紫花牡荆素体外抑制人宫颈癌HeLa细胞增殖的研究
2010-08-14谢晶白军盛习锋曹建国谢宛玉
谢晶 白军 盛习锋 曹建国 谢宛玉
1.南华大学第一附属医院妇产科,湖南 衡阳 421001;2. 湖南师范大学药物工程实验室,湖南 长沙 410013
紫花牡荆素(Casticin)是一种从蔓荆子(Vitex trifolia L)中提取的具有广泛药理活性的多甲基黄酮化合物。有研究表明Casticin具有抗炎[1]、抗性激素[2]和抗肿瘤作用[3]。近年来,有文献报道Casticin对多种肿瘤细胞有诱导凋亡和抑制增殖的作用,但其对人宫颈癌HeLa细胞的作用尚缺乏文献资料。本研究检测了Casticin 抑制人宫颈癌HeLa细胞增殖的效应,并进一步选取细胞G2/M期运行的关键蛋白P21和Cyclin B1为研究对象,探讨Casticin诱导HeLa细胞增殖的细胞分子生物学机制。
1 材料和方法
1.1 实验材料 Casticin由湖南师范大学药物工程实验室提取、分离、纯化获得。人宫颈癌 HeLa细胞株购自中南大学湘雅医学院细胞中心。RPMI 1640(Gibco公司);新生小牛血清(杭州四季青公司);DMSO(Amresco公司);MTT(Sigma公司);多西他赛(Docetaxel,恒瑞医药有限公司);兔抗P21单克隆抗体、兔抗Cyclin B1多克隆抗体(Beyotime公司)。
1.2 实验方法
1.2.1 细胞培养 用含10%小牛血清的RPMI 1640培养基,置37 ℃、CO2体积分数为5%的饱和湿度条件下培养。细胞贴壁生长,取对数生长期细胞用于实验。
1.2.2 MTT比色测定 细胞以每孔180 μL(含5×103个细胞)接种于96孔培养板中。培养24 h,待细胞贴壁后加入20 μL含受试物(1、2、4 μg/mL Casticin)和对照药品(0.1 μg/mL 多西他赛)的培养基。每组设5个复孔,分别作用24、48、72 h,每孔加入MTT(5 mg/mL)20 μL继续培养6 h,吸弃培养基,加入100 μL DMSO,振荡10 min使紫蓝色沉淀充分溶解,用酶标仪(ELX-800)以570 nm波长检测吸光度(A) 值[4],计算相对细胞活性抑制率(IR):IR=(1-A实验组均值/A空白对照组均值)×100%。按改良寇氏法求出IC50值。以上实验重复3次。
1.2.3 平皿克隆法检测 取对数生长期的细胞以每孔2×102个细胞接种于24孔培养板,培养4~6 h待细胞贴壁后,分为6组(每组设4个复孔)加入药物,分别为:NS空白对照组,0.2%DMS溶媒对照组,0.1 μg/mL多西他赛阳性药物对照组,1、2、4 μg/mL Casticin实验组。继续培养7 d后终止培养,甲醇固定,Giemsa染色,显微镜下计数大于50个细胞的克隆数[5],计算克隆形成抑制率:克隆形成抑制率(%)=(1-给药组平均克隆数/NS对照组平均克隆数)×100%。以上实验重复3次。
1.2.4 碘化丙啶染色FCM分析 取1%小牛血清同步化处理24 h的HeLa细胞,用含受试物和对照药品的10%小牛血清培养基继续培养48 h,收集细胞,用冰PBS洗2遍,用4 ℃70%的乙醇固定24 h后,碘化丙啶4 ℃避光染色[6], 流式细胞仪检测。以上实验重复3次。1.2.5 Western blot蛋白印迹检测 用终浓度为1、2、4 μg/mL的Casticin 溶液分别处理HeLa细胞48 h后,用冰PBS洗3次,加入细胞裂解液提取蛋白[3],用BCA蛋白定量试剂盒测定蛋白浓度,取30 μg样品用 SDS-PAGE电泳分离,然后将蛋白转移至PVDF膜上,再用5%脱脂牛奶-TBST室温摇床封闭2 h,一抗于37 ℃下温育3 h,二抗于37 ℃下温育1 h,ECL发光剂激发荧光,压片显影定影。结果用灰度扫描仪处理分析。
