我国几种油茶植物染色体研究进展
2025-01-24韦开萍蒙进芳郭晓春






摘要我国油茶植物新品种少,良种化程度低,培育高产、优质、高抗新品种迫在眉睫。通过整理普通油茶(CamelliaoleiferaAbel)、小果油茶(Camelliameiocarpa)、越南油茶(Camelliavietnamensis)、高州油茶(Camelliagauchowensis)、攸县油茶(Camelliayuhsienensis)、五柱滇山茶(Camelliayunnanensis)、滇山茶(Camelliareticulat)、怒江红山茶(Camelliasaluenensis)、西南红山茶(Camelliapitardii)、浙江红山茶(Camelliachekiangoleasa)、长尾红山茶(Camellialongicaudata.)11种中国现有作为油料资源人工栽培利用或有较大利用前景的油茶物种染色体核型分析资料,从染色体倍性与核型方面叙述几种油茶物种染色体核型研究情况,为油茶杂交育种等工作提供理论指导。
关键词油茶;染色体;核型
中图分类号S794.4"文献标识码A"文章编号0517-6611(2025)02-0007-06
doi:10.3969/j.issn.0517-6611.2025.02.002
开放科学(资源服务)标识码(OSID):
ComprehensiveAnalysisoftheChromosomeStudiesonCamelliaoleiferaTraditionallyUsedasOilPlantinChina
WEIKai-pingMENGJin-fangGUOxiao-chun2
(1.SouthwestForestryUniversity,Kunming,Yunnan650224;2.InstituteofOilTeaResearch,YunnanAcademyofForestryandGrassland,Guangnan,Yunnan663306)
AbstractCamelliaoleiferaistraditionallyusedasoilplantinChina.Currently,theplantingvarietiesofCamelliaoleiferawerelimitedandlowdegreeofimprovedbreedinginChina,indicatedtheurgentneedforthecultivationofhigh-yield,superiorquality,andresilientnewvarieties.Thisstudywasacomprehensivereviewontheprogressioninchromosomalkaryotypesof11traditionallyoilplantspeciesofCamelliaoleifera,includingCamelliaoleiferaAbel,Camelliameiocarpa,Camelliavietnamensis,Camelliagauchowensis,Camelliayuhsienensis,Camelliayunnanensis,Camelliareticulata,Camelliasaluenensis,Camelliapitardii,CamelliachekiangoleasaandCamellialongicaudata.ThesespeciesserveasvitalsourcesofoilinChina,showcasingimmenseprospectivebenefitsforcultivationandutilization.Thisstudyofferedanin-depthunderstandingofthesespeciesfromchromosomalperspective,consideringtheirploidyandkaryotypes.Byavailingthisessentialtheoreticalguidance,itaimedtosupportandenhancetheongoingworkinCamelliaoleiferahybridbreedingandrelatedendeavors.
