不同生境类型的华北落叶松林人工促进天然更新措施
2025-01-24王正安李颖余治家吴苗马杰佘萍贾宝光





摘要[目的]研究不同生境类型的华北落叶松人工促进天然更新措施效果,发展先进可行的人工促进天然更新技术,为提升华北落叶松人工林天然更新能力及林分质量提供技术支撑。[方法]在宁夏固原市,沿六盘山主脉不同方位选择5个林场作为试验点,建立不同生境类型的12m×12m的人工促进天然更新样地20个(林内9个,林窗6个,林缘5个)。每个样点于2021年秋季(10月)和2022春季(5月)人工撒播种子;每个试验点布设6个地表处理措施[A对照(CK)、B开沟、C挖穴、D随机小穴密点、E粗翻和F精细整地],每处理3次重复。基于连续2年的监测数据,分析确定影响天然更新的主要因素,提出人工促进天然更新措施。[结果]从连续2年各处理措施保存密度来看,在林内生境的秋播情况下表现为F(3.1株/m2)gt;E(2.8株/m2)gt;B(2.1株/m2)gt;C(1.8株/m2)和gt;D(1.1株/m2)gt;CK(0.8株/m2),春播情况下表现为F(6.4株/m2)gt;E(5.8株/m2)gt;B(5.1株/m2)gt;C(3.0株/m2)gt;D(2.0株/m2)gt;CK(1.8株/m2)。在林窗生境的秋播情况下表现为E(12.6株/m2)gt;CK(8.7株/m2)gt;B(8.1株/m2)gt;C(8.0株/m2)gt;F(7.7株/m2)gt;D(6.3株/m2);春播情况下表现为E(13.9株/m2)gt;F(12.4株/m2)gt;B(8.9株/m2)gt;CK(8.2株/m2)gt;D(7.0株/m2)gt;C(6.5株/m2)。在林缘生境的秋播情况下依次为C(1.9株/m2)gt;B(1.8株/m2)gt;E(1.4株/m2)gt;F(1.1株/m2)gt;CK(0.6株/m2)gt;D(0.3株/m2),春播情况下依次为A(1.6株/m2)gt;B(1.4株/m2)gt;E(1.0株/m2)gt;F(0.9株/m2)gt;C、D(0.5株/m2)。[结论]综合来看,在3类生境中幼苗保存密度最高的为林窗,林内和林缘次之。在播种季节上,3类生境的出苗和保存密度均为春播优于秋播。在地表处理措施中,无论是秋播还是春播,林内生境保存密度以E和F处理整地较好,林窗生境的以E和B处理较好,林缘生境以F、E和B处理较好。
关键词华北落叶松;人工促进天然更新措施;六盘山
中图分类号S754.1"文献标识码A"文章编号0517-6611(2025)02-0127-07
doi:10.3969/j.issn.0517-6611.2025.02.027
开放科学(资源服务)标识码(OSID):
ArtificialMeasurestoPromoteNaturalRegenerationinDifferentHabitatTypesofLarixprincipisrupprechtiiForest
WANGZheng-an,LIYing,YUZhi-jiaetal
(GuyuanBranch,NingxiaAcademyofAgriculturalandForestrySciences,Guyuan,Ningxia756000)
Abstract[Objective]TostudytheeffectsofartificialpromotionofnaturalregenerationmeasuresindifferenthabitattypesofNorthChinalarchplantations,developadvancedandfeasibleartificialpromotionofnaturalregenerationtechnologies,andprovidetechnicalsupportforimprovingthenaturalregenerationcapacityandstandqualityofLarixprincipis-rupprechtiihplantations.[Method]InGuyuanCity,Ningxia,fiveforestfarmswereselectedasexperimentalpointsalongthemainveinofLiupanMountainindifferentdirections.Atotalof20artificialandnaturalregenerationplotsof12m×12msizewereestablishedfordifferenthabitattypes(ninewithintheforest,sixintheforestgap,andfiveattheforestedge).Eachsamplewasmanuallysownseedsintheautumnof2021(October)andspringof2022(May);sixsurfacetreatmentmeasuresweredeployedateachexperimentalsite(A-control,B-trenching,C-holedigging,D-randomsmallholedensitypoints,E-roughplowing,andF-finelandpreparation),witheachtreatmentrepeatedthreetimes.Basedonmonitoringdatafromtwoconsecutiveyears,analyzeanddeterminethemainfactorsaffectingnaturalrenewal,andproposemeasurestopromotenaturalrenewalthroughartificialmeans.[Result]Intermsoftheconservationdensityofeachtreatmentmeasurefortwoconsecutiveyears,underautumnsowingconditionsintheforesthabitat,theorderisF(3.1plants/m2)gt;E(2.8plants/m2)gt;B(2.1plants/m2)gt;C(1.8plants/m2)gt;D(1.1plants/m2)gt;CK(0.8plants/m2).Underspringsowingconditions,theorderisF(6.4plants/m2)gt;E(5.8plants/m2)gt;B(5.1plants/m2)gt;C(3.0plants/m2)gt;D(2.0plants/m2)gt;A(1.8plants/m2).Underautumnsowingconditionsintheforestgaphabitat,theorderisE(12.6plants/m2)gt;CK(8.7plants/m2)gt;B(8.1plants/m2)gt;C(8.0plants/m2)gt;F(7.7plants/m2)gt;D(6.3plants/m2);inspringsowing,theorderisE(13.9plants/m2)gt;F(12.4plants/m2)gt;B(8.9plants/m2)gt;CK(8.2plants/m2)gt;D(7.0plants/m2)gt;C(6.5plants/m2).Undertheautumnsowingconditionsintheforestedgehabitat,theorderisC(1.9plants/m2)gt;B(1.8plants/m2)gt;E(1.4plants/m2)gt;F(1.1plants/m2)gt;CK(0.6plants/m2)gt;D(0.3plants/m2).Inspringsowing,theorderisCK(1.6plants/m2)gt;B(1.4plants/m2)gt;E(1.0plants/m2)gt;F(1.0plants/m2)gt;C,D(0.5plants/m2).[Conclusion]Overall,amongthethreetypesofhabitats,thehighestseedlingpreservationdensityisintheforestgap,followedbywithintheforestandattheforestedge.Inthesowingseason,theemergenceandpreservationdensityofthethreetypesofhabitatsarebetterinspringsowingthaninautumnsowing.Insurfacetreatmentmeasures,whetheritisautumnorspringsowing,thepreservationdensityoftheforest,habitatisbetterwithEandF.EandBarebetterwithforestgaphabitat,andF,EandBarebetterwithforestedgehabitat.
