短跑运动控制的生物力学分析
2010-09-06魏书涛傅维杰钟运健
魏书涛,刘 宇,傅维杰,李 庆,钟运健
短跑运动控制的生物力学分析
魏书涛1,刘 宇1,傅维杰1,李 庆2,钟运健1
目的:通过建立短跑最高速阶段下肢的生物力学模型,探讨短跑运动下肢肌肉在多关节运动中协调、控制功能的生物力学机制,为短跑技术分析、技术最佳化提供依据。方法:使用三维红外高速摄像系统(300 Hz)、三维测力台(1 200 Hz)采集8名高水平短跑运动员在塑胶跑道上全力跑动时的数据。使用环节互动动力学分析短跑最高速时一个步态周期的运动学、动力学数据。研究结果:1)触地初期,地面反作用力通过膝关节和髋关节前方,在膝、髋关节处产生伸膝、屈髋力矩的外力矩(EXF),此时肌肉力矩(MUS)的主要作用是对抗地面反作用力产生的外力矩(EXF)。此时,膝、髋关节处最大MUS分别为203.40±93.60 Nm和455.24±198.72 Nm;蹬伸末期,在髋关节处出现较大的由大腿加速度和髋线加速度产生的惯性力矩(INT),在离地后小腿后摆运动中起到积极作用;2)摆动初期,小腿的后摆主要是惯性力矩引起的;摆动末期肌肉力矩(MUS)与惯性力据(INT)出现最大值,膝关节处为249.32±38.81 Nm、194.01±30.90 Nm,髋关节处为650.81±101.06 Nm、410.80± 78.67 Nm。结论:支撑期,肌肉力矩(MUS)和地面反作用力产生的外力矩(EXF)是主要控制下肢运动的主要力矩。支撑末期,大腿加速度及髋线加速度在髋关节处产生较大的惯性力矩(INT),为离地后小腿的积极后摆提供帮助;腾空期,摆动腿的运动主要受肌肉力矩(MUS)和惯性力矩(INT)控制,其中,惯性力矩(INT)主要由小腿的角加速度产生的,两力矩相互作用,以控制与完成下肢的目标动作。
环节互动动力学;运动控制;关节力矩;短跑
1 前言
短跑运动成绩主要受运动员在腾空期下肢快速摆动能力、支撑期承受地面反作用力冲击能力以及快速产生推蹬反作用力能力的影响[13,25]。因此,针对下肢三环节间的相互作用力矩和它们在膝、髋关节处产生的影响进行研究,有利于帮助运动员和教练员了解短跑运动中支撑期与腾空期下肢肌肉协调、控制运动的功能,进一步理解人体多关节运动控制的生物力学机制,并为技术分析、技术最佳化提供依据。
环节互动动力学必须是通过对肢体运动的动力学方程的逆过程表达式进行重新整理,以分别计算主动的肌肉力和被动的反作用力的分量,据此,探讨人体运动如何受到主动力和被动力的影响,特别是运动过程中出现的肢段间的相互作用现象。它已经被应用在对闭链系统(closedlinked system)运动(如跑的支撑期)[6,12]及开链系统(openlinked systems)运动(如踢、投,跑的腾空期)[8,10,11,16,18,20]的研究中。其中,Hunter等人[12]已对短跑支撑期的肌肉力矩和外力矩进行了研究,但他们的研究局限在短跑加速阶段。Mann[14,15]等人在对短跑的研究中首次计算了肌肉力矩,发现肌肉在膝关节处产生了一个使膝关节屈的力矩,这一力矩由摆动末期一直延续到支撑初期。针对这一现象,Mann等人的解释为:在接续摆动后期的着地动作中,大腿带动小腿快速下压,膝关节屈膝力矩的作用是向后快速抓地,配合髋关节伸展带动身体向前,使身体快速通过支点正上方。此论述虽在现象上解释了这一力矩的功能,但在力学原理上无法自圆其说。因为跑步支撑阶段,身体受到重力与地面反作用力的作用,所产生的结果,一般认为会使下肢关节受到压缩,而此时如果主动肌肌肉力矩的作用为使关节屈曲,那膝关节岂不坍塌(collapse)而无法支撑身体?因此,Mann等人的解释并不能完全令人信服。此后,Mann、Putnam[14-18]等人在对短跑腾空期及踢球等开链运动模式的研究中发现,用于平衡肌肉力矩(MUS)的力矩是由运动引起的惯性力矩(INT),但研究并没有提及惯性力矩主要来自于环节运动的哪个分量,而且受试者的运动速度较低,不利于分析惯性力矩对运动控制造成的影响。另外一些研究[21,22]使用神经肌肉模型来探寻短跑下肢运动控制机理,但是这些模型研究也仅局限于腾空期。
