‘库尔勒香梨’与‘翠冠’杂交后代花器若干性状的遗传学研究
2025-01-20薛大虎吴玉霞徐诗琪程嘉宝王言君张银军宋玟萱何天明张峰

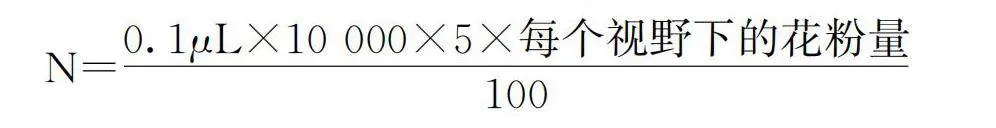


摘 要:【目的】对‘库尔勒香梨’与‘翠冠’及其杂交后代花器、花粉特征进行研究,以期为生产实践中梨授粉品种的选用、种质创新以及后期育种工作提供参考。【方法】以‘库尔勒香梨’与‘翠冠’及其杂交后代为试验材料,调查腋花芽比率、单花序花朵数、雄蕊数、花冠直径,连续两年测定花粉量和花粉活力,对其遗传规律进行分析,并对两年测定的花粉量和花粉活力进行显著性分析,对不散粉后代以及散粉量较少的部分后代进行了石蜡切片观察。【结果】在参与调查的杂交后代中,花粉量、花粉活力、雄蕊数虽然整体呈现退化趋势,但超高亲率分别为38.2%、11.1%和5.9%,都或多或少出现了超高亲后代,也有一定的选择优系的潜力;腋花芽比率在后代中有所降低,具有较高的抗寒和结高品质果实的潜力。【结论】‘库尔勒香梨’ב翠冠’杂交组合腋花芽比率表现出了一定的退化趋势,后代腋花芽占比减少,结优果概率提高;虽然其余3个性状或多或少也都表现出来一定的退化趋势,但都有超高亲后代存在,超高亲率最高的花粉量甚至达到了38.2%。同时后代变异系数都较高,变异系数最低的雄蕊数也在13.6%,性状分离较广泛,该组合具有较高的选育优系的潜力。雄性可育的亲本杂交,出现了正常发育型、少粒空壳型和完全败育型这3种育性的后代。
关键词:梨;杂交后代;花器;遗传规律
文章编号:2096-8108(2025)01-0001-07 中图分类号:S661.2 文献标识码:A文献标志码
Study on Genetics of Several Traits of Floral Organs in Hybrid Offspring of ‘Korla Fragrant Pear’ and ‘Cuiguan’
XUE Dahu1,WU Yuxia1,XU Shiqi1,CHENG Jiabao2,WANG Yanjun1,ZHANG Yinjun1,SONG Wenxuan1,HE Tianming1*,ZHANG Feng2
(1.College of Horticulture, Xinjiang Agricultural University, Urumqi Xinjiang 830052, China; 2.Korla Research Center of Fragrant Pear, Korlar Xinjiang 841003, China)
Abstract:【Objective】 The flower and pollen characteristics of ‘Korla Fragrant Pear’ and ‘Cuiguan’ and their hybrid offspring were studied in order to provide reference for the selection of pear pollination cultivars, germplasm innovation and later breeding work in production practice. 【Methods】 ‘Korla Fragrant Pear’‘Cuiguan’ and their hybrid offspring were selected as experimental materials. The axillary flower bud ratio, the number of flowers per inflorescence, the number of stamens, and the diameter of corolla were investigated. The pollen quantity and pollen viability were measured for two consecutive years, and the genetic law was analyzed. The significant analysis of the pollen quantity and pollen viability measured in two years was carried out. The paraffin sections of the offspring without pollen shedding and some offspring with less pollen shedding were observed. 【Results】 Among the hybrids involved in the survey, although the pollen amount, pollen viability, and stamen number showed a general trend of degradation, the ultra-high affinity were 38.2%, 11.1%, and 5.9%, respectively, and more or less ultra-high progeny offspring appeared, and there was also a certain potential for selecting superior lines. The ratio of axillary flower buds decreased in the progeny, with high cold resistance and high quality fruit potential. 【Conclusion】 In the hybrid combination of ‘Korla Fragrant Pear’ × ‘Cuiguan’, the ratio of axillary flower buds showed a certain degradation trend, the proportion of axillary flower buds in offspring decreased, and the probability of excellent fruit increased. Although the other three traits more or less also showed a certain trend of degradation, there were ultra-high progeny, and the pollen quantity with the highest their ultra-high affinity even reached 38.2%. At the same time, the coefficient of variation of the progeny was high, and the number of stamens with the lowest coefficient of variation was also 13.6%. The trait separation was extensive, and the combination had a high potential for breeding superior lines. There are three types of fertility progeny,normal development type, few grain empty shell type and complete abortion type by male fertile parents hybridization.
