CONCLUSION:ClassⅠintegron resided in AmpC enzyme-producing Escherichia coli widely. The cause of drug resistance of integorn to aminoglycosides,sulfonamides,chloramphenicol is drug resistance gene cassette.Gene cassette does not play important role in integron-mediated antibiotic resistant gene transformation.
Integron;AmpC enzyme-producing Escherichia coli;Gene cassettes;PCR
R378.2+1;R969.3
A
1001-0408(2010)14-1293-03
2009-12-06
2010-01-24)