KeywordsCamelliaoleifera;Chromosome;Karyotype
中国的油茶按照油用资源分类共有15组3大类200余种[1]。栽培面积最大的为普通油茶,其次为小果油茶、越南油茶等,其余油茶种类大多数处于半野生、野生状态[2]。油茶植物油营养丰富,是理想的食用油,其副产品综合利用价值高,广泛应用于工业、农业、医药等方面;而有些油茶物种还具有花大花美、类型多样等特点,观赏价值极高[3-4]。近年来,油茶产业成为林业行业践行习近平总书记“绿水青山就是金山银山”和“生态产业化、产业生态化”生态文明思想的载体,有着极其重要的战略地位。目前油茶产业面对的关键问题是新品种少,良种化程度低,迫切需要培育高产、优质、高抗新品种[5]。
对物种进行染色体核型分析是物种进化分析、研究和培育新品种的重要手段,核型参数是了解和研究植物的重要基础数据,是植物分类、鉴定的重要指标,也是指导杂交育种的重要基础。我国主要油茶物种的染色体观察研究工作开始于1980年,之后有关油茶植物的染色体数目及核型研究成果陆续报道[6]。但是各种油茶植物染色体核型分析资料较为分散、未进行系统整理研究。因此,笔者整理现有中国作为油料资源人工栽培利用或有较大利用前景的油茶物种染色体核型分析资料(主要为白花和红花油茶类植物),从染色体倍性与核型方面叙述几种油茶物种染色体核型研究概况,并对整理的资料进行简单数据分析,为油茶杂交育种等工作提供理论指导。
由于山茶属物种的系统发育和分类,目前学界还没有形成统一的认识,该研究叙述的分类采用张宏达分类系统[7]。在收集整理过程中发现,有些资料仅有染色体长臂、短臂长度,染色体相对长度等基础数据,所以在收集过程中对此类数据进行再次计算和分析,其中核型分析方法参照文献[8],染色体长度比(L/S)=最长染色体/最短染色体,平均臂比值(AR)=总臂比值/染色体数目,核型不对称系数(As.K.)=(长臂总长/全组染色体总长)×100%。
1白花油茶类植物
1.1普通油茶
普通油茶是我国栽培面积最广、总产油量最高的木本油料植物[9]。在长期的栽培选育过程中,现有普通油茶品种达300多种。国内现有普通油茶染色体核型研究资料见表1。
1981年,黎麦秋[10]首次对普通油茶进行染色体核型研究,结果表明普通油茶染色体数目为2n=30,染色体基数为x=15,为二倍体。而后,三倍体、四倍体、六倍体普通油茶逐渐被发现[11-13]。其中,二倍体普通油茶染色体核型明确,三倍体、四倍体核型资料未见报道,六倍体普通油茶核型研究资料众多。由表1可知,六倍体普通油茶核型多样,核型变化明显。
2020年,叶天文等[14]研究发现,“华硕”“华鑫”“华金”3个品种的实生苗染色体数目为90、87、85和75,出现五倍体和非整倍体,油茶实生后代出现染色体缺失现象。3个品种扦插苗染色体数均为2n=90,为六倍体,不同品种间核型不一致,随体数量和位置都不相同,核型组成存在差异,但核型不对称系数相近,其进化程度基本一致。2022年,李艳民等[15]对油茶种间杂交F1代18个优良单株进行核型分析结果表明,母本“攸杂2”为非整倍体,父本“华硕”为整倍体,杂交子代的核型参数介于父本与母本之间,少数杂交子代的核型出现较大变异。