Keywords:pear; hybrid offspring; floral organ; genetic law
我国是梨(Pyrus Spp.)的起源中心之一,栽培历史悠久、种植范围广泛,具有丰富的梨种质资源[1]。梨是配子体型自交不亲和树种,自然条件下多数梨品种自花不结实或结实率低,栽培时要配置授粉品种或采用人工辅助授粉以促进坐果提高产量[2]。同时,花粉是研究植物遗传、育种的重要材料,所以,对梨花粉量和花粉活力进行研究,对产业发展具有十分重要的意义。目前,关于梨花粉量和花粉萌发率差异以及影响因素的研究已有较多报道[3-7],但对杂交后代花粉量和花粉萌发率及其遗传规律的研究较少,只有蒋媛[8]稍有提及,而在枣花上,潘依玲等[9]人对此做过较为深入的研究,对雄蕊数遗传变异的阐述更是集中在其他植物上,且材料为不同区域的种质资源,并非亲本与杂交后代的关系[10-11]。对植物花药进行解剖学研究,能更直观地看出花粉的发育情况,何天明[12]、马海英[13]、岳丽昕[14]等人分别在梨、锦绣杜鹃、大葱上进行过相关研究。因此,试验对‘库尔勒香梨’与‘翠冠’及其杂交后代花器特征进行了研究,探索其遗传规律,以期为生产实践中梨授粉品种的选用、种质创新以及育种研究工作提供参考。
1 材料和方法
1.1 试验地概况
试验于2023年—2024年在新疆库尔勒市沙依东园艺场新疆农业大学梨育种基地(东经 84°13′,北纬 41°47′)进行,该地属暖温带大陆性干旱气候,年降水量61 mm,年蒸发量2 800 mm,无霜期216 d,7月平均气温27 ℃,1月平均气温-7.1 ℃,年均温度12.2 ℃,有效日照时数2 844 h,≥10 ℃年有效积温4 894 ℃,土壤为戈壁荒漠灌淤土,pH值8.3,含盐量0.6%,土壤有机质含量为1.3%。
1.2 试验材料
供试的杂交后代母本为‘库尔勒香梨’,父本为‘翠冠’,砧木为‘杜梨’,定植行株距为5 m×3 m,常规管理。杂交工作于2011年在沙依东一分场进行。2012年在乌鲁木齐杂种圃实生育苗。2014年后陆续将实生接穗嫁接于库尔勒梨育种基地。
1.3 试验方法
1.3.1 腋花芽比率、雄蕊数的调查统计
在形态分化初期依据曹玉芬等[15]编著的《梨种质资源描述规范和数据标准》对亲本‘库尔勒香梨’‘翠冠’以及杂交后代腋花芽及顶花芽、雄蕊数进行调查统计,其中对25个具有3个及以上大枝的杂交后代进行了腋花芽比率调查,每个品种(株系)取3个大枝,计算腋花芽比率;对34个开花量较大的杂交后代进行了雄蕊数调查,每个品种(株系)取10朵大蕾期花朵调查雄蕊数。