KeywordsLarixprincipis-rupprechtii;Artificialpromotionofnaturalrenewalmeasures;LiupanMountain
森林天然更新,从狭义上讲是指植物体的部分有机体丢失或损伤的自然再生长过程,从广义上讲是生物的生态过程,如植物的开花和结实,存活和生长,物种繁殖过程及其伴生种的种群变化等[1]。森林的天然更新影响着森林的结构和功能的动态变化,是森林资源再生产的重要生态过程。从种子萌发到最终枯倒,各生长阶段都受到不同的生物和非生物因素影响,这些因素如何影响天然更新是目前森林生态学研究的热点之一。影响森林天然更新的因子很多,主要包括种子库动态[2]、落物因子[3]、土壤因子[4]、林分结构[5]、林内光热条件和立地因子等[6]。目前,国内外学者对森林更新的研究大多集中于树种类型、干扰类型、森林更新影响因素、林窗特征与更新、林窗动态、森林天然更新的方式以及天然林更新过程研究等方面[7]。但是由于林分结构与研究区域的差异所得结果并不一致,因此分区域、分生境、分树种研究森林天然更新很有必要。20世纪80年代以来,部分国外学者开始对北美森林进行了一系列天然更新研究,引起了学术界关注。如Bonfil[8]研究认为,不同树种的种子大小与品质差异对更新的影响不同。Huth等[9]研究表明,林窗大小对林内小环境影响很大,进而影响幼苗更新和存活。Oliver等[10]对栎林的研究认为,弱度火干扰能促进萌芽体更新,而风倒和采伐干扰对前期幼苗、幼树的更新生长有加速作用。Myers等[11]研究认为,间伐产生的林隙干扰对森林更新有重要影响。Scariot等[12-13]研究发现,枯枝落叶层通过阻碍种子进入土壤以及减少光照而对森林更新限制较大。Muscolo等[14]研究表明,土壤理化性质(C、N、P含量)对乔木更新有重要影响。Howe等[15]研究认为,草本盖度对更新苗木的存活和生长有显著影响。Kirby等[16-17]研究认为,光照通常是林下更新的主要限制因素。总之,以上研究多是分析天然更新的影响因素,关于如何促进更新的研究相对较少。
我国森林天然更新研究起步较晚,研究范围涉及东北的阔叶红松林和暗针叶林、亚热带常绿阔叶林、热带山地雨林等森林类型[18],取得了林隙动态更新规律研究成果,如罗梅等[19]对吉林省金沟岭落叶松人工林天然更新动态研究表明,聚集分布格局有利于形成幼苗幼树生长的小环境,并利于增强群落竞争力和维持种群稳定。盖力岩等[20]对河北省木兰围场华北落叶松人工林林隙的天然更新影响研究认为,随林隙面积增大,华北落叶松和山杨的大径级和大高度级更新幼苗数量基本呈增加趋势,且林内林隙缺乏和人为活动干扰抑制了更新;朱教君等[21]对辽东地区长白落叶松人工林研究表明,林内光热环境及地被物是影响天然更新的主要因子和间伐更新树种的丰富度无明显增加,但密度和频度显著增加。张树梓等[22-23]对塞罕坝地区华北落叶松人工林研究表明,土壤-枯落物因子和土壤化学性质对幼苗更新生长的影响随龄级而变。李俊清等[24]对兴安华北落叶松幼苗生长和存活的研究表明,光照是影响幼苗更新的主要环境因素。目前大多研究集中在基于野外调查来确定天然更新的影响因素方面,但还需开展控制研究,从而更加可靠地确定主要影响因素及其定量关系,特别是提出先进可行的人工促进天然更新技术,从而有效促进天然更新,实现可持续经营。
