人乳头瘤病毒18型E6蛋白真核表达载体的构建及其在子宫颈癌细胞株Hela细胞中的表达
2010-05-30王青元范少君肖卫华
卫 莹,朱 靖,王青元,范少君,李 敏,肖卫华,凌 斌,周 颖
(1.安徽医科大学附属省立医院妇产科分子实验室安徽省分子医学重点实验室,安徽合肥230001;2.中国科技大学生命科学院免疫学研究所)
人乳头状瘤病毒(human papillomavirus,HPV)持续感染是宫颈癌发生的最重要危险因素之一,高危型HPV,如HPV16和18的感染在宫颈癌中最常见,其E6基因及产物常常导致细胞基因失活、变异和异常表达,最终诱导宫颈上皮细胞永生化,是最主要的癌蛋白编码基因[1]。近年来对E6蛋白的研究逐步深入,但是E6蛋白在子宫颈癌细胞株中的表达水平很低,很难检测[2]。本文将HPV18E6基因克隆到3×flag真核表达载体上,构建重组质粒p3×flag-myc-CMV-24-HPV18E6,经鉴定后转染到Hela细胞中,为进一步研究HPV18E6在宫颈癌发病中的作用奠定实验基础。
1 材料和方法
1.1 材料 总RNA提取试剂Trizol购自Invitrogen公司,逆转录试剂盒购自TakaRa公司,DEPC购自Sigma公司,TaqDNA聚合酶购自 TakaRa公司,pMDTM18-T Vector购自TaKaRa公司,小剂量质粒提取试剂盒购自Axygen公司,限制性内切酶HindⅢ、SalⅠ及DNA连接酶均购自Fermentas公司,大肠肝菌菌株E.coli DH5α购自上海生工生物工程有限公司,脂质体转染试剂盒Lipofectamine2000、DMEM培养基、胎牛血清购自Invitrogen公司,HPV18E6抗体购自santa cruz公司,GAPDH抗体购自Cell signaling公司,flag抗体购自sigma公司,Western-blot化学发光底物购自PIERCE公司。
1.2 方法
1.2.1 p3×flag-myc-CMV-24-HPV18E6真核表达载体的构建及鉴定 参考GenBank中HPV18E6碱基序列,采用Primer Premier 5.0软件进行分析后设计出18E6上下游引物:P1:GCGCGCTTTGAGGATCCAA;P2:TTATACTTG T GTTTCTCTGCGTCG.Fw1:CCCaagctt GCGCGCTTTGAGGATCCAA(HindⅢ);Rv1:GCGTCGAC TACTTGTG TTTCTCTGCGTCG(SalⅠ);分别加入了HindⅢ、SalⅠ酶切位点(划线部分)。用Trizol法从Hela细胞中提取总RNA,以Oligod(T)18为逆转录引物,以提取的总RNA为模板,在M-mlv反转录酶催化下反转录出cDNA。取 2 μ l cDNA为模版,采用Takara Ex TaqDNA聚合酶进行PCR,扩增目的基因。反应条件:94℃5 min 1个循环,94℃30 s,55℃30 s,72℃30 s共35个循环,72℃10 min 1个循环。取15 μ l PCR产物在1%琼脂糖凝胶中电泳。依据TaKaRa胶回收纯化试剂盒操作规程,胶回收纯化目的片段,回收成功后将目的片段连接到pMD-18T载体中,转化后提取质粒测序鉴定正确后,以pMD-18T-18E6为模板,经Fw1,Rv1引物扩增得到编码区片段。将18E6编码区片段与载体分别用HindⅢ、SalⅠ酶切,胶回收获得载体片段和HPV18E6片段,然后用DNA连接酶,20℃连接过夜,将连接产物转化进DH5α感受态大肠杆菌,涂板(氨苄霉素抗性),37℃生长16 h-24 h,长出菌斑约100个,挑出14个菌斑溶于1 ml氨苄霉素抗性的LB培养基中少量摇菌12 h,取2 μ l作为模板进行PCR,有四个克隆扩增出HPV18E6片段,阳性的菌液加入氨苄霉素抗性的LB液体培养基中震荡摇菌,37℃过夜。参照Axygen质粒小量提取试剂盒操作规程提取纯化p3×flagmyc-CMV-24-HPV18E6,HindⅢ、SalⅠ酶切质粒,电泳鉴定,鉴定证实后的细菌扩增培养,提取质粒,送到上海生工生物工程有限公司测序鉴定。