1.3.2 花粉量的测定
分别对‘库尔勒香梨’‘翠冠’以及34个开花量较大的杂交后代进行了雄花粉量调查,取大蕾期花朵10朵左右,用解剖针剥离花药,随机取100枚发育完整、饱满的花药装到标记好的青霉素小瓶中,将完全散粉的上述青霉素小瓶中加入配置好的20%六偏磷酸钠溶液5 ml。充分震荡摇匀成悬浮液。血球计数板上盖好载玻片,用移液枪取上清液移取数滴,使其慢慢渗入血球计数板,用显微镜进行观察,重复6次,并计算出单花药花粉量N。
N=0.1μL×10 000×5×每个视野下的花粉量/100
1.3.3 花粉活力的测定
分别对‘库尔勒香梨’‘翠冠’以及27个开花量较大且可离体散粉的杂交后代进行了花粉活力调查,各品种(株系)花粉活力按何天明等[5]的方法测定,培养基用料及用量为蔗糖10 g、琼脂1 g、硼酸0.03 g、蒸馏水100 mL。
花粉活力/%=已萌发花粉数/花粉总数×100%
1.3.4 花药解剖
分别对‘库尔勒香梨’‘翠冠’以及34个开花量较大的杂交后代的花药进行了解剖,按试验所需用量将选定的梨品种枝条上大气球期花朵摘下,解剖针剥离花药,随机取20~30枚发育完整、饱满的花药固定在FAA (福尔马林-醋酸-酒精)固定液中,置于4 ℃冰箱内长期保存。采用石蜡切片法制片,切片厚度约8~12 μm,番红-固绿染色,用Nikon E-CLIPSE 80i显微镜进行观察并拍照。
1.3.5 计算公式
2 结果与分析
2.1 ‘库尔勒香梨’与‘翠冠’杂交后代腋花芽比率遗传变异
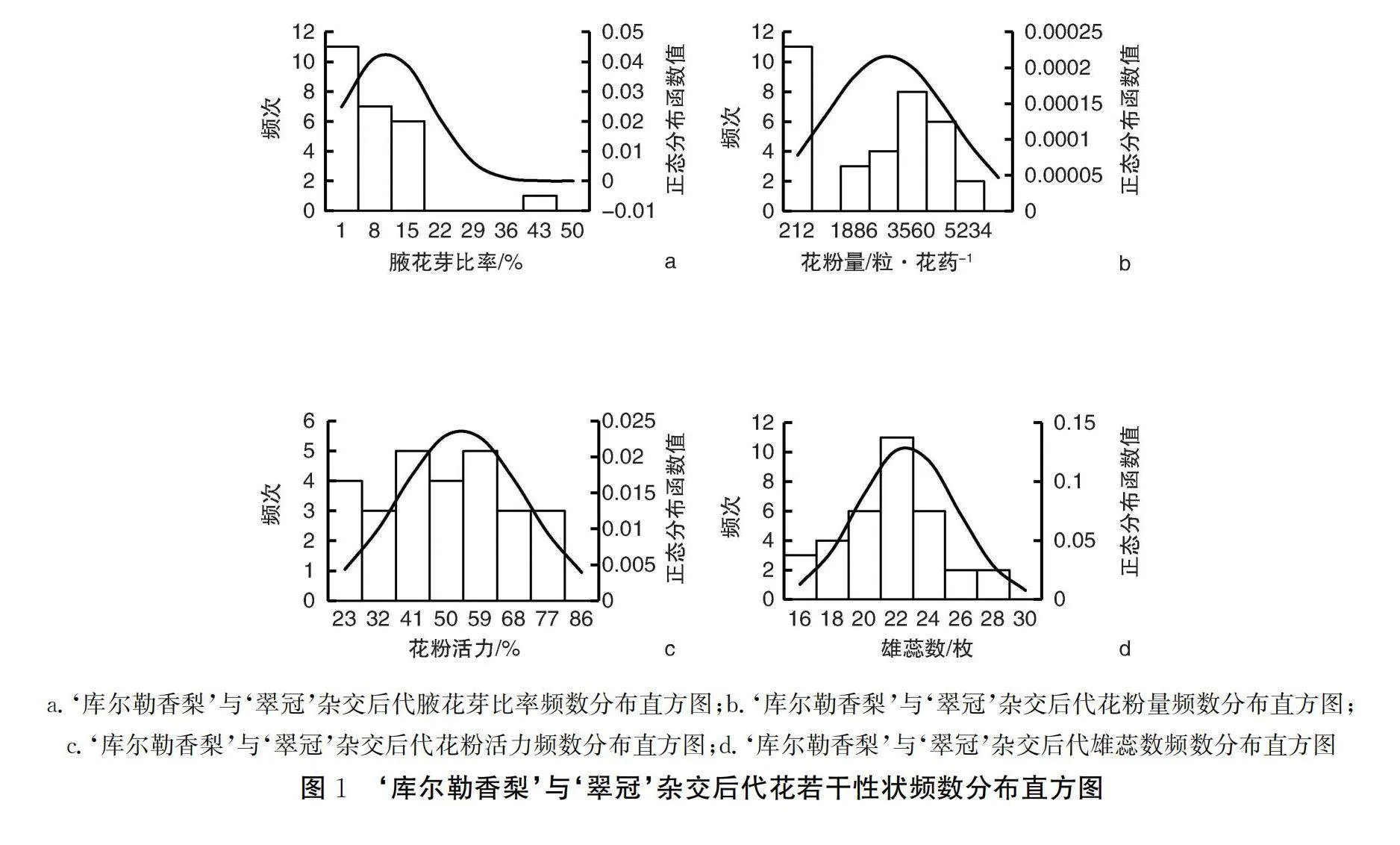
由表1可知,在参与调查的杂交后代中,腋花芽比率分布范围为1.0%~47.2%,超高亲率为8.0%,低低亲率为40.0%。杂种后代平均腋花芽比率为10.7%,与亲本相比较,要小于亲中值11.9%,大于低值亲本‘库尔勒香梨’。变异系数为86.7%,超亲优势率为-36.7%,中亲优势率为-10.1%,组合传递力为89.9%。数据比较离散,出现了较多的低低亲植株,克服了‘翠冠’腋花芽较多的特点。
由图1a可见,在‘库尔勒香梨’ב翠冠’参与调查的25株杂交后代中,除了1个后代单株腋花芽比率为47.2%(E-323)之外,其余均在22%以下。腋花芽比率在1%~8%范围内的最多,有11株,占比为44%;其次为8%~15%范围,占比为28%;15%~22%范围的占比为24%;43%~50%范围的最少,占比仅为4%。