华北落叶松属强阳性树种,具有极耐寒、旱等特点,对土壤适应能力强,是宁夏六盘山区最主要的人工造林树种之一,其人工林营造始于二十世纪六七十年代,占人工针叶林面积的89%,在保持水土、改善环境、调节气候及固碳释氧等方面发挥着重要作用。但由于六盘山地处内陆,受大陆季风气候及地形的影响,降水少而不均,且造林初期大部分为人工纯林,造成中后期成林密度过大,加之自2000年实施天然林保护工程以来长期执行的禁伐政策影响下,未进行及时合理的间伐抚育,造成林分过密,枯落物层厚,土壤酸化,植被稀少等问题,已成为制约六盘山区华北落叶松林健康稳定和可持续经营的关键问题。近年来,在六盘山区开展的有关华北落叶松林的研究主要集中在森林水文方面,而天然更新研究很少,人工促进天然更新鲜见报道。因此,笔者在华北落叶松林布设不同生境类型及人工促进天然更新的有效措施,并用出苗密度和保存密度的均值定量评价措施的有效性,以期为实现华北落叶松人工林的高质量经营管理提供科学依据。
1材料与方法
1.1研究地点和样地概况
研究区域为宁夏南部六盘山区,地处我国半湿润区向半干旱区的过渡带,黄土高原的中西部,是西北地区重要的生态安全屏障,地理位置为106°09′~106°30′E,35°15′~35°41′N,海拔为1700~2942m,地势呈东南—西北走向的狭长山脉,具有大陆性季风气候特点,春季低温少雨,夏季短暂多雹,秋季阴涝霜早,冬季严寒绵长。六盘山区光热资源较少,年日照时数2100~2400h,年均气温5.8℃,年降水量450~800mm,是泾河、清水河、葫芦河的发源地。六盘山区的植被类型既有水平地带性的森林、草原,又有由低山草甸草原、阔叶混交林、针阔混交林、阔叶矮林等组成的垂直植被景观。土壤类型带有明显的山地特征,且随海拔升高和气候差异呈现较规律的垂直分布,林区以山地灰褐土为主。
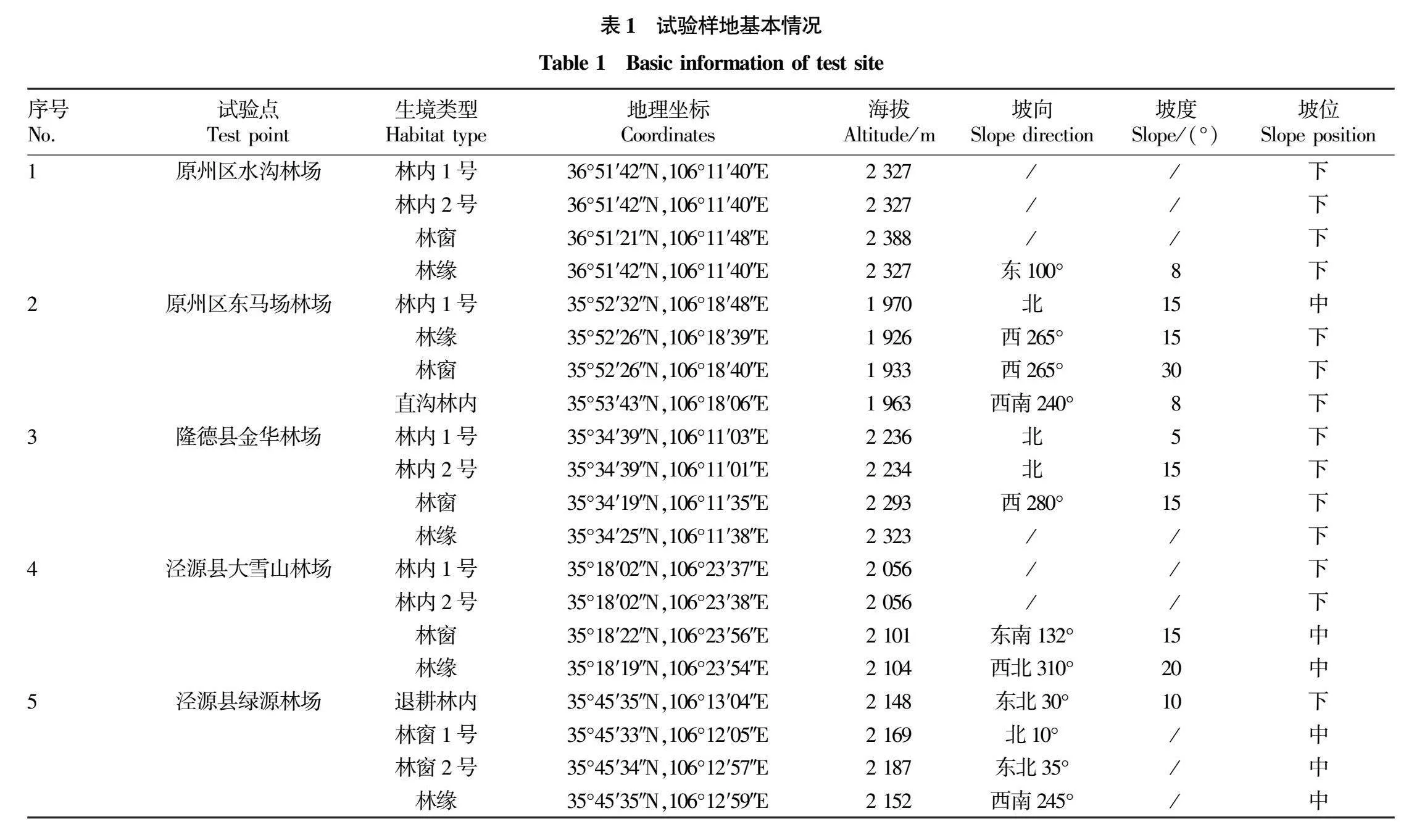
按照六盘山的南中北东西方位,选择原州区水沟林场和东马场林场、隆德县的金华林场、泾源县的大雪山林场和绿源林场5个试验点(图1),各试验点依据具体情况布设1~2块12m×12m的林内(郁闭度lt;0.8)、林窗和林缘不同生境的人工促进天然更新试验(图2),共有林内样地9个,林窗样地6个,林缘样地5个。将样地划分成4m×4m的小样方,从中随机选择1个1m×1m的观测样方。分别采取6种不同地表处理措施(A.对照;B.开沟;C.挖穴;D.随机小穴密点;E.粗翻地;F.精细整地)和2种播种方式(2021年10月秋播;2022年5月春播),每处理3次重复,随机区组排列,各试验样方基本概况见表1。
1.2试验材料
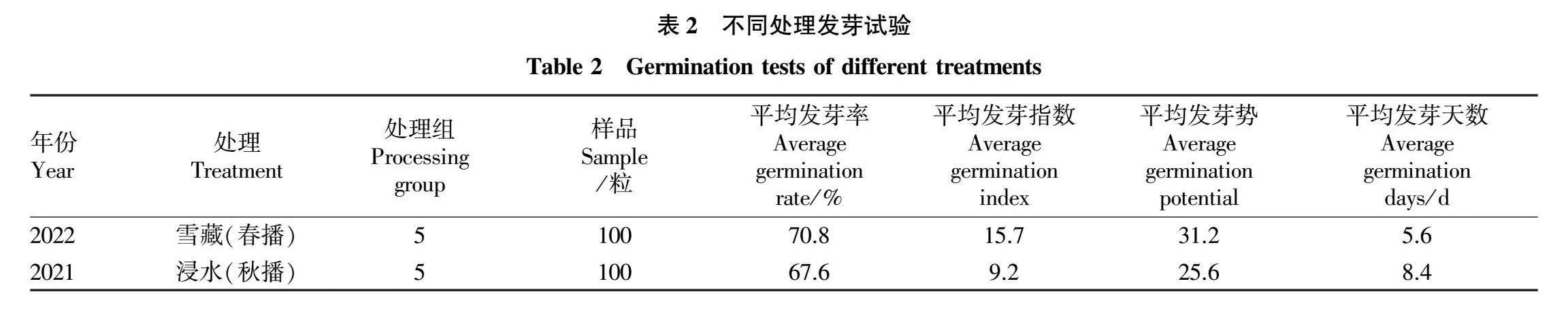
试验用种子于2021年购自六盘山华北落叶松林种源地山西宁武县管涔山,千粒重为7.5g,净度95.6%。2021年秋季播种试验是将种子在水中浸泡后播种;2022年春季播种试验是对种子雪藏处理后播种,各自的室内发芽试验结果见表2。
1.3试验方法和设计