环节互动动力学[10,11,29]主要特点与优势是可以将关节力矩区分为多个分量:关节净力矩(net joint torque,NET)、重力矩(gravitational torque,GRA)、肌力矩(muscle torque, MUS)、惯性力据(inertial torque,INT)和外力矩(contact torques,EXF)。因此,本研究的目的是,利用环节互动动力学探究短跑运动中支撑期与腾空期不同力矩在下肢运动控制中所扮演的角色[6,8-10,12,20],定量分析每个力矩在下肢关节运动中的作用与控制机制。
2 方法
2.1 实验对象
青年优秀短跑运动员8名(表1)。

表1 本研究受试者基本情况一览表 (X ±SD,n=8)
2.2 实验方法
2.2.1 实验仪器
VICON红外高速摄影系统(英国,VICON公司),摄像头为MX13,采样频率为300 Hz,软件版本为WORKSTATION 5.1,marker球直径为14 mm,专用实验服装。
KISTLER三维测力台(瑞士,KISTLER公司),型号9287B,面积600×900 mm2,内置信号放大器,采样频率为1 200 Hz,通过数模转换器与VICON系统连接并同步,牢固地安放于专门为本次实验在田径场内挖设的测力台坑内。
Visual3D三维分析软件(美国,C-MOTION公司),软件版本为Visual3D Version 3.91.1。实现对运动学和动力学数据的处理分析。
2.2.2 实验步骤
架设红外高速摄像机,拍摄范围约为9×2×2 m3,连接摄像机与VICON工作站。对VICON系统进行设置并进行拍摄范围三维空间的标定。
运动员进行充分热身,穿着专用实验服装,安放marker球,位置如图1所示。
运动员起跑位置大约距测力台40 m。运动员听从实验员的口令,全力奔跑,通过拍摄区。同时,开启实验设备,采集实验数据,每个运动员需成功采集数据两次。
2.2.3 数据处理
以右脚脚尖触地至右脚脚尖再次触地为一个步态周期,支撑期为右脚脚尖触测力台至右脚脚尖离开测力台。

图1 Marker在人体上的标定位置示意图

表2 触地初期MUS、EXF的峰值力矩及产生EXF的垂直地面反作用力一览表(X ±SD)
根据Visual3D软件建立骨架模型的要求,身体分为14个环节[7]。左右髂棘和大转子点为骨盆的近端与远端,骨盆近端与远端半径分别为左右髂棘和大转子点间距离的一半,骨盆深度为0.144 m,膝关节中心定义为胫骨内外粗隆的中点,踝关节中心定义为内外踝的中点。身体重心根据每个环节的相对近端与远端的环节重心及环节测量学参数计算获得[9]。运动学数据使用Butterworth数字滤波器进行低通滤波,截止频率为17 Hz[24,26],地面反作用力使用Butterworth数字滤波器进行低通滤波,截止频率为55 Hz。
参考Zernicke等人的环节互动动力学模型[11,17,28],自主编程计算大腿、小腿、脚三环节间的肌肉力矩及三环节间交互作用力矩,定量分析主动的肌肉力矩对下肢运动的影响,以及一个环节的运动是如何影响另一个环节的运动。在该模型中,关节的力矩包含下列分量:关节净力矩、重力矩、肌力矩、惯性力据、外力矩。其中,各力矩的关系表达式如下:
关节净力矩(NET)=重力矩(GRA)+惯性力矩(INT) +外力矩(EXF)+肌力矩(MUS)

图2 支撑期膝、髋关节处力矩变化曲线图
支撑期膝、髋关节处惯性力矩分量见图3。在支撑期,膝、髋关节处的惯性力矩主要来自于小腿、大腿及髋线加速度产生的惯性力矩。
3 研究结果
所有受试者的重心平均最大速度为9.69±0.31 m/ s,步长为1.91±0.34 m,支撑时间、腾空时间分别为0.11 ±0.01 s和0.16±0.03 s。
支撑期,膝、髋关节处的关节力矩时间曲线见图2。膝、髋关节处肌力矩MUS在触地初期(支撑期的0~5%),出现一个负的峰值,同时,地面方作用力产生的EXF出现一个正的峰值。表2为支撑期膝、髋关节处肌力矩(MUS)及外力矩(EXF)的最大值,及产生外力矩的地面反作用力(GRF)。在膝关节处,其余力矩值均相对较小,对支撑期的膝关节运动没有显著影响,但在髋关节处,惯性力矩(INT)在支撑期30%以后,其作用开始显现,先屈髋然后伸髋。

图3 支撑期膝、髋关节处惯性力矩分量时间曲线图
支撑期小腿、大腿的角速度及角加速度曲线见图4。图5为地面反作用力呈现双波峰,最大峰值为2 816.67± 321.79N,约为体重的5倍。

图4 支撑期小腿、大腿的角加速度曲线图

图5 支撑期地面反作用力曲线图
图6、图7为腾空期膝、髋关节的力矩-时间曲线(上)及惯性力矩构成分量的力矩-时间曲线(下)。肌肉力矩(MUS)与惯性力据(INT)基本成对称分布,在摆动末期出现峰值(表3),其他的力矩较小,对膝关节的运动没有显著影响。膝、髋关节处INT主要由小腿加速度产生的力矩构成(LAAT)。
图8为腾空期小腿和大腿的角度、角速度及角加速度时间曲线,在摆动末期,小腿的负向加速度最大(表4)。

表3 摆动末期膝、髋关节处MUS、INT和LAAT的峰值一览表
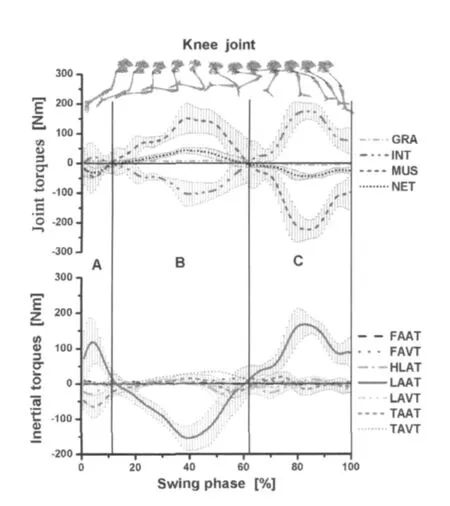
图6 腾空期膝关节处力矩及惯性力矩分量的时间曲线图
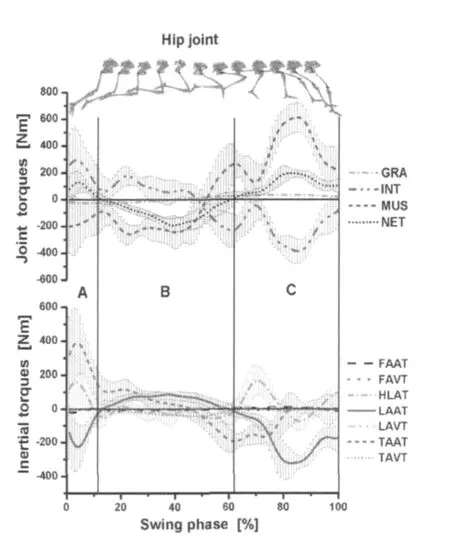
图7 腾空期髋关节处力矩及惯性力矩分量时间曲线图
表4 摆动末期大腿、小腿的角速度及角加速度峰值一览表SD)

表4 摆动末期大腿、小腿的角速度及角加速度峰值一览表SD)
角速度(rad/s)角加速度(rad/s2)Time(%)小腿11.98±1.55400.36±70.7282大腿6.46±1.3752.93±36.05

图8 腾空期小腿、大腿的角度、角速度、角加速度曲线图
4 分析与讨论
4.1 支撑期
研究结果表明,支撑阶段下肢关节起主要作用的力矩为MUS与EXF,MUS主要抵抗平衡EXF的作用(图2)。膝关节处,INT等力矩相对较小,对支撑期膝关节的运动控制没有显著影响。在触地初期(支撑期的5%),地面反作用力通过膝、髋关节前方,在膝、髋关节处产生较大的伸膝、屈髋力矩(EXF)(图2),此时,MUS的主要功能是对抗EXF,产生屈膝伸髋力矩(表2)。支撑初期(0~20%),地面反作用力的方向在20 ms左右的时间内与膝、髋关节的前后位置关系发生了连续变化(图2:A区),导致膝、髋关节处EXF变化相对剧烈,因此,在此阶段内,控制膝、髋关节运动的肌肉工作条件相对较为复杂,是下肢肌肉群损伤发生的一个高危期[3,12,19]。