2.2 ‘库尔勒香梨’与‘翠冠’杂交后代花粉量遗传变异
由表1可知,在参与调查的杂交后代中,花粉量分布范围为213~6 067粒·花药-1,超高亲率为38.2%,低低亲率为61.8%。杂种后代平均花粉量为2 846粒·花药-1,与亲本相比较,要小于亲中值3 816粒·花药-1,亦小于低值亲本‘翠冠’,变异系数为64.9%,说明后代分离极为广泛,超亲优势率为-25.6%,中亲优势率为-25.4%,组合传递力为74.6%,较亲本有所退化。数据比较离散,出现了较多的低低亲植株,花粉量在遗传过程中表现出了趋小方向的回归。
根据图1b,综合两年数据,两年共有34株散粉的不同杂交后代单株参与花粉量调查,花粉量集中分布在212~1 000粒·花药-1和3 560~5 234粒·花药-1区间。其中,花粉量在212~1 049粒·花药-1区间的杂交后代最多,占比32.4%;1 886~2 723粒·花药-1区间占比为8.8%;2 723~3 560粒·花药-1区间占比为11.8%;3 560~4 397粒·花药-1区间占比为23.5%;4 397~5 234粒·花药-1区间占比为17.6%;5 324~6 071粒·花药-1区间占比为5.9%。
2.3 ‘库尔勒香梨’与‘翠冠’杂交后代花粉活力遗传变异
由表1可知,在参与调查的杂交后代中,花粉活力分布范围为23.6%~82.1%,超高亲率为11.1%,低低亲率为88.9%。杂种后代平均花粉活力为50.4%,与亲本相比较,要小于亲中值76.1%,亦小于低值亲本‘翠冠’,变异系数为31.2%,说明后代分离广泛,超亲优势率为-33.9%,中亲优势率为-33.8%,组合传递力为66.2%,较亲本明显退化。数据比较离散,出现了许多的低低亲植株,花粉量在遗传过程中表现出了趋小方向的回归。
根据图1c,综合两年数据,至少有1年散粉的杂交后代共有27株,范围在23%~86%。花粉活力在41%~50%区间和59%~68%区间的杂交后代最多,分别占比18.5%;其次在23%~32%区间和50%~59%区间,分占比14.8%;在32%~41%、68%~77%和77%~86%区间的杂交后代最少,分别占比为11.1%。
2.4 ‘库尔勒香梨’与‘翠冠’杂交后代雄蕊数遗传变异