在每个试验点的样地内均布设6种地表处理措施(图3):A对照(CK),轻微除去枯枝落叶层及杂草后,撒播种子并耙平;B开沟,每隔20cm挖1条宽20cm深10~20cm的沟播撒种子(共计6条沟);C挖大穴,每隔25cm,挖1行4个深30cm的圆筒状穴坑,播撒种子后人工覆土(1m2挖坑共4行,每行4个);D随机小穴密点,挖深5~10cm,直径5~10cm的小穴,播撒种子并耙平;E粗翻地,地表全部深翻20cm,然后播撒种子并耙平;F精细整地,地表全部清理后进行整地及清除草根等杂物,然后播撒种子并耙平。在每个试验点,设置每个地表处理重复3次,包括2m×2m样方36个,其中秋播和春播各18个样方,播撒约580g种子,每1m2样方32g(8g/m2=1200粒)。于2022年5月开始随机从4个1m2的样方选1个1m2,于2022—2023年持续监测小幼苗,每个样地36个样方(1m2),其中林内9个样地总计324个;林缘5个样地总计180个样方;林窗6个样地总计216个样方。
利用5个试验点的观测数据,分别将2022年10月和2023年10月中旬最后一次计数的幼苗量作为保存量,计算各生境幼苗保存株数的平均值,对比不同处理差异,采用SPSS21.0软件进行单因素方差分析和Duncan多重比较。
1.4数据统计
2021年10月和2022年5月播种后,对3种生境的幼苗进行连续2年监测,计数各样方中1年生和2年生的幼苗个数,利用5个试验点的观测数据,计算各生境和处理的出苗密度和保存密度均值,对比人工促进天然更新措施的效果及其生境差异。通过Excel软件和SPSS22.0对相关数据进行分析处理。
2结果与分析
2.1林内生境下地表处理措施对幼苗出苗和保存密度的影响
由表3可知,在林内生境条件下2021秋季播种各处理中,2022年最大出苗密度存在显著差异(Plt;0.05),各处理表现为F(38.6株/m2)gt;E(30.3株/m2)gt;B(19.4株/m2)gt;D(19.1株/m2)gt;C(18.0株/m2)gt;CK(14.8株/m2);2022年保存密度各处理间也存在显著差异(Plt;0.05),各处理表现为F(11.1株/m2)gt;E(7.4株/m2)gt;C(2.9株/m2)gt;CK(2.8株/m2)gt;B(2.5株/m2)gt;D(2.0株/m2);2023年保存密度在各处理间无显著差异(Pgt;0.05),各处理表现为F(3.1株/m2)gt;E(2.8株/m2)gt;B(2.1株/m2)gt;C(1.8株/m2)gt;D(1.1株/m2)gt;CK(0.8株/m2)。从各处理均值来看,与2022年最大出苗密度(23.4株/m2)相比,2022年保存密度(4.8株/m2)降低了18.6株/m2(79%),2023年保存密度(2.0株/m2)降低了21.4株/m2(91%)。
在林内生境下,从2022春季播种各处理中,2022年最大出苗密度存在显著差异(Plt;0.05),各处理表现为F(45.1株/m2)gt;E(41.1株/m2)gt;D(31.5株/m2)gt;B(31.4株/m2)gt;C(25.1株/m2)gt;CK(21.4株/m2);2022年保存密度各处理间无显著差异(Pgt;0.05),各处理大小表现为E、F(12.8株/m2)gt;B(9.1株/m2)gt;C(7.6株/m2)gt;D(5.1株/m2)gt;CK(4.4株/m2);2023年保存密度各处理间无显著差异(Pgt;0.05),各处理大小表现为F(6.4株/m2)gt;E(5.8株/m2)gt;B(5.1株/m2)gt;C(3.0株/m2)gt;D(2.0株/m2)gt;CK(1.8株/m2)。从各处理均值来看,与2022年保存密度(8.6株/m2)相比,2022年最大出苗密度(32.6株/m2)减少24.0株/m2(74%),2023年保存密度(4.0株/m2)减少28.6株/m2(88%);2023年保存密度较2022年保存密度减少4.6株/m2(53%)。