在支撑末期(图2:C区),由于地面反作用力逐渐减小(图4),膝、髋关节处的EXT已趋向于零,同时,膝关节处MUS也趋向于零,但髋关节处MUS已经由伸髋力矩转变为屈髋力矩,这表明在支撑末期,髋关节处MUS已不再为下肢的蹬伸做出贡献,其主要功能是平衡髋关节处小腿、大腿及髋线角加速产生的伸髋INT(图3:A区;图4:A区),为离地后的大腿前摆(屈髋)做准备。这一惯性力矩在下肢蹬离地面后带动小腿产生顺时针运动(小腿后摆),使小腿与大腿快速折叠,减小摆动腿转动半径,使之快速前摆,帮助完成腾空期下肢运动(控制)目标。
通过计算MUS和EXT绝对值的积分,我们发现,在膝关节处,MUS的70%被用来克服EXT的作用,而在髋关节处,相对较少的MUS(62%)被用来克服EXT的作用,余下的一部分被用来对抗平衡INT的作用(特别是在支撑期的30%以后,支撑末期更为明显),以便产生髋关节角加速度。
4.2 腾空期
研究发现,摆动阶段下肢的主要力矩为MUS和INT,其他力矩(如GRA)相对较小,对下肢各环节的运动控制没有显著影响。膝、髋关节处的INT主要分量是由小腿的角加速度产生的惯性力矩(LAAT)构成。
摆动初期(0~12%),由于支撑期下肢蹬伸的作用,摆动初期重心获得较大的水平速度,同时,髋及大腿的速度变化率也最大,因此,髋关节在髋线加速度产生的惯性力矩(HLAT)及大腿角加速度产生的惯性力矩(TAAT)的作用下持续伸展(图7:A区);摆动中期(12%~62%),大腿、髋的INT迅速减弱,MUS增强,NET值为负(图7:B区)。髋关节在MUS的作用下产生屈髋运动,使大腿在腾空期的50%左右获得最大逆时针角速度(图8:D点,表4)。此后(腾空末期62%~100%)在MUS的作用下,大腿产生顺时针方向的角加速度,使大腿由逆时针方向运动(屈髋)转变为顺时针运动(伸髋),使大腿先于小腿达到最大角度(图8:E,F点),所以,小腿在后摆(屈膝)开始前,大腿已经开始下压动作(伸髋),这符合多环节运动是由近端到远端的运动模式[17],并在腾空末期(腾空期的80%~90%)MUS和INT达到峰值(表1)。此时,主要为股后肌群和臀部的伸髋肌肉群在起作用,使MUS平衡INT后,达到伸髋的目的。
赵杰等人[1,4]对短跑屈蹬技术的综述表明,屈蹬技术在支撑末期及腾空期能保持较大的肌拉力角和肌拉力矩,从而提高大、小腿折叠的力量,缩短前摆半径,提高摆动腿前摆速度。由图4和图8可见,小腿在支撑后期及离地瞬间的角速度、角加速度均为负值,表明小腿顺时针方向加速运动,使小腿与大腿快速折叠,减小摆动腿转动半径,使之快速前摆,这点与短跑屈蹬技术要求一致。但是,小腿的加速后摆运动(顺时针方向旋转)是否是由较大的肌拉力角和肌拉力矩(股后肌群)引起的呢?实验数据表明(图6),在腾空期的12%~62%,膝关节处的MUS为伸膝力矩,并非为带动小腿后摆的屈膝力矩。此时,小腿的后摆动力的主要来源是小腿和髋线加速度在膝关节处产生的屈膝惯性力矩(LAAT、HLAT)。此时MUS的主要功能是对抗INT,使小腿产生逆时针方向的角加速度,减慢小腿后摆的角速度。所以,小腿加速后摆的运动并非由屈膝肌群(股后肌群)收缩产生的,相关研究的肌电图也证明了这点[27],腾空期的0~62%股二头肌(BF)及半腱肌(ST)等使小腿后摆的双关节肌肉活动均较弱,而是由于在支撑期,运动员为获得最大的水平速度,全力后蹬,大腿、小腿和髋在离地前就已经在髋关节处产生较大的惯性力矩(图2)。因此,在腾空期(12%~62%),小腿在惯性力矩带动下后摆[2],并在摆动期的15%左右获得最大顺时针速度(图8:A点)。此后,在伸膝MUS的作用下,小腿产生逆时针方向的加速度,并于摆动期的60%左右获得最大逆时针方向的速度(图8:C点)。这与Putman[17]等人对踢、跑等动作研究得出的下肢运动过程是由近端到远端传递,由近端带动远端运动的理论模式相似。所以,高速跑动过程中,重视支撑期下肢的积极后蹬运动,使小腿、大腿及髋获得尽可能大的速度,在髋关节处产生较大的惯性力矩(INT),对腾空期小腿的快速折叠、前摆具有重要意义。