由表1可知,在参与调查的杂交后代中,雄蕊数分布范围为16.7~28.5枚,超高亲率为5.9%,低低亲率为67.6%。杂种后代平均雄蕊数为22.6枚,与亲本相比较,要小于亲中值26枚,亦小于低值亲本‘库尔勒香梨’,变异系数为13.6%,说明后代有所分离,超亲优势率为-19.9%,中亲优势率为-13.1%,组合传递力为86.9%,较亲本有所退化。数据比较离散,出现了许多的低低亲植株,雄蕊数在遗传过程中表现出了趋小方向的回归。
根据图1d,共有34株不同杂交后代单株参与雄蕊数调查,呈现出正态分布,集中分布在20~26枚区间。其中,雄蕊数在22~24枚区间的杂交后代最多,占比32.4%;其次在20~22枚区间和24~26枚区间,占比各为17.6%;18~20枚区间占比为11.8%;16~18枚区间占比为8.8%;26~28枚区间和28~30枚区间的杂交后代最少,占比各为5.9%。
2.5 ‘库尔勒香梨’与‘翠冠’及其杂交后代花药解剖研究
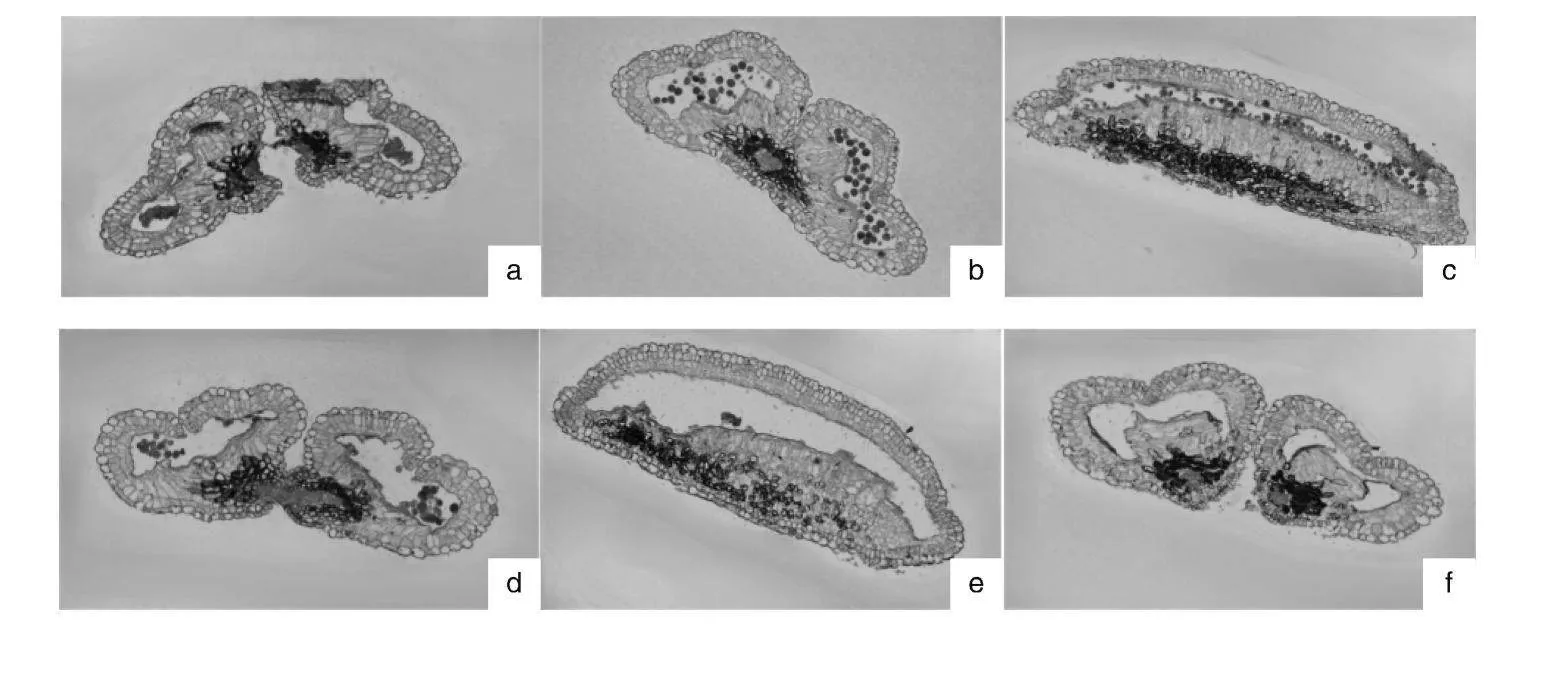
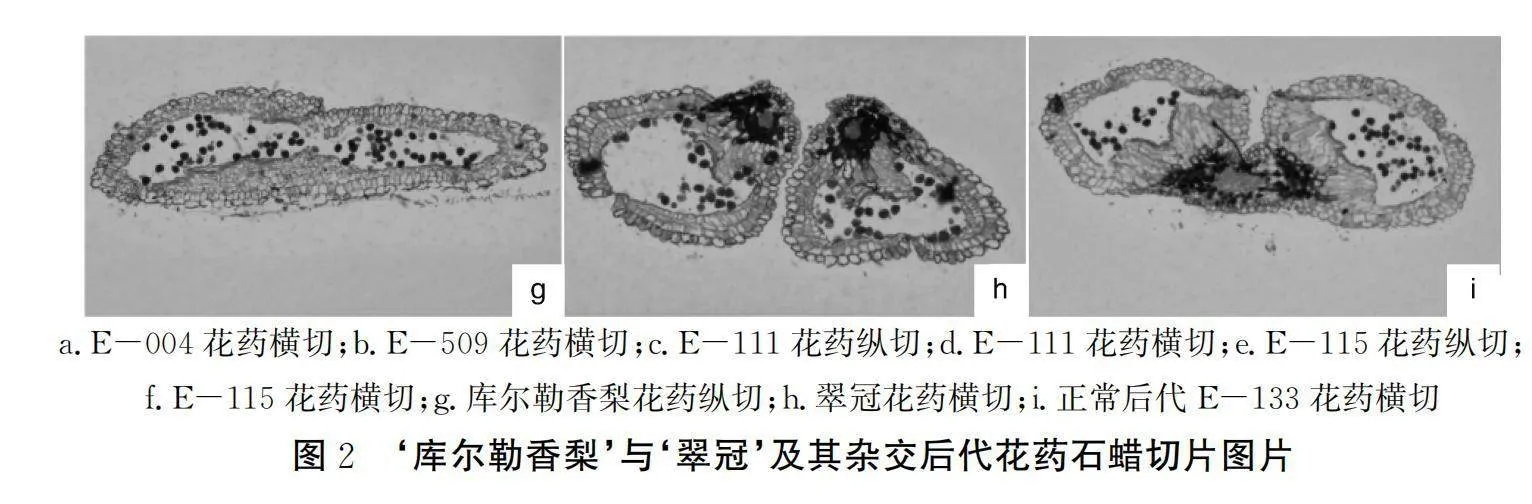
由图2可得,我们通过对后代进行石蜡切片观察,发现除了正常散粉的,还有有3种情况:药室内花药早已败育且药隔并不开裂,代表为E-004,同类型的E-115横切时药室内几乎空无一物,纵切还可以看出靠近药壁的内缘有少许花粉败育的残迹;虽然有部分发育正常花粉,但花粉粒大小不一,整齐度差,代表为E-509;虽然药隔能开裂,但药室内花粉早已败育,并无正常形态的花粉从药隔裂口散出,代表为E-111。
3 讨论
3.1 腋花芽比率的遗传
腋花芽多位于生长旺盛的新梢中上部叶腋间,其分化开始晚,母枝又处于营养生长状态,芽内养分不足,分化质量差,表现为芽体小、芽鳞少,翌年开花较晚、花瓣小、花柱短、花药少、子房小、坐果少、果个小和品质较差[16]。在王东升[17]对3个韩国梨品种的研究中,发现‘圆黄’‘晚秀’的单果质量均为顶花芽果实大于腋花芽果实,只有‘华山’相反,但差异并不显著,并且‘圆黄’‘晚秀’的可溶性固形物质量分数也是顶花芽果实大于腋花芽果实,由此可见,顶花芽果实的品质一般要高于腋花芽果实的品质。李含芬等[18]人的调查结果表明,在-15℃左右的低温条件下,梨腋花芽的受冻率最高。所以,腋花芽比率越低,果实品质越好,受冻害影响越低。本研究中,亲本和近乎全部杂交后代腋花芽比率要低于22%,杂交后代低低亲率为40.0%,出现了较多的腋花芽比率低于亲本的后代,表明该性状杂种优势较弱,即后代花芽中顶花芽具有较强的杂种优势,说明‘库尔勒香梨’ב翠冠’这一杂交组合的后代具有较高的抗寒和结高品质果实的潜力。