1.2.2 重组质粒转染Hela细胞 将人子宫颈癌细胞株Hela种到 6孔板中,每孔2×105,完全培养基培养24 h后用DMEM 洗涤2次,分别将p3×flagmyc-CMV-24-HPV18E6质粒、p3×flag-myc-CMV-24对照质粒各 4 μ g,脂质体 Lipofectamine2000 10 μ l/孔 。转染6 h后,更换完全培养基,48 h后收取细胞。
1.2.3 蛋白的提取及Western-blot检测 收取转染细胞,冷PBS洗3次后,加入30 μ l RIPA(临用前加入PMSF、抑蛋白酶肽)冰上30 min,4℃12 000 rpm离心30 min,移上清至一新的微量离心管,蛋白定量后进行SDS-PAGE电泳,再电转到PVDF膜上,经封闭2小时后一抗 flag(1∶1 000)或 GAPDH(1∶1 000)或HPV18E6(1∶50)4℃过夜,TBS-T洗后加入相应的二抗孵育2小时,膜经TBS-T洗后与ECL发光检测剂结合,经X片压片曝光,显示结果。
2 结果
2.1 HPV18E6的PCR产物电泳结果 DNA marker为DL2000标准参照物,PCR产物琼脂糖凝胶电泳显示在500 bp的一条特异性目的片段,与所设计的HPV18E6目的基因片段大小一致(图1)。

图1 HPV18E6目的基因片段电泳图
2.2 p3×flag-myc-CMV-24-HPV18E6重组质粒酶切鉴定、测序 经HindⅢ和SalⅠ双酶切后出现分子量分别为4 700 bp和500 bp的2条带,正好与质粒p3×flag-myc-CMV-24和HPV18 E6基因的分子量一致(图 2)。将重组质粒 p3×flag-myc-CMV-24-HQV18E6送到上海生工生物有限公司测序,测序结果均与目的序列完全相同,表明构建载体成功。

图2 HPV18E6目的基因片段电泳图及p3×flag-myc-CMV-24-HPV18E6酶切鉴定
2.3 p3×flag-myc-CMV-24-HPV18E6重组质粒转染Hela细胞后,E6蛋白和flag标签的表达情况p3×flag-myc-CMV-24-HPV18E6重组质粒转染Hela细胞48 h后,Western blot检测结果显示转染组E6蛋白显著升高并可检测到flag标签(图3)。
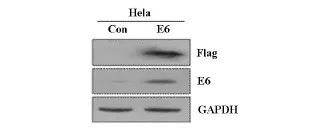
图3
3 讨论
目前,子宫颈癌发病率居高不下,且发病年龄趋于年轻化,宫颈癌发病与人乳头状瘤病毒(HPV)感染密切相关,HPV为双链环状DNA,其片段长度约为8.0 kb,编码6个早期蛋白和2个晚期蛋白,HPV通过抑制一系列的靶蛋白,如肿瘤抑制因子,避免了被感染细胞的凋亡,从而使细胞处于永生化状态[3、4]。其中 E6、E7是HPV 致癌的关键分子,在宫颈癌组织中持续表达,能促使宿主细胞进入增殖周期而导致细胞复制,在细胞恶性转化中起关键作用。HPV E6蛋白由151个氨基酸残基组成,含有2个锌指结构,每个锌指结构含有2个C-X-X-C序列,锌指结构是E6发挥功能所必需的[5]。高危型HPV E6与E7相互作用使人类的角质形成细胞永生化,并通过作用于ras致癌基因从而使细胞发生恶性转化[6]。E6蛋白是HPV感染过程中最早表达的蛋白之一,通过招募E6AP泛素酶形成复合体,然后与p53核心部分相结合,E6将活化的泛素转移到p53蛋白,p53进行泛素化降解,使p53蛋白控制的G1/S节点失去控制,导致细胞染色体不稳定和基因突变增加以及外源DNA整合到染色体中机率大大升高[7、8]。此外,E6还可与细胞内许多蛋白相互作用而协同参与细胞转化过程:如抑制宿主细胞凋亡、激活端粒酶等对宫颈癌细胞的恶性转化及细胞永生化过程有重要作用[9]。但由于E6蛋白在子宫颈癌细胞株Hela细胞中的表达水平太低,很难用Westernblotting等方法检测[2],阻碍了我们对其致癌机制的进一步研究。
随着组蛋白学的迅速发展,重组蛋白质的应用近年来大大增加,重组蛋白杂合体含有一个多肽融合担体也就是亲和标签可用于辅助目标蛋白的检测。Flag标签系统是利用一短的亲水8氨基酸肽并融合到目标蛋白,常标记于重组蛋白质的氨基端以帮助分析靶蛋白的生物学功能,并且可以用专门的单抗进行检测[10]。为了进一步提高flag标签的检测能力,目前常用3×flag标签系统,这一三联体flag抗原表位是亲水性的,由22个氨基酸组成,可以检测表达低至10 fmol水平的融合蛋白。而且研究发现使用这些很小的肽标签不会对融合蛋白质的功能及定位发生干扰。此外,含CMV启动子可以高效启动目的蛋白在细胞中的表达,进一步有助于目的蛋白的检测。