在腾空末期(腾空期60%~100%),MUS与INT为主要控制膝关节运动力矩(图6:C区)。MUS的主要功能是对抗平衡INT,该INT主要是由小腿的角加速度在膝关节处产生的(LAAT)。小腿最大角加速度(图8:B点)产生的惯性力矩占到INT值的95.77%±7.30%(表3),髋关节处,LAAT的值占INT也达到88.25%±20.28%。因此,在腾空末期,MUS的主要功能是平衡INT,而INT又主要是由小腿的加速度产生的力矩构成(图6和图7)。在腾空期的80%左右,膝、髋关节处MUS与INT达到最大值(表3)。在此阶段,股后肌群的双关节肌在伸髋、屈膝的运动起主要作用,相关研究的肌电图也表明,此时股二头肌(BF)及半腱肌(ST)活动剧烈[27],相关研究[8,12,14,15,17,19]表明,此时股后肌群被拉伸到最大长度,承受较大负荷,是股后肌群损伤发生的一个高危期。所以,腾空末期膝关节处的MUS主要功能是平衡由小腿角加速度在膝关节处产生的INT,减慢小腿前摆的速度,防止膝关节在腾空末期出现过伸的现象[14],在腾空末期,使小腿后摆,为积极着地做准备。
有关小脑功能的环节互动动力学相关研究[5,23]表明,小脑的主要作用是通过在关节产生肌肉力矩,预判由其他的环节运动在关节处产生的惯性力矩并且补偿它们,以实现精确控制人体运动的目标。短跑运动速度以及环节摆动速度非常快,其所产生的惯性力矩极其明显,说明短跑运动员的协调能力、特别是小脑功能既预测与协调惯性力矩的能力非常强。通过本研究的结果,我们可以推论,短跑运动员腾空期下肢的快速摆动可能是通过小脑产生的一个动作控制策略来完成的,通过反复练习(既学习)与适应,该策略预测惯性力矩的大小与方向,控制下肢肌肉群的活动产生肌肉力矩,该力矩的主要作用是平衡其他关节运动产生的惯性力矩,以实现运动的目标。
5 结论
支撑期,肌肉力矩(MUS)与外力矩(EXF)为控制下肢运动的主要力矩,肌肉力矩(MUS)的主要作用是抵抗平衡外力矩(EXF)的作用。触地初期,地面反作用力通过膝、髋关节前方,在膝、髋关节处产生较大的伸膝、屈髋力矩(EXF),此时,MUS产生屈膝伸髋力矩,对膝关节屈肌、髋关节伸肌-股后肌提出较高要求,是股后肌损伤发生的高危险期。而在支撑末期(80%~100%),膝、髋关节处外力矩(EXF)趋向于零,此时,肌肉力矩已不再为推动人体前进做贡献,髋关节处肌肉力矩(MUS)的主要功能是平衡大腿角加速度及髋线加速度在髋关节处产生的惯性力矩(INT),该力矩有利于小腿在离地后的后摆动作。
腾空期,肌肉力矩(MUS)的主要作用则是抵抗平衡惯性力矩(INT)的作用。在腾空前期(10%~60%),小腿在惯性力矩(INT)的作用下做后摆运动。在腾空后期(80%左右),肌肉力矩(MUS)与惯性力矩(INT)达到最大,此阶段MUS的作用同支撑初期一样是伸髋、屈膝,说明股后肌承受较大负荷。本研究发现,惯性力矩是拉伸股后肌的力量来源,其惯性力矩(INT)主要由小腿角加速度产生的力矩(LAAT)构成,此后,MUS的主要作用是减慢小腿的前摆速度,防止膝关节过度伸展,并在腾空末期,使小腿后摆,为积极着地做准备。
[1]黄达武.短跑技术理论研究热点综述[J].体育学刊,2009,16 (3):77-82.