基于林内生境2年连续监测数据可知,2022年春播的最大出苗密度和2022、2023年保存密度均高于2021年秋播的对应值,分别为1.4、1.8、2.0倍。此外,在林内生境下6种地表处理措施的出苗密度和保存密度均以F和E处理较好,CK最差。这是由于林冠遮阴使林内幼苗得到的光照、水分等资源有限,而精细整地和粗翻清除了枯落物及杂草,利于幼苗获得较多的光照和水分;而CK未清除枯落物和杂草,幼苗遭受光照、水分等竞争激烈,尤其是深厚枯落物层限制种子接触和幼苗根系长入矿质土壤层,极易在干旱胁迫下死亡。
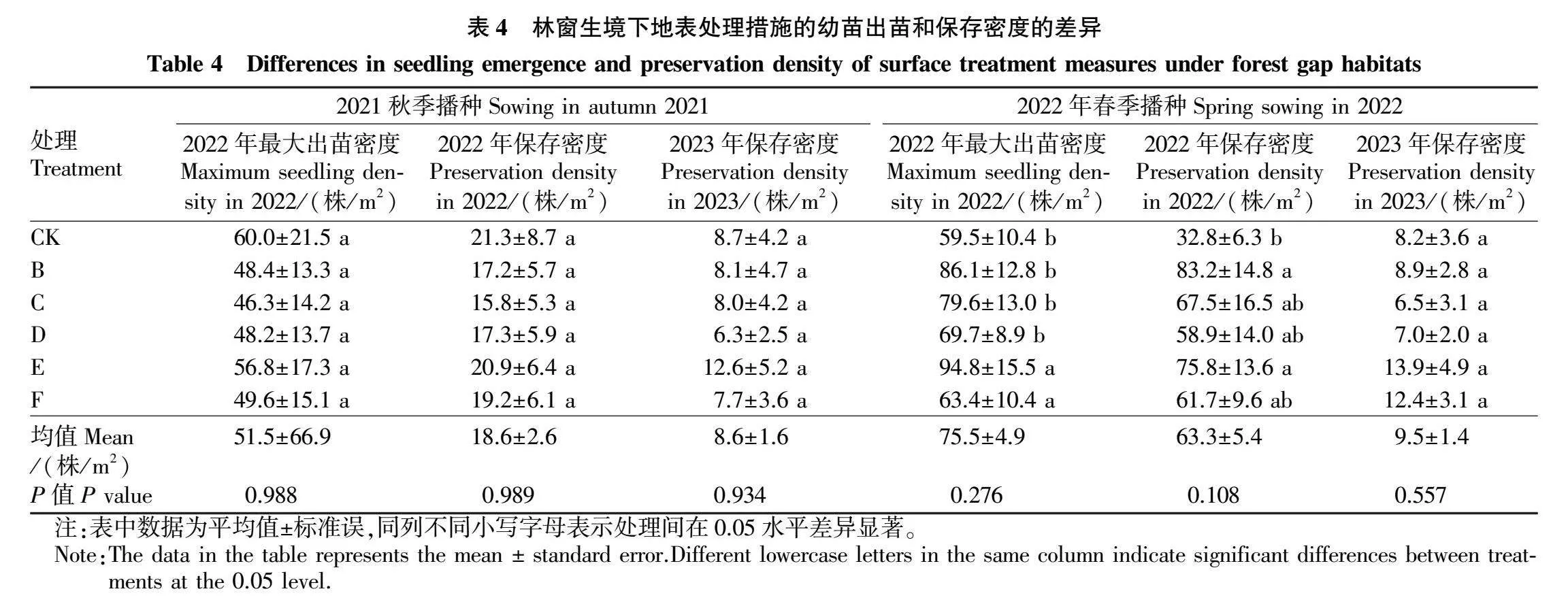
2.2林窗生境下地表处理措施对幼苗出苗和保存密度的影响
由表4可知,林窗生境下2021秋季播种各处理2022年最大出苗密度间无显著差异(Pgt;0.05),各处理表现为CK(60.0株/m2)gt;E(56.8株/m2)gt;F(49.6株/m2)gt;B(48.4株/m2)gt;D(48.2株/m2)gt;和C(46.3株/m2);2022年保存密度各处理间无显著差异(Pgt;0.05),其表现为CK(21.3株/m2)gt;E(20.9株/m2)gt;F(19.2株/m2)gt;D(17.3株/m2)gt;B(17.2株/m2)gt;C(15.8株/m2);2023年保存密度各处理间也无显著差异(Pgt;0.05),各处理表现为E(12.6株/m2)gt;CK(8.7株/m2)gt;B(8.1株/m2)gt;C(8.0株/m2)gt;F(7.7株/m2)gt;D(6.3株/m2)。从各处理均值来看,与2022年最大出苗密度(51.5株/m2)相比,2022年保存密度(18.6株/m2)降低了32.9株/m2(64%),2023年保存密度(8.6株/m2),降低了42.9株/m2(83%)。
林窗生境下2022春季播种各处理2022年最大出苗密度间无显著差异(Pgt;0.05),各处理表现为E(94.8株/m2)gt;B(86.1株/m2)gt;C(79.6株/m2)gt;D(69.7株/m2)gt;F(63.4株/m2)gt;CK(59.5株/m2);2022年保存密度各处理间无显著差异(Pgt;0.05),各处理表现为B(83.2株/m2)gt;E(75.8株/m2)gt;C(67.5株/m2)gt;F(61.7株/m2)gt;D(58.9株/m2)gt;CK(32.8株/m2);2023年保
存密度各处理间无显著差异(Pgt;0.05),各处理表现为E(13.9株/m2)gt;F(12.4株/m2)gt;B(8.9株/m2)gt;CK(8.2株/m2)gt;D(7.0株/m2)gt;C(6.5株/m2)。从各处理均值看,与2022年最大出苗密度(75.5株/m2)相比,2022年保存密度(63.3株/m2)降低了12.2株/m2(16%),2023年保存密度(9.5株/m2)降低了66.0株/m2(87%)。
由林窗生境下2年连续监测数据可知,2022年春播的最大出苗密度和其2022、2023年保存密度均高于2021秋播的对应值,分别为1.5倍、3.4倍、1.1倍。此外,林窗生境下6种地表处理措施的出苗和保存密度均以E和F较高。
2.3林缘生境下地表处理措施对幼苗出苗和保存密度的影响