本实验中利用亲和标签flag对目的蛋白进行检测和监测的方法,将已合成的p3×flag-myc-CMV-24-HPV18E6质粒导入子宫颈癌细胞株Hela细胞中,并对其蛋白复合物进行检测,不仅提高了E6蛋白的表达量,通过flag标签系统使得E6蛋白更易于检测,为进一步研究HPV18E6蛋白及其复合物的结构功能及其在子宫颈癌中所发挥的作用奠定了基础。
[1]ZUR HAUSEN H.Papillomaviruses and cancer:from basic studies to clinical application[J].N at Rev Cancer,2002,2:342.
[2]Hall AH,Alexander KA.RNA interference of human papillomavirus type 18 E6 and E7 induces senescence in HeLa cells.J Virol,2003,77(10):6066.
[3]Woodman CB,Collins SI,Young LS.The natural history of cervical HPV infection:unresolved issues[J].Nat Rev Cancer,2007,7(1):11.
[4]Zhang Y,Dasgupta J-Ma RZ,et al.Structures of a HPV-E6 polypeptide bound to MAGUK proteins:mechanisms of targeting tumor suppressors by a high-risk HPV oncoprotein.J Virol,2007,81(7):3618.
[5]Lipari F,McGibbon GA,Wardrop E,et al.Purification and biophysical characterization of a minimal functional domain and of an N-terminal Zn2+-binding fragment from the human papillomavirus type 16 E6 protein.Biochemistry,2001,40(5):1196.
[6]Yoshida S,Kajitani N,Satsuka A,et al.Ras modifies proliferation and invasiveness of cells expressing human papillomavirus oncoproteins[J].J Virol,2008,82(17):8820.
[7]Massimi P,Shai A,Lambert P,et al.HPV E6 degradation of p53 and PDZ containing substrates in an E6AP null background.[J].Oncogene,2008,27(12):1800.
[8]FiammaM,Lawrence B.The human papillomavirus E6 protein and its contribution to malignant progression.[J].Oncogene,2001,20:7874.
[9]Nomine Y,Masson M,Charbonnierm S,et al.Structural and functional analysis of E6 oncoprotein:insightsin the molecular pathogenesis[J].Mol Cell,2006,21(5):665.
[10]Nonaka H,Fujishima SH,Uchinomiya SH,et al.FLAG-tag selective covalent protein labeling via a binding-induced acyl-transfer reaction.Bioorg Med Chem Lett,2009,19(23):6696.