1.2小果油茶
小果油茶(C.meiocarpa)是我国特有的油料作物,皮薄,出籽率高,栽培面积和年产量仅次于普通油茶,是优良油茶树种[16]。
资料显示,小果油茶的染色体数目为2n=60,为四倍体,核型资料至今未见报道[13,17]。对于普通油茶与小果油茶是否为一个种的问题,至今仍有争论,如基于微卫星分子标记的分析结果,小果油茶与普通油茶极有可能为同一物种,但是基于nrITS的结果,则认为小果油茶与普通油茶不是同一物种[18]。而根据两者的细胞倍性,有学者推测四倍体的小果油茶可能是普通油茶的祖先之一,通过与野外二倍体普通油茶杂交后经过多倍化形成异源六倍体的普通油茶[17]。总之,两者之间的关系扑朔迷离,其细胞学水平上的研究还有待深入。
1.3高州油茶
高州油茶(C.gauchowensis)是油茶中较为优良的物种,果大叶茂,单位面积产量、出籽率、出油率都较高,栽培面积位于全国油茶栽培面积第3[19]。
关于高州油茶染色体核型的研究较少,目前仅发现顾建志等[20]报道,高州油茶染色体数为2n=30,为二倍体,L/S 1.46,AR 1.72,核型公式为2n=30=17m+12sm+1st,无随体,核类型为2A。相对于其他种类的油茶植物来说,高州油茶的染色体研究极少,遗传背景不够清晰。而作为中国油茶栽培面积第3位的油茶树种,高州油茶在中国栽培油茶植物中占有重要地位,其在染色体方面的研究有待深入。
1.4越南油茶
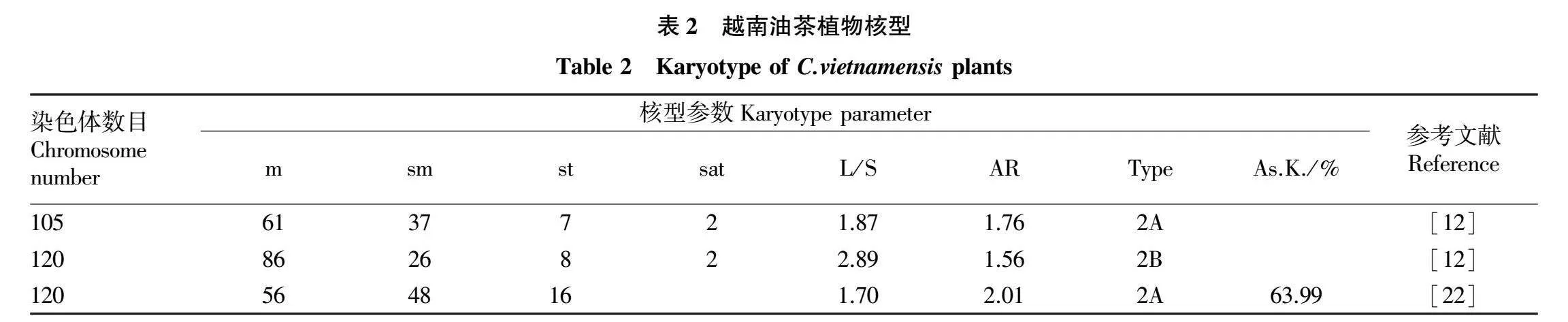
相较于普通油茶、小果油茶、高州油茶,越南油茶(C.vietnamensis)栽培面积较小、产量最低、但却最具有地域特色[21]。其染色体核型研究资料见表2。
从现有的研究结果看,对越南油茶染色体研究不多,都是早期进行的。结果表明,越南油茶染色体数目为2n=105、120,既有七倍体,也有八倍体,主要由m、sm、st 3种核型组成,七倍体核型明确,但八倍体的两个核型研究结果却有显著差异[12,22]。但由于早期染色体研究技术不成熟,造成越南油茶八倍体核型显著差异的原因还有待研究。
1.5攸县油茶
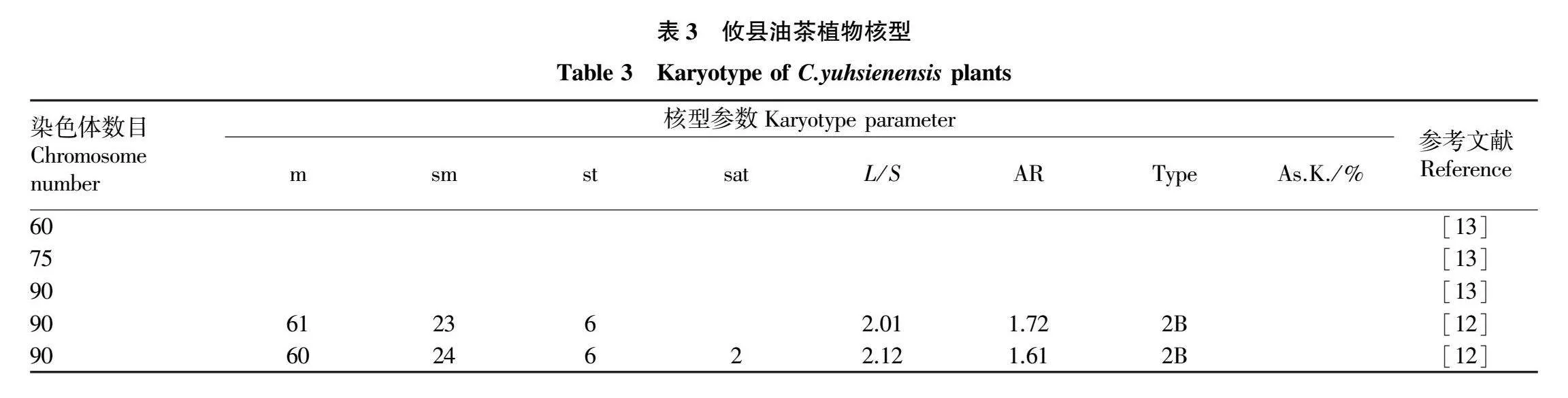
攸县油茶(C.yuhsienensis)属短柱茶组,具有树形紧凑、果壳薄、出籽率高、抗病性强的特点,具有良好的应用前景[23]。其染色体核型研究资料见表3。
关于攸县油茶的染色体研究结果表明,攸县油茶染色体数有2n=60、75、90,有四倍体、五倍体、六倍体,种内多倍性明显[13]。在核型研究中,四倍体及五倍体攸县油茶核型未见相关报道;关于六倍体核型的2个研究结果几乎一致,仅相差1条m型染色体,且随体数量有差异[12]。