2 结 果
2.1 Casticin对人宫颈癌 HeLa细胞活性的影响 MTT比色测定的结果显示,与空白对照组相比,在Casticin(1、2、4 μg/mL)在24、48、72 h时对HeLa细胞都有明显的抑制作用(P<0.05);Casticin作用48 h时,不同药物浓度组对HeLa细胞的增殖抑制率差异有统计学意义(P均为0.000);多西他赛阳性药物对照组对HeLa细胞的增殖抑制率与空白对照组相比,差异有统计学意义(P=0.000),与Casticin 4 μg/mL相接近,但差异有统计学意义(P=0.043);Casticin抑制HeLa细胞活性,呈时间和剂量依赖性,其中Casticin作用48 h的IC50值为2.82 μg/mL(图1)。
2.2 Casticin对人宫颈癌 HeLa细胞集落形成的影响 Casticin经1、2、4 μg/mL的作用7 d后,HeLa细胞克隆形成数逐渐减少,克隆形成率逐渐下降,HeLa细胞的集落抑制率分别为18.62%、43.16%和72.24%,明显低于空白对照组(P均为0.000 ),表明Casticin对体外培养人宫颈癌HeLa细胞克隆形成具有抑制作用,呈浓度依赖性;溶媒对照组与空白对照组比较,差异无统计学意义(P=0.397);多西他赛阳性药物对照组对HeLa细胞克隆形成抑制率大于Casticin 4 μg/mL组,但两组相比差异无统计学意义(P=0.056,表 1,图2)。
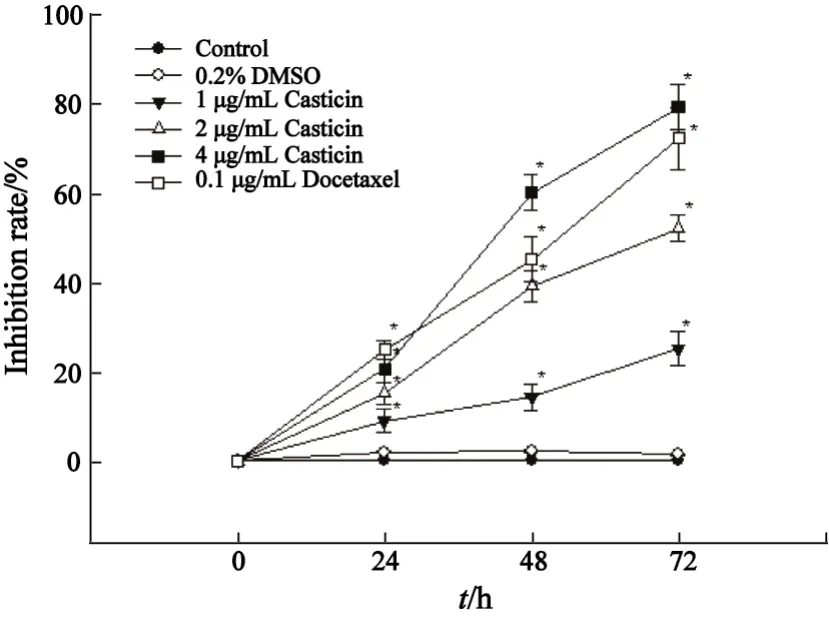
图1 不同浓度Casticin对HeLa细胞增殖活性抑制率Fig.1 Effect of Casticin with different concentrations for different time on proliferation of HeLa
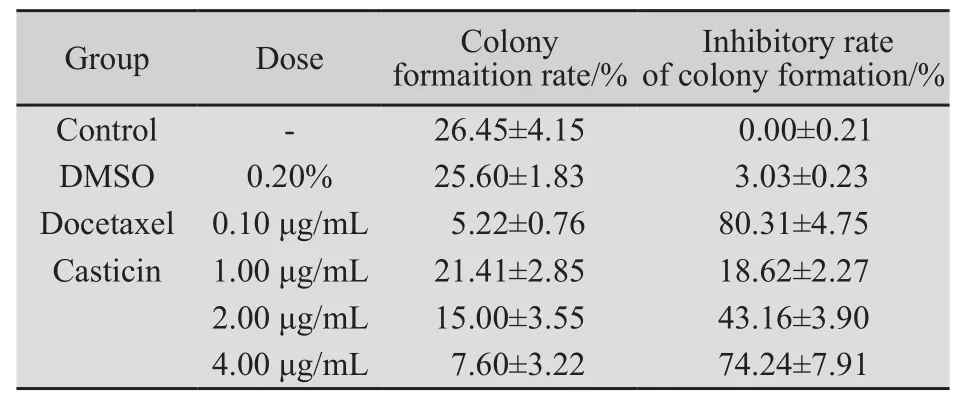
表1 Casticin对HeLa细胞集落形成的影响Tab.1 Effect of Casticin with different concentrations on the colony formation of HeLa cells
2.3 Casticin对人宫颈癌 HeLa细胞周期的影响 PI染色流式细胞术分析结果表明,Casticin(1、2、4 μg/mL)处理HeLa细胞48 h,细胞明显被阻滞于G2/M期。Casticin各处理组与空白对照组之间比较,差异有统计学意义(P均为0.000);Casticin各处理组之间比较差异亦有统计学意义(P<0.05);多西他赛阳性药物对照组与空白对照组之间比较,差异有统计学意义(P=0.000);多西他赛阳性药物对照组对HeLa细胞的G2/M期的阻滞率与Casticin 2 μg/mL组相近,两者相比差异无统计学意义(P=0.056,表2,图3)。
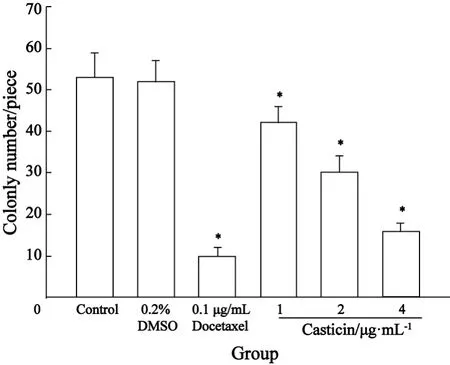
图2 Casticin对HeLa细胞集落形成的影响Fig.2 The effect of Casticin on colony formation of HeLa cells
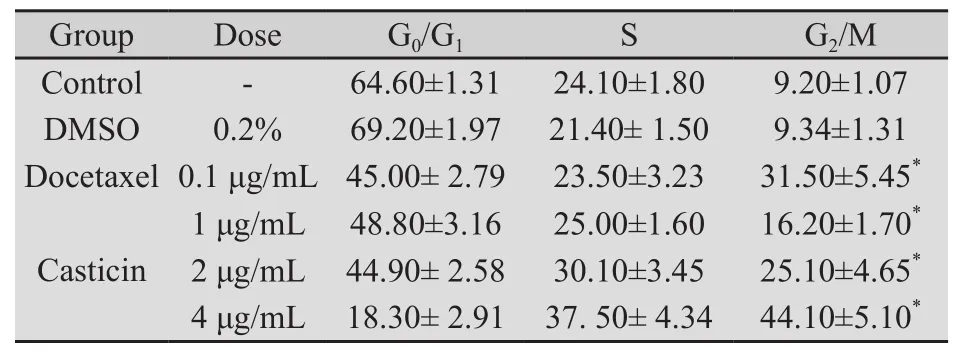