3.2 花粉量的遗传
植物花粉量是开花植物学特征性研究中的一项重要内容,各种果树花药内花粉量的多寡取决于花药发育的是否正常[19],其与品种本身的遗传特性有关[3],同时也会受到树体营养、花芽发育状况以及所在地气候条件的影响[17]。根据Henselek等[20]在杏上的研究,花粉量对授粉成功的概率有较为显著影响的,通过对花粉量的测定和探讨,可以为授粉树在生产中的选择提供参考依据,同时也为杂交育种和相关领域的研究提供参考依据。在本研究中,部分后代两年间花粉量差异较大,这点与刘志虎[19]及王晓庆[4]的研究结果相同,亦说明该性状易受到上述因素的影响。花粉量呈正态分布,具有数量性状遗传特征,这点与潘依玲等[9]在枣上的研究结果相同,但低低亲率为61.8%,出现了较多的低于低值亲本的后代,与之相悖。本研究中杂交后代花粉量超亲优势率和中亲优势率均为负值,组合传递力仅为74.6%,花粉量有减少趋势,这点与蒋媛等[7]在库尔勒香梨与早酥梨杂交后代中得到的结论一致。
3.3 花粉活力的遗传
花粉活力是花粉具有生长、萌发或发育的能力[21]。其不仅与花粉自身遗传特性有关,而且受外界环境如温度、相对湿度、贮藏介质、辐射、光照、空气质量等多重因素影响[3]。花粉活性的高低直接决定了授粉效果,花粉活性不高授粉将无意义[22]。花粉活力与坐果率成正相关[23],高花粉活力不仅能提高坐果率,对提高授粉效率也起着积极的作用,通过对花粉活力的测定和探讨,可以在实际生产中为授粉树的选择提供参考依据。本研究中,有一半后代花粉活力两年间差异显著,这点与王晓庆[4]对砂梨的测定结果相同。花粉活力呈正态分布,具有数量性状遗传特征,这点与潘依玲等[9]对枣的研究结果一致。低低亲率高达88.9%,组合传递力仅为66.2%,具有明显的退化趋势,与蒋媛等[8]结论相似。后代花粉活力自23.6%到82.1%不等,不同后代间花粉活力差异明显,这点与前人对不同品种的梨的花粉活力的研究结果一致[3,24]。
3.4 雄蕊数的遗传