1.6五柱滇山茶
五柱滇山茶(C.yunnanensis)属实果茶组,植株挂果率高,优良单株可产茶籽超过100 kg,是兼具食用、药用、观赏、绿化等多重价值的优良树种[24]。其染色体核型研究资料见表4。
五柱滇山茶染色体为2n=30,为二倍体。其核型主要由m、sm、st 3种染色体组成,核型组成差异较大,核型多态性特征显著[11-12,25]。五柱滇山茶从倍性以及核型看在进化程度上属于较为原始的物种,是优良的遗传种质资源,应做好保护工作。
2红花油茶类植物
2.1滇山茶
滇山茶(C.reticulata),国家二级保护珍稀植物,云南特有的优良木本油料树种,云南山茶花的原始种,具有出油率高、花大、花类型丰富的优良特性,利用价值极高[26-27],目前学者们对滇山茶的重视程度越来越高,其细胞学方面的研究资料也越来越多,遗传背景逐渐清晰,各研究结果见表5。
在滇山茶染色体研究中,二倍体、四倍体、六倍体、八倍体、十倍体滇山茶逐渐被发现[13,28-33]。可见,在长期的人工选择培育下,滇山茶倍性多样,种内多倍性极其明显。不同学者研究的滇山茶六倍体核型差异极大,共相差10条m染色体[31]。杜宁[32]收集的研究资料显示,四倍体、六倍体滇山茶的核型均有差异,即使核型组成一样,其随体数量与位置也都有差异。2023年,李拓键等[35]采用流式细胞术结合常规压片的方法对滇山茶进行染色体核型分析表明,3种倍性的滇山茶主要染色体类型为m和sm,其中绝大多数为m,其中二倍体滇山茶的核型与杜宁[32]收集的资料结果相比较,其核型也有差异。所以,从总体的核型研究结果资料来看,其二倍体、四倍体、六倍体滇山茶染色体的主要类型为m和sm,绝大多数为m,其核型多态性特征也极其明显。其中,腾冲红花油茶为六倍体,应是六倍体滇山茶类型,核型资料未见研究报道。
染色体倍性的研究表明,多倍化是滇山茶进化的主要途径,杜宁[32,36]研究表明,二倍体滇山茶和西南山茶是四倍体滇山茶的杂交亲本,怒江红山茶、四倍体滇山茶为六倍体滇山茶杂交亲本,同时推测参与六倍体滇山茶形成的其他亲本有西南红山茶(C.pitardii)、猴子木(C.yunnanensis)。同时,顾志建[30]研究发现,滇山茶二倍体类型的分布相对局限,四倍体类型在有些地区成片分布;六倍体类型分布区相对广泛,有些地区构成一连续分布的格局。可见,滇山茶多倍体类型适应环境能力显然更强[36]。其次,从表格中现有的数据资料来看,不同倍性下的核型差异程度从小到大为二倍体→四倍体→六倍体。由此,推测滇山茶进化程度越高,核型差异越大,核型多态性特征越明显,环境适应能力越强。该途径可为其他山茶属植物的育种工作提供参考理论,通过杂交多倍化途径提高一些物种的利用率。