表2 Casticin作用HeLa细胞48 h对细胞周期的影响Tab.2 Effect of Casticin with different concentrations for 48 h on cell cycle of HeLa cells
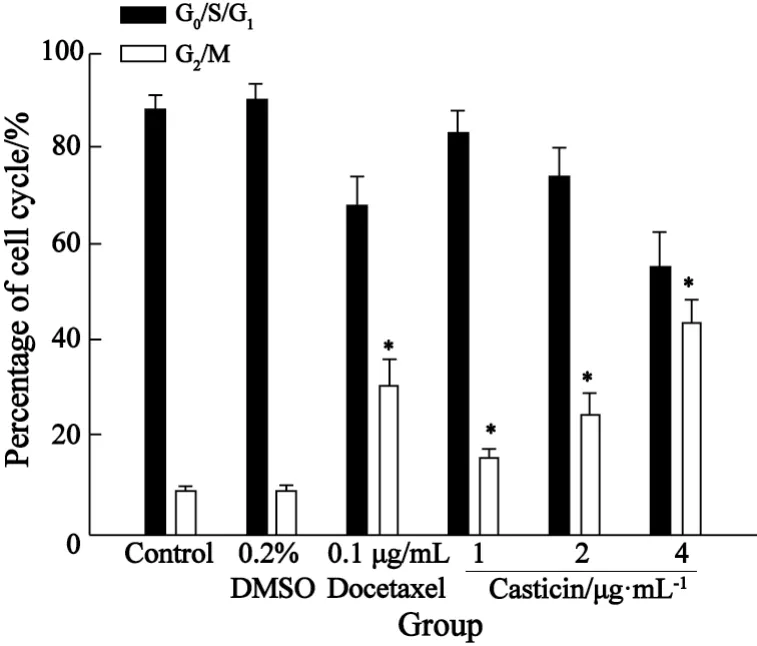
图3 Casticin作用HeLa细胞48 h后对细胞周期的影响Fig.3 Effect of Casticin with different concentrations for 48 h on cell cycle of HeLa cells
2.4 Casticin对HeLa细胞P21和Cyclin B1蛋白表达的影响 Western blot检测结果显示:经Casticin(1、2、4 μg/mL)处理 HeLa细胞48 h后,P21蛋白表达上调,呈剂量依赖性,与空白对照组相比,差异有统计学意义(Pcontrolvs1 μg/mL Casticin=0.031、Pcontrol vs 2 μg/mL Casticin=0.000、Pcontrolvs4μg/mLCasticin=0.004);Cyclin B1蛋白表达呈剂量依赖方式下调,Casticin 1 μg/mL与空白对照组相比,差异无统计学意义(P=0.767),Casticin 2 μg/mL、 Casticin 4 μg/mL与空白对照组差异有统计学意义(Pcontrol vs 2 μg/mL Casticin=0.022、Pcontrol vs 4 μg/mL Casticin=0.000,图4)。
3 讨 论
Casticin是一种从蔓荆子中提取的具有广泛药理活性的多甲氧基黄酮类化合物,是蔓荆子的主要成分,研究表明Casticin对正常细胞增殖无影响或影响较小,对人白血病、乳腺上皮癌、口腔鳞状细胞癌、人结肠癌、人大肠癌、人肺癌、人纤维肉瘤、前列腺癌等癌细胞株的增殖具有抑制活性[3,6-7]。Kobayakawa等[7]报道Casticin对人表皮癌KB细胞的生长有显著抑制作用,认为抑制微管蛋白的聚合作用而破坏有丝分裂纺锤体结构,使其阻滞于G2/M期。Haidara等[6]通过研究Casticin对乳腺上皮癌MCF-7细胞增殖的影响,发现Casticin通过作用于多个靶标分子来启动细胞周期G2/M期阻滞和诱导细胞凋亡。G2/M与微管蛋白结合而干扰有丝分裂纺锤体结构的形成,导致G2/M期阻滞。而微管蛋白的破坏会诱导P21的形成,从而抑制CDK1活性,阻滞细胞于G2/M期。同时Casticin对Cyclin A的直接下调作用参与了CDK1活性抑制,使Casticin不依赖P53而发挥抑制增殖活性。研究还发现Bcl-2的下调与凋亡前蛋白Bax上调相关,提高Bcl-2/Bax两者的比例有利于促进诱导凋亡作用。王海燕等[8]研究认为Casticin抑制白血病k562细胞株增殖的机制为激活了线粒体调控的凋亡途径,而Shen等[3]通过对K562细胞研究发现,联合使用PI3K/AKT特异性抑制剂能够增强Casticin的抗肿瘤效果。目前有关Casticin抑制细胞的增殖是独立的过程还是细胞有丝分裂毁坏的结果的问题仍未清楚。