雄蕊是被子植物花的雄性生殖器,是雄花的一部分,其作用是产生花粉。种子植物产生花粉的器官,由花丝和花药两部分组成[25]。花中雄蕊的数目因植物的种类不同而不同,雄蕊数的多寡,直接影响着花药的多寡,进而影响单花可生产花粉量的多寡。通过对雄蕊数的测定和探讨,可以为生产中授粉树的选取提供一定的参考。根据前人对望谟古茶树不同地区的种质资源的研究,变异系数不高,雄蕊数分离程度较低[9],与本研究后代之间的雄蕊情况类似。本研究中雄蕊数低低亲率高达67.6%,超亲优势率、中亲优势率均为负值,组合传递力86.9%,呈现出了下降的现象,具有一定的退化趋势,表明杂交的方式对于提高雄蕊数的潜力不高。
3.5 花药解剖
花药是产生花粉粒的地方,是雄蕊的主要部分,着生在花丝顶端,分为左右两半,中间以药隔相连。通过对花药进行石蜡切片观察,我们可以更直观地看到花药各结构以及花粉粒的发育情况。根据何天明等[12]的研究,梨花粉的育性可以分为3种——正常发育型、少粒空壳型和完全败育型。本研究中散粉不正常的杂交后代的花药切片中,以E-004为代表的,花粉完全败育,最终导致药囊干瘪,进而无法开裂,为完全败育型;而以E-509,为代表的,花粉量少,整齐度差,形状不规则,为少粒空壳型。这点与何天明等[12]的研究结果一致。而E-111,2023年花粉量为1 200粒·花药-1,但2024年为0粒·花药-1,但切片中又有明显的败育痕迹,初步推测2024年不散粉可能是受到了外在因素的影响,仍需后续跟进验证。
散粉异常后代,两年花粉量数据对比存在两种情况:即相差不大和差异显著。根据刘志虎等[19]的研究,树体发育强弱对花粉量具有密切的联系,因此对花粉败育是否为一个相对稳定的可遗传性状,环境对其影响的程度究竟有多大,我们仍需进一步研究和探讨。
4 结论
‘库尔勒香梨’ב翠冠’杂交组合,腋花芽比率表现出了一定的退化趋势,后代腋花芽占比减少,结优果概率提高;虽然其余3个性状或多或少也都表现出来一定的退化趋势,但都有超高亲后代存在,超高亲率最高的花粉量甚至达到了38.2%。同时后代变异系数都较高,变异系数最低的雄蕊数也在13.6%,性状分离较广泛,该组合具有较高的选育优系的潜力。
通过对该组合梨花药进行石蜡切片,并参考花粉量和花粉活力,雄性可育的亲本杂交,出现了正常发育型、少粒空壳型和完全败育型这3种育性的后代,但后代中的花粉败育是否为一个相对稳定的可遗传性状,环境对其影响的程度究竟有多大,我们仍需进一步研究和探讨,以期更好地解析梨雄性不育的遗传与机理。
参考文献
[1] 滕元文. 梨属植物系统发育及东方梨品种起源研究进展[J]. 果树学报,2017,34(3):370-378.