腾冲红花油茶植物是滇山茶中主要的油用植物类型,有丰富多样的自然类型,根据花瓣轮数,可分为单瓣、半重瓣和重瓣3个类型。在DNA水平上的差异表明,3种瓣型具有各自独特的遗传物质基础,说明划分为单瓣、半重瓣和重瓣3个自然类型具有一定的科学性和合理性[4,37]。而由于基因剂量效应,染色体多倍化后植物性状会发生相应改变,如器官巨型化、花果增大增重等[38]。并且,六倍体滇山茶杂交亲和性、育性、抗逆性以及适应性均强于二倍体、四倍体滇山茶[39]。由此可推测,3种瓣型腾冲红花油茶植物花瓣数量、花瓣大小、胚珠发育情况有显著差异的原因与其倍性有紧密联系,应做进一步的研究探索。
2.2怒江红山茶
怒江红山茶(C.saluenensis)是滇山茶的近缘种,植物根系较发达,耐旱耐贫瘠,花朵密集美丽,种子所含油脂可食用[40],是滇山茶多倍化进化的杂交亲本。其核型资料见表6。
在怒江红山茶的染色体核型研究中发现,其染色体数目为2n=30、60,既有二倍体,也有四倍体[11-12]。早期研究发现怒江红山茶核型由18条m型、8条sm型、4条st型染色体组成,核类型为2B,而后期不同研究结果中的核型组成不尽相同,与早期的结果差异较大[12,41]。孙静贤等[41]对中国特有植物怒江红山茶3个居群的染色体数目和核型进行研究发现,不同居群的染色体数目一样,但染色体存在多态性变异,3居群m、sm和st染色体数目存在变化,有m染色体向sm转化,或者sm染色体向st染色体转化的现象。
2.3西南红山茶
西南红山茶(C.pitardii)是木本油料植物,初春开花,花期长,花色从白色、粉红色、桃红色到红色,也是滇山茶多倍化进化的杂交亲本[42]。其核型资料见表7。
从现有的染色体核型研究的结果看,西南红山茶染色体为2n=30、90,即二倍体和六倍体。二倍体西南红山茶核型主要由m、sm型染色体组成,核类型均为2B,其核型有差异[11、31],但其六倍体核型未见报道。
2.4浙江红山茶
浙江红山茶(C.chekiangoleasa)是我国南方山区的特有树种,花红色艳且花期长,园林观赏价值较高[43]。其染色体核型研究资料见表8。
对浙江红山茶的染色体研究也较少,研究结果表明,其染色体数目为2n=30,为二倍体,从已有的2个核型结果看,其核型主要由m、sm 2种染色体类型组成,核型差异不大,但其随体数量及位置不一致,核型对称系数接近[13,44-45]。
2.5长尾红山茶
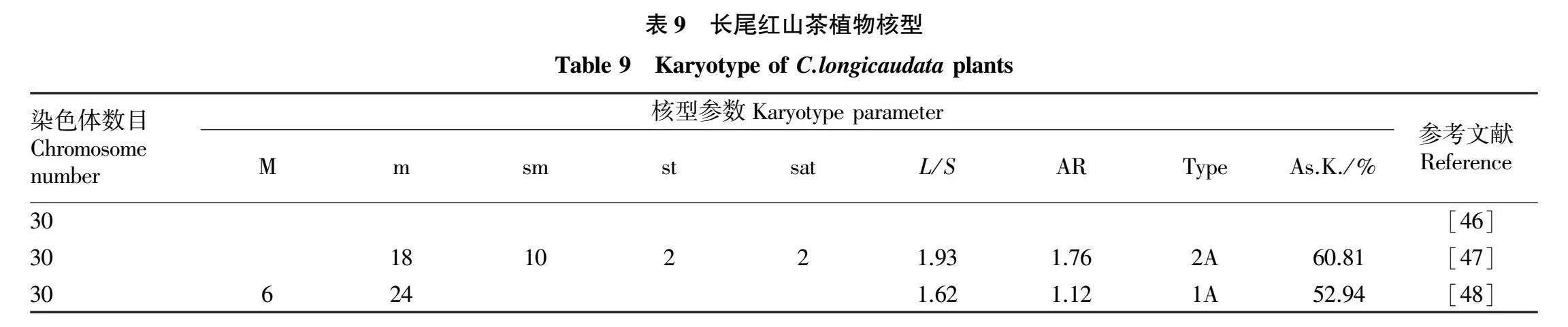
长尾红山茶(C.longicaudata)人工栽培利用程度低,目前基因还未被开发利用,是良好的育种材料。其染色体核型研究资料见表9。
长尾红山茶染色体数目为2n=30,为二倍体,核型分析结果也有较大差异;张志刚等[48]研究发现,长尾红山茶由6条M型染色体,24条m型染色体组成,核类型为1A,核型不对称系数为52.94%,属于核型较为对称类型,与陈维新等[46-47]的研究结果差异较大。且该核型与其他红花油茶类植物差异都较大,按照核型由对称向不对称方向进化来看,长尾红山茶处于较为原始的状态。
3总结及展望