本研究选取了人宫颈癌HeLa细胞为研究对象,检测了Casticin对HeLa细胞的增殖抑制作用及其对细胞周期的阻滞效应,并进一步检测其分子生物学机制。Cyclin B1是一种重要的周期蛋白,而细胞G2/M期的调节有赖于Cyclin B1/CDC2(CDK1的催化亚基)复合物的生成,复合物的形成增加可使细胞通过G2/M检测点,从而引发细胞周期的运行,因此Cyclin B1的增高与降低都会影响细胞周期的运行[9-10]。本研究结果显示:不同浓度Casticin作用于HeLa细胞后,Cyclin B1蛋白表达水平随药物浓度的升高而降低,说明细胞的增殖抑制可能与下调Cyclin B1表达,影响Cyclin B1/CDC2复合物的生成,以及负调节G2/M期运行有关,这与Kobayakawa等[7]和Haidara等[6]的报道结果相似。
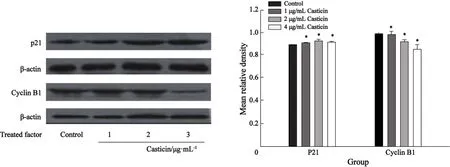
图4 Western Blot检测Casticin作用HeLa细胞48 h后蛋白表达变化Fig.4 The expression of proteins of HeLa cells treated with Casticin for 48 h (analyzed by Western blot)
P21是周期蛋白CDK1抑制物[11-12],它有两个独特的功能区,其中N-端部分的残基的功能是介导CDK1的阻断效应,抑制周期蛋白CDK1活性的表达,从而减少Cyclin B1-CDK1复合物的生成及其活性,影响细胞周期进程,进而使细胞阻滞于G2/M期[13],从而影响细胞的增殖。本实验结果显示不同浓度的Casticin作用于HeLa细胞后增殖受到抑制,P21蛋白表达上调,与Shen等[3]和Haidara等[6]报道的Casticin作用于K562细胞后,P21蛋白表达上调结果相似。
综上所述,Casticin抑制HeLa细胞增殖可能是通过活化P21蛋白表达,降低Cyclin B1蛋白表达来实现的。其对HeLa细胞的半数抑制浓度(IC50值)的影响在微克每毫升水平,其作用明显弱于临床常用生物碱类化疗药物多西他赛,但是Casticin是从传统中药蔓荆子中提取的多甲基黄酮类化合物,对人体正常细胞无生长抑制作用,而对肿瘤细胞表现出较强的生长抑制作用,其对细胞的选择性强于多西他赛[6-7]。本研究通过对Casticin抑制HeLa细胞增殖效应以及分子生物学机制的探讨,提示它可能是治疗宫颈癌等恶性肿瘤的有效化疗药物,具有较大的开发价值,但目前的研究还仅限于体外实验,其在体内对细胞的影响还有待于进一步研究。
[1] Shan L, Hong Z, Ting H, et al. In vivo effect of casticin on acute inflammation[J]. J Chin Integr Med, 2007, 5(5): 573-576.
[2] Hu Y, Xin HL, Zhang QY, et al. Anti-nocieptive and antihyperprolactinemia activities of fructus viticis and its effective fractions and chemical constituents[J]. Phytomedicine,2007, 14:668-674.
[3] Shen JK, Du HP, Yang M, et al. Casticin induces leukemic cell death through apoptosis and mitotic catastrophe[J].Ann Hematol, 2009, 88(8): 743-752.