由表5可知,林缘生境下2021秋季播种各处理的2022年最大出苗密度无显著差异(Pgt;0.05),各处理表现为C(11.7株/m2)gt;B(9.1株/m2)gt;D、E(8.8株/m2)gt;CK(6.7株/m2)gt;F(6.4株/m2);2022年保存密度各处理间也无显著差异(Pgt;0.05),各处理表现为E(5.8株/m2)gt;B(4.9株/m2)gt;F(4.5株/m2)gt;C(4.3株/m2)gt;D(2.8株/m2)gt;CK(2.6株/m2);2023年保存密度在各处理间也无显著差异(Pgt;0.05),各处理表现为C(1.9株/m2)gt;B(1.8株/m2)gt;E(1.4株/m2)gt;F(1.1株/m2)gt;CK(0.6株/m2)gt;D(0.3株/m2)。从各处理均值来看,与2022年最大出苗密度(8.6株/m2)相比,2022年保存密度(4.1株/m2)降低了4.5株/m2(52%),2023年保存密度(1.2株/m2)降低了7.4株/m2(86%)。
林缘生境下2022春季播种各处理2022年最大出苗密度间无显著差异(Pgt;0.05),各处理表现为B(28.5株/m2)gt;C(28.2株/m2)gt;CK(22.3株/m2)gt;E(19.8株/m2)gt;F、D(18.3株/m2);2022年保存密度各处理间也无显著差异(Pgt;0.05),各处理表现为F(21.8株/m2)gt;E(20.7株/m2)gt;CK(15.1株/m2)gt;B(13.6株/m2)gt;C(10.7株/m2)gt;D(9.2株/m2);2023年保存密度各处理间无显著差异(Pgt;0.05),各处理表现为CK(1.6株/m2)gt;B(1.4株/m2)gt;E(1.0株/m2)gt;F(0.9株/m2)gt;C、D(0.5株/m2)。从各处理均值看,与2022年最大出苗密度(22.6株/m2)相比,2022年保存密度(15.2株/m2)降低了7.4株/m2(33%),2023年保存密度(1.0株/m2)降低了21.6株/m2(96%)。
由林缘生境2年连续监测数据可知,2022春季播种的最大出苗密度和其2022年保存密度均高于2021年秋季播种的对应值,分别为2.6倍、3.7倍。此外,在林缘生境下6种地表处理措施中,从其保存密度看,秋季播种以E和B较高;春季播种以E和F处理较高。
3讨论与结论
该研究在六盘山区不同空间位置5个试验点华北落叶松人工林,选择林内、林窗和林缘3种生境,在经过6种地表处理措施的基础上进行了秋播和春播,然后调查和对比出苗密度和保存密度的差异,以便寻找能够有效人工促进华北落叶松天然更新的措施。在林内生境各地表措施处理中,无论秋播还是春播,均以精细整地、粗翻处理的出苗和保存密度较高,对照最差,这与樟子松人工林天然更新的研究结果相似[25],研究表明人为干预下的出苗率更高。这是由于幼苗生长需要植根与土壤,地表处理清除了枯落物层的阻碍,使幼苗根系更易接触到土壤,进而从土壤中获取足够的水分和营养[26];另一方面,清理地表的枯落物和草灌后使幼苗能受到更多阳光照射,进而改善地表的水热等微环境[27]。在林窗生境下,地表处理措施同样具有促进出苗密度及提高幼苗保存密度的作用,其中以粗翻和精细整地的效果好,CK的效果最差。这是由于林窗内灌草生长茂盛,在水分、光照、养分等方面与幼苗有较强的竞争,会导致种子萌发和幼苗定植困难,甚至引发幼苗大量死亡[28]。也有研究表明,去除草本根系利于增加华北落叶松幼苗根系生长的空间和养分竞争力[29]。在林缘生境下,当年出苗密度以对照较好,这是由于六盘山地区在种子出苗阶段(5—6月)的降水较少,强烈日光照射导致地温升高和表层土壤湿度降低,但对照样方的草本层遮阴保存了较高的表层土壤湿度,为种子发芽提供了较好条件,因此出苗密度较好;然而后期的幼苗保存密度较低,主要是由于降水增加导致草本快速生长,与幼苗竞争光照、水分、养分,导致死亡率增加。