已有资料表明,该研究中的油茶物种染色体数目研究已完成,但是各物种的核型研究资料仍不完善,如作为栽培面积和年产量仅次于普通油茶的小果油茶核型研究至今未见报道,其细胞学研究程度不深;滇山茶中作为主要油用类型植物的腾冲红花油茶的核型资料也不清晰,其后期研究的六倍体滇山茶核型与腾冲红花油茶核型是否有差异也未知;三倍体、四倍体的普通油茶核型,四倍体、五倍体攸县油茶,八倍体、十倍体滇山茶,四倍体怒江红山茶,六倍体西南红山茶的核型资料缺失。总之,现有中国作为油料资源人工栽培利用或有较大利用前景的油茶物种染色体研究程度还不深入,遗传背景资料还不够完善,且不同倍性的油茶植物都是以后油茶物种进行遗传改良的优良材料,由此可见,对其进行核型研究是极其必要的。同时,由五柱滇山茶、长尾红山茶现有的核型研究资料来看,其进化程度低,基因利用程度不高,是培育新品种的优良材料,应加以关注。
根据已有研究结果,在中国作为油料资源的几种白花与红花类油茶植物中部分物种具有山茶属植物种内多倍性与核型多态性的特点,即同一物种除了二倍体之外,还有多倍体类型,不同居群间存在核型变异以及同一居群不同个体间具有不同的细胞型和核型,如普通油茶、越南油茶、攸县油茶、五柱滇山茶、滇山茶、怒江红山茶、西南红山茶。而其余几种植物细胞学研究的资料较少,是否跟普通油茶、滇山茶等物种具备相应的特征,还有待深入研究。该研究的植物种类中,普通油茶与滇山茶在各自的分组中属于被人工栽培利用、研究选育最多的物种,而其种内多倍性、核型多态性特征也最明显,按照山茶属植物由二倍体向多倍体、核型对称向不对称的演化方向来看,普通油茶与滇山茶都处于较为进化的地位,由此可见,人工选择会加快物种进化速度。
综上可知,许多油茶物种试验材料来自种子发芽的根尖部位,而有些油茶植物为异花授粉植物,种子中的细胞染色体数量和核型并不能代表其植株本身,且大多数由种子发芽的根部测定的染色体数会出现非整倍体或是结果不一致的现象。因此,建议在油茶植物进行细胞学研究,尤其是对品种进行细胞学鉴定时采用植株幼嫩茎尖或者扦插苗根尖部位。前期进行染色体倍数的鉴定时,大多数学者都是根据山茶属植物染色体基数为n=15来计算其染色体倍性,结果有一定的局限性。流式细胞术(FCM)是应用流式细胞仪进行分析、分选的技术,是基于DNA含量变化的基础进行的,比常规染色体计数更加快捷方便、快速、准确[49]。由此,后期在进行油茶植物染色体倍性鉴定时,建议采用流式细胞术。虽然前人对油茶植物进行过大量的染色体研究,但由于技术的局限性、使用材料的不定性以及山茶属植物同物异名导致研究物种的不确定等问题,使以往的研究结果可能与实际情况有出入,同一物种不同学者的研究结果不尽相同,其原因除开其山茶属植物具有核型多态性的特点外,就是因为不同学者的试验技术、人工计数误差所致。因此,建议在各油茶物种细胞学基础上进行其他研究时,重新对其进行细胞学研究,以使研究数据更为严谨。
参考文献
[1]姚小华,任华东.中国油茶遗传资源[M].北京:科学出版社,2020.