[4] Hu Y, Hou TT, Zhang QY, et al. Evaluation of the estrogenic activity of the constituents in the fruits of Vitex rotundifolia L.for the potential treatment of premenstrual syndrome[J]. J Pharm Pharmacol, 2007, 59(9): 1307-1312.
[5] 赵环宇, 张维铭, 陈锦飞. X线照射联合RNA干扰STAT3基因对人食管癌细胞增殖凋亡的影响[J].中国癌症杂志,2009, 19(11): 836-841.
[6] Haidara K, Zamir L, Shi QW, et al. The flavonoid Casticin has multiple mechanisms of tumor cytotoxicity action[J].Cancer Lett, 2006, 242(2): 180-190.
[7] Kobayakawa J, Nishimori FS, Moriyasu M, et al .G2-M arrest and antimitotic activity mediated by casticin, a flavonoid isolated from Viticis Fructus[J]. Cancer Lett, 2004,208(1):59-64.
[8] 王海燕, 蔡兵, 崔承彬, 等. 蔓荆子活性成分vitexicarpin诱导K562细胞凋亡的机制 [J]. 药学学报, 2005, 40 (1):27-31.
[9] Nakata Y, Shetzline S, Sakashita C, et al. c-Myb contributes to G2/M cell cycle transition in human hematopoietic cells by direct regulation of cyclin B1 expression[J]. Mol Cell Biol,2007, 27(6): 2048-2058.
[10] Suzuki T, Urano T, Miki Y, et al. Nuclear cyclin B1 in human breast carcinoma as a potent prognostic factor[J]. Cancer Sci, 2007, 98(5): 644-646.
[11] Kaler P, Sasazuki T, Shirasawa S, et al. HDAC2 deficiency sensitizes colon cancer cells to TNF alpha-induced apoptosis through inhibition of NF-kappaB activity[J]. Exp Cell Res,2008, 314: 1507-1518.
[12] Mologni L, Cleris L, Magistroni V, et al. Valproic acid enhances bosutinib cytotoxicity in colon cancer cells[J]. Int J Cancer, 2009, 124(8): 1990-1996.
[13] Buchynska LG, Nesina IP, Yurchenko NP, et al. Expression of P53, P21(WAF1/CIP1), P16(INK4A) and Ki-67 proteins in serous ovarian tumors[J]. Exp Oncol, 2007, 29(1): 49-53.