在3种生境的不同地表处理措施中,无论是当年最大出苗密度还是连续2年监测的幼苗保存密度,均为春播高于秋播。这主要由于春播对种子进行雪藏,一是利于预防猝倒病发生[30];二是对种子起到了催芽作用,可以有效提高出苗率;而秋播种子处理在从冬初到第2年发芽前的一段时间内易遭动物(鼠类、鸟类等)啄食种子[31]。例如在黄土高原子午岭地区的调查表明,从油松种子降落到翌年4月,5种林分的土壤种子库总量下降了42.34%~53.59%,动物的搬运、取食和种子腐烂死亡是种子库损耗的主要原因[32]。
综合3类生境来看,秋季播种情况下的2022年保存密度表现为林窗(18.6株/m2)gt;林内(4.8株/m2)gt;林缘(4.1株/m2),2023年保存密度表现为林窗(8.6株/m2)gt;林内(2.0株/m2)gt;林缘(1.2株/m2);春季播种情况下的2022年保存密度表现为林窗(63.3株/m2)gt;林缘(15.2株/m2)gt;林内(8.6株/m2),2023年保存密度表现为林窗(9.5株/m2)gt;林内(4.0株/m2)gt;林缘(1.0株/m2),综合来看,林窗内幼苗保存密度最高,林缘和林内次之。这是由于林窗中较高的空气温度、较强的光照条件和适宜的土壤湿度能够满足幼苗喜光耐旱的生长特性[33];而林内遮阴的环境和较厚的枯落物层,使大量幼苗在萌发生根过程中难以穿透持水能力较低的枯落物层进入水分供应相对稳定的矿质土层,极易受到干旱胁迫[34],或因其在生长阶段未获得足够光照而导致保存率较低。因此,林窗生境对于减少幼苗萌发和生长过程中的死亡率有积极作用。这与李进等[35]对关帝山同树种的更新苗生长的研究结论基本一致。总之,幼苗在野外发芽及保存受到多种因素交叉影响,如环境因子(地形、土壤、水分、灌草竞争、光照等)与林地微环境,其影响过程和机理复杂,还需进一步加强综合研究,从而深入认识、科学调控、充分利用华北落叶松的天然更新。
参考文献
[1]李小双,彭明春,党承林.植物自然更新研究进展[J].生态学杂志,2007,26(12):2081-2088.
[2]CARLTONGC,BAZZAZFA.Regenerationofthreesympatricbirchspeciesonexperimentalhurricaneblowdownmicrosites[J].EcolMonogr,1998,68(1):99-120.
[3]姚佳峰,郭钰,董媛,等.枯落物厚度对华北落叶松人工林天然更新的影响[J].应用生态学报,2024,35(4):1025-1032.
[4]ESSERYR.Borealforestsandsnowinclimatemodels[J].HydrolProcess,1998,12(1011):1561-1567.
[5]曾德慧,尤文忠,范志平,等.樟子松人工固沙林天然更新障碍因子分析[J].应用生态学报,2002,13(3):257-261.