[2]李利敏.八种油茶蒲提取物活性物质研究及其抗氧化活性比较[D].杭州:浙江大学,2013.
[3]吴雪辉,黄永芳,谢治芳.茶油的保健功能作用及开发前景[J].食品科技,2005,30(8):94-96.
[4]沈立新,梁洛辉,王庆华,等.腾冲红花油茶自然类型及其品种类群划分[J].林业资源管理,2009(6):75-79.
[5]陈永忠,罗健,王瑞,等.中国油茶产业发展的现状与前景[J].粮食科技与经济,2013,38(1):10-12.
[6]顾志建.山茶属植物染色体研究概述[C]//周光召.西部大开发科教先行与可持续发展:中国科协2000年学术年会文集.北京:中国科学技术出版社,2000:470-471.
[7]张宏达.中山大学学报(自然科学)论丛(1):山茶属植物的系统研究[M].广州:中山大学学报编辑部,1981.
[8]李懋学,陈瑞阳.关于植物核型分析的标准化问题[J].武汉植物学研究,1985(4):297-302.
[9]庄瑞林.中国油茶[M].2版.北京:中国林业出版社,2008.
[10]黎麦秋.普通油茶、板栗染色体组型和GiemsaC—带的带型研究[J].林业科技通讯,1981(4):9-12.
[11]李光涛,梁涛.山茶属植物的染色体数目和核型[J].广西植物,1990(2):127-137.
[12]张文驹,闵天禄.山茶属的细胞地理学研究[J].云南植物研究,1999,21(2):184-196.
[13]黄少甫,赵治芬.中国主要油茶物种染色体观察[J].亚热带林业科技,1981(4):18-24.
[14]叶天文,李艳民,张健,等.普通油茶染色体制片技术优化及核型分析[J].南京林业大学学报(自然科学版),2020,44(5):93-99.
[15]李艳民,袁德义,叶天文,等.油茶种间杂交F1代18个优良单株核型分析[J].林业科学,2022,58(4):165-174.
[16]黄勇.小果油茶遗传多样性分析及杂交渐渗研究[D].北京:中国林业科学研究院,2011.
[17]秦声远.普通油茶系统发育及种群遗传分化的地理格局[D].南昌:南昌大学,2018.
[18]姚小华,黄勇.小果油茶资源与遗传多样性研究[M].北京:科学出版社,2013.
[19]戚嘉敏,张鹏,奚如春,等.高州油茶果实性状的遗传变异特征[J].中南林业科技大学学报,2018,38(4):108-113.
[20]顾志建,孙先凤.山茶属17个种的核形态学研究[J].云南植物研究,1997,19(2):159-170.
[21]郭钰柬.越南油茶果实生长及种籽油脂肪酸含量变化分析[D].海口:海南大学,2020.
[22]莫泽乾.越南油茶的核型分析[J].广西植物,1990(1):31-32.