[2]王保成,周志雄.短跑技术专门练习的创新与教学训练效果的实验比较研究[J].体育科学,2001,21(4):46-49.
[3]魏书涛,刘宇,钟运健,等.短跑支撑期股后肌损伤的动力学分析[J].中国运动医学杂志,2009,28(6):639-643.
[4]赵杰.后蹬式与屈蹬式短跑技术分析[J].武汉体育学院学报, 1995,29(1):63-65.
[5]BASTIAN A J,MARTIN T A,KEATINGJ G,et al.Cerebellar ataxia:abnormal control of interaction torques across multiple joints[J].J Neurophysiol,1996,76:492-509.
[6]CROSBIEJ,HERBERT R D,BRIDSON J T.Intersegmental dynamics of standing from sitting[J].Clinical Biomechanics,1997, 12:227-235.
[7]HAY J G.The Biomechanics of Sports Techniques[M].Englewood Cliffs,NJ:Prentice-Hall,Inc.,1993.
[8]HEIDERSCHEIT B C,HOERTH D M,CHUMANOV E S,et al.Identifying the time of occurrence of a hamstring strain injury during treadmill running:a case study[J].Clinical Biomechanics, 2005,20:1072-1078.
[9]HINRICHS R N.Adjustments to the segment center of mass proportions of Clauser et al.(1969)[J].J Biomech,1990,23:949-951.
[10]HOY M G,ZERNICKE R F.Modulation of limb dynamics in the swing phase of locomotion[J].J Biomech,1985,18:49-60.
[11]HOY M G,ZERNICKE R F.The role of intersegmental dynamics during rapid limb oscillations[J].J Biomech,1986,19: 867-877.
[12]HUNTER J P,MARSHALL R N,MCNAIR P J.Segment-interaction analysis of the stance limb in sprint running[J].J Biomech,2004,37:1439-1446.
[13]LIU Y.Kinematik,dynamik und simulation des leicht-athletischen sprints:Kinematics,dynamics and simulation of sprint running[D].Verlag Peter Lang:Frankfurt am main,1993.
[14]MANN R V.A kinetic analysis of sprinting[J].Med Sci Sport Exe,1981,13:325-328.
[15]MANN R V,SPRAGUE P.A kinetic analysis of the ground leg during sprint running[J].Res Q Exe Sport,1980,51:334-348.
[16]PUTNAM C A.Sequential motions of body segments in striking and throwing skills:descriptions and explanations[J].J Biomech,1993,26:125-135.
[17]PUTNAM C A.A segment interaction analysis of proximal-todistal sequential segment motion patterns[J].Med Sci Sports Exe,1991,23:130-144.
[18]PUTNAM C A,DUNN E G.Performance variations in rapid swinging motions:effects on segment interaction and resultant joint moments[A].Jonsson B.Biomechanics X-B Human Kinetics[M].Champaign,IL,1987:661-665.
[19]SCHACHE A G,WRIGLEY T V,BAKER R,et al.Biomechanical response to hamstring muscle strain injury[J].Gait Posture,2009,29:332-338.
[20]STIEF F,KLEINDIENST F I,WIEMEYER J,et al.Inverse dynamic analysis of the lower extremities during nordic walking,walking,and running[J].J Applied Biomechanics,2008, 24:351-359.
[21]THELEN D G,CHUMANOV E S,BEST T M,et al.Simulation of Biceps Femoris Musculotendon Mechanics during the Swing Phase of Sprinting[J].Med Sci Sport Exe,2005,37: 1931-1938.