[6]杨莎.关帝山天然次生针叶林林下更新苗空间格局研究[D].太谷:山西农业大学,2014.
[7]唐建忠.影响天然林树种更新的因素探讨[J].黑龙江科技信息,2014(35):272.
[8]BONFILC.Theeffectsofseedsize,cotyledonreserves,andherbivoryonseedlingsurvivalandgrowthinQuercusrugosaandQ.laurina(Fagaceae)[J].AmericanJEcol,1998,85(1):79-87.
[9]HUTHF,WAGNERS.GapstructureandestablishmentofSilverbirchregeneration(BetulapendulaRoth.)inNorwaysprucestands(PiceaabiesL.Karst.)[J].ForEcolManag,2006,229(1/2/3):314-324.
[10]OLIVERCD,LARSONBC.Foreststanddynamics[M].NewYork:JohnWileyandSons,1996:520.
[11]MYERSGP,NEWTONAC,MELGAREJOO.TheinfluenceofcanopygapsizeonnaturalregenerationofBrazilnut(Bertholletiaexcelsa)inBolivia[J].ForEcolManag,2000,127(1/2/3):119-128.
[12]SCARIOTA.SeedlingmortalitybylitterfallinAmazonianforestfragments[J].Biotropica,2000,32(4):662-669.
[13]BREWERSW,WEBBMAH.Ignorantseedpredatorsandfactorsaffectingtheseedsurvivalofatropicalpalm[J].Oikos,20093(1):32-41.
[14]MUSCOLOA,SIDARIM,MERCURIOR.Inliuenceofgapsizeonorganicmatterdecomposition,microbialbiomassandnutrientcycleinCalabrianpine(Pinuslaricio,Poiret)stands[J].ForEcolManag,2007,242(2/3):412-418.
[15]HOWEHF,SMALLWOODJ.Ecologyofseeddispersal[J].AnnuRevEcolSyst,1982,13:201-228.
[16]KIRBYKJ.Changesinthegroundfloraunderplantationsonancientwoodlandsites[J].Forestry,1988,61(4):317-338.
[17]KOBERK,PACALASW,SILANDERJAJR,etal.Juveniletreesurvivorshipasacomponentofshadetolerance[J].EcolAppl,1995,5(2):517-532.
[18]杨娟,葛剑平,刘丽娟,等.卧龙自然保护区针阔混交林林隙更新规律[J].植物生态学报,2007,31(3):425-430.
[19]罗梅,郑小贤.金沟岭林场落叶松人工林天然更新动态研究[J].中南林业科技大学学报,2016,36(9):7-11.
[20]盖力岩,于树峰,徐学华,等.木兰围场华北落叶松人工林林隙对天然更新的影响[J].河北林果研究,2014,29(3):253-257.
[21]朱教君,刘足根,王贺新.辽东山区长白落叶松人工林天然更新障碍分析[J].应用生态学报,2008,19(4):695-703.
[22]张树梓,李梅,张树彬,等.塞罕坝华北落叶松人工林天然更新影响因子[J].生态学报,2015,35(16):5403-5411.
[23]孙国龙,李文博,黄选瑞,等.华北落叶松人工林天然更新及与土壤因子的关系[J].安徽农业大学学报,2017,44(6):1047-1051.
[24]李俊清,李景文.中国东北小兴安岭阔叶红松林更新及其恢复研究[J].生态学报,20023(7):1268-1277.
[25]马存平,石长春,贾艳梅,等.不同措施处理下樟子松人工林天然更新比较[J].防护林科技,2023(4):4-6.
[26]杨秀清.影响关帝山华北落叶松天然更新与幼苗存活的微生境变量分析[J].山西农业大学学报(自然科学版),2010,30(6):542-547.
[27]李龙杰,王杰,任云卯,等.人工促进更新措施对落叶松种子萌发和早期生长的影响[J].北京林业大学学报,20245(4):24-35.
[28]CONNELLJH.Ontheroleofnaturalenemiesinpreventingcompetitiveexclusioninsomemarineanimalsandinforesttrees[J].DynPopul,1970,298:298-312.
[29]张静静,刘宁,南宏伟,等.草本地下竞争对3种幼苗存活和生长的影响[J].中南林业科技大学学报,2019,39(1):71-76.
[30]王月玲,张源润,季波,等.宁夏南部土石质山区华北落叶松规范化造林技术[J].宁夏农林科技,2008,49(3):84-85,74.
[31]吴敏,张文辉,周建云,等.秦岭北坡不同生境栓皮栎种子雨和土壤种子库动态[J].应用生态学报,20122(11):2807-2814.
[32]张希彪,王瑞娟,上官周平.黄土高原子午岭油松林的种子雨和土壤种子库动态[J].生态学报,2009,29(4):1877-1884.
[33]许宁,憨宏艳,甘小洪.光照及地面覆盖物对水青树种子萌发和幼苗初期生长的影响[J].植物资源与环境学报,2015,24(3):85-93.
[34]陈龙涛.华北落叶松和油松幼苗对干旱胁迫的响应及其抗旱性评价[D].太谷:山西农业大学,2017.
[35]李进,石晓东,高润梅,等.华北落叶松天然次生林更新及影响因素[J].森林与环境学报,2020,40(6):588-596.