[23]邹玉玲.攸县油茶优良无性系评价及遗传多样性研究[D].北京:中国林业科学研究院,2021.
[24]蒋习林,杨济荣,段明伦,等.五柱滇山茶芽苗砧嫁接育苗技术[J].现代农业科技,2011(18):81-82.
[25]吕华飞,周丽华,顾志建,等.山茶属5种植物的核型研究[J].云南农业大学学报,1993,8(4):307-31361.
[26]中国科学院昆明植物研究所.云南山茶花[M].昆明:云南人民出版社,1981:3-5.
[27]冯国楣.云南山茶属植物的种类和利用[M]//全国油茶科研究协作组.油茶科研资料选编.昆明:昆明植物所,1980.
[28]曹慧娟,李天庆.一些山茶属(CamelliaL.)植物的细胞染色体研究[J].北京林业大学学报,1986(2):35-41.
[29]夏丽芳,顾志建,王仲朗,等.探讨云南山茶起源的一线曙光:野生二倍体类型在金沙江流域的发现[J].云南植物研究,1994,16(3):255-262.
[30]顾志建.云南山茶花四倍体的首次发现及其科学意义[J].植物分类学报,1997,35(2):107-116.
[31]李光涛.山茶属植物核型研究进展[J].中国野生植物资源,20020(5):9-14.
[32]杜宁.滇山茶复合多倍体基因组原位杂交研究[D].昆明:中国科学院昆明植物研究所,2004.
[33]杨桂英,何瀚,曹子林,等.3种不同倍性滇山茶的耐寒性研究[J].云南大学学报(自然科学版),2016,38(2):335-343.
[34]徐晓丹,邵维助,郑伟.滇山茶多倍体栽培品种的细胞倍性[J].林业科学,2018,54(9):44-48.
[35]李拓键,屈燕,王兵益,等.不同倍性云南山茶染色体核型分析[J].中南林业科技大学学报,2023,43(3):167-174.
[36]刘利勤.滇山茶多倍体复合体起源和演化的分子细胞遗传学研究[D].昆明:中国科学院昆明植物研究所,2009.
[37]甘沛华,李旦,沈德周,等.不同瓣型腾冲红花油茶的遗传多样性与遗传结构分析[J].西南林业大学学报(自然科学),2019,39(4):46-52.
[38]李铁柱,田大伦,乌云塔娜,等.四倍体油茶的鉴定及变异[J].林业科学,2009,45(3):150-154.
[39]王炜.滇山茶多倍化的转录组变异分析[D].昆明:昆明理工大学,2018.
[40]叶红莲,王连春.多指标综合选择野生怒江红山茶优良单株[J].中南林业科技大学学报,2016,36(6):66-69,74.
[41]孙静贤,杨时宇,杨雪娟,等.怒江红山茶(Camelliasaluenensis)3个居群核型分析[J].云南大学学报(自然科学版),2019,41(6):1246-1253.
[42]李彦福,霍达,王进.西南红山茶种质资源及开发利用[J].生物资源,2018,40(4):377-381.
[43]周文才,王仲伟,董乐,等.浙江红花油茶种实性状多样性分析[J].南京林业大学学报(自然科学版),20245(2):51-59.
[44]黄少甫,赵治芬,吴若菁,等.浙江红山茶色染体核型的分析[J].广西植物,1984(4):285-288.
[45]贾文庆.山茶花生殖生物学及倍性育种基础研究[D].北京:中国林业科学研究院,2015.
[46]陈维新,叶志云,梁盛业,等.山茶属十个物种染色体计数[J].广西林业科技,1987(2):19-22.
[47]王雅琴,黄少甫,徐炳声,等.长毛红山茶和长尾红山茶的核型分析[J].广西植物,1990,10(1):25-29.
[48]张志刚,何方.山茶属部分种的核型研究[J].经济林研究,1989,7(1):31-52.
[49]陈林,宋丽.流式细胞术的发展及在植物研究中的应用[J].生物工程学报,2023,39(2):472-487.