[22]THELEN D G,CHUMANOV E S,SHERRY M A,et al.Neuromusculoskeletal models provide insights into the mechanisms and rehabilitation of hamstring strains[J].Exe Sport Sci Rev, 2006,34:135-141.
[23]WICKEL GREN I.The cerebellum:the brain’s engine of agility [J].Sci,1998,281:1588-1590.
[24]WINTER D A.Biomechanics and motor control of human movement[M].Hoboken,NJ:John Wiley and Sons.Inc,2009. [25]WOOD G.Biomechanical limitations to sprint running[J].Med Sport Sci,1987,25:58-71.
[26]YU B.Determination of the optimum cutoff frequency in the digital filter Data smoothing procedure[R].Proceedings ofXII International Congress of Biomechanics.Los Angeles,CA,1989.
[27]YU B,QUEEN R M,ABBEY A N,et al.Hamstring muscle kinematics and activation during overground sprinting[J].J Biomech,2008,41:3121-3126.
[28]ZERNICKE R F.Biomechanical Insights into neural control of Movement[A].ROWELL LB,SHEPHERD J T.Handbook of Physiology[M].New York:Oxford University Press,1996: 293-330.
[29]ZERNICKE R F,SCHNEIDER K,BUFORD J A.Intersegmental dynamics during gait:implications for control[A].PATLA AEE.Adaptability of Human Gait:Implications for the Control of Locomotion.Amsterdam[M].Elsevier Science Publishers, 1991:187-202.
Biomechanical Analysis on Movement Control during Sprint Running
WEI Shu-tao1,LIU Yu1,FU Wei-jie1,LI Qing2,ZHONG Yun-jian1
The purpose of this study was to analyze the movement control of the lower extremity by using biomechanical model during sprint running and provide the basis for technical analysis and optimization of sprint running.Methods:Three-dimensional video graphic and ground reaction force(GRF)data were collected from 8 outstanding male sprint runners performing sprint running at their maximum effort.The inter-segmental dynamics was quantified and the kinematics,the ground reaction force were analyzed during a running gait cycle of the maximal speed phase in sprinting.Results:During the initial stage of stance phase,the ground reaction force passes through in front of knee and hip joint,produces a knee extension torque and hip flexion torque.To counteract the torque of GRF,the hamstring muscle contracted and produced a flexion torque at knee(203.40±93.60Nm)and an extension torque at hip(455.24± 198.72 Nm).At the hip joint,The INT due to thigh angular acceleration and hip velocity has contributions to back swing of leg during later stance phase.During initial swing phase,The INT due to leg and thigh angular acceleration and hip acceleration has contributions to back swing of leg.The MUS and the INT are the dominating torques and their maximal values were 249.32±38.81Nm,194.01±30.90Nm at knee joint and 650.81±101.06Nm,410.80± 78.67Nm at hip joint respectively during later swing.Conclusions:During stance phase,the muscle torques and the ground reaction torques are the main torques that tends to counteract each other for controlling the movement.The INT due to thigh angular acceleration and hip velocity has contributions to back swing of leg during later stance phase.During swing phase, segment movement is controlled mainly by active muscle torques and motion-dependent torques (due to leg angular acceleration),and their functions were opposite and canceled out.
segment-interaction;movement control;joint torque;sprint running
G804.6 文献标识码:A
1000-677X(2010)09-0037-07
2010-05-30;
2010-08-30
国家自然科学基金项目(30871210);上海科委项目(08490512700);上海市“东方学者基金”资助项目。
魏书涛(1980-),男,山东临沂人,在读博士研究生,主要研究方向为运动生物力学,Tel:(021)51253239,E-mail: qfwst@126.com;刘宇(1959-),男,河北张家口人,教授,博士,德国科隆体育大学博士后,主要研究方向为运动生物力学的理论与方法、体育工程,Email:yuliu@sus. edu.cn;傅维杰,男,浙江嘉善人,在读博士研究生,主要研究方向为运动生物力学。
1.上海体育学院运动科学学院,上海200438;2.清华大学体育部,北京100084 1.Shanghai University of Sport,Shanghai 200438,China;2.Tsinghua University,Beijing 10084,China.