[2] 范净,陈启亮,杨晓平,等. 砂梨种质资源花粉量及花粉萌发率的遗传多样性分析[J]. 华中农业大学学报,2016,35(4):20-24.
[3] 葛彤,齐开杰,谢智华,等. 不同梨品种花粉量与活力差异的研究[J]. 江苏农业科学,2024,52(6):142-149.
[4] 王晓庆,骆军,张学英,等. 2008—2010年大棚与露地梨花粉生活力的比较[J]. 上海交通大学学报(农业科学版),2010,28(6):514-517.
[5] 何天明,张琦,程奇. 香梨花器特征及花粉生活力研究初报[J]. 新疆农业大学学报,2000,23(3):35-38.
[6] 戴雨沁,程梦雨,顾佩乾,等. 钙源对梨花粉萌发及花粉管生长的影响[J]. 江苏农业科学,2020,48(19):142-144,155.
[7] 陈迪新,李小静,赵静,等.开花前低温对梨花粉活力和花粉管伸长的影响[J].中国果树,2013(2):27-28.
[8] 蒋媛,位杰,张琦. 不同品种(系)梨花器官结构、花粉特性比较研究[J].河南农业科学,2015,44(3):100-107.
[9] 潘依玲,鲍荆凯,吴翠云,等. 雄性不育枣‘JMS2’ב交城5号’杂交F1代花性状遗传变异分析[J]. 西北农业学报,2023,32(12):1913-1921.
[10] 陈艳君,何立敏,白鼎臣,等. 望谟古茶树种质资源基于花果形态的遗传多样性分析[J/OL]. 分子植物育种,2024:1-14[2024-06-30]http://kns.cnki.net/kcms/detail/46.1068.S.20230825.1532.008.html.
[11] 钟敏,廖光联,李章云,等. 野生毛花猕猴桃雄花花器性状及SSR遗传多样性研究[J]. 果树学报,2018,35(6):658-667.
[12] 何天明,吴玉霞. 香梨杂种后代花粉发育类型及其花药的解剖学研究[J]. 西北植物学报,2011,31(11):2231-2234.
[13] 马海英,杨瑞,杨柳,等.锦绣杜鹃花药发育及散粉孔形成的形态与解剖学研究[J].广西植物,2024,44(4):721-729.
[14] 岳丽昕,王清华,刘泽洲,等.大葱雄性不育花药败育的形态学特征和细胞学研究[J].中国蔬菜,2023(9):58-68.
[15] 曹玉芬. 梨种质资源描述规范和数据标准[M]. 北京:中国农业出版社,2006.
[16] 程嘉宝. ‘库尔勒香梨’授粉品种的筛选[D]. 乌鲁木齐:新疆农业大学,2023.
[17] 王东升,郭献平,吴中营,等. 三个韩国梨品种在河南生长结果特性观察[J]. 果树学报,2016,33(S1):140-146.
[18] 李含芬,郭永丽,马春晖. 冬季低温对梨树花芽冻害的影响[J]. 北方园艺,2018(15):32-37.
[19] 刘志虎,何天明,钟芳. 梨花粉量的测定与分析[J]. 甘肃林业科技,2003,28(1):34-35,54.
[20] YUKI HENSELEK,ELISABETH J. EILERS,CLAIRE KREMEN, et al. Pollination requirements of almond (Prunus dulcis): combining laboratory and field experiments[J] Journal of Economic Entomology, 2018, 111(3):1006-1013.
[21] 刁永强,耿文娟,廖康,等. 新疆野生杏花粉量及花粉活力研究初报[J]. 新疆农业科学,2012,49(1):40-45.
[22] 孙佩光,程志号,李洪立,等. 大红火龙果花粉离体萌发条件的优化及其活力测定方法筛选[J]. 贵州农业科学,2020,48(5):112-115,173.
[23] 姬雅欣,袁迪,王念,等. 长山核桃花粉活力及坐果率初步研究[J]. 河南林业科技,2021,41(4):5-6,9.
[24] 程嘉宝,王春峰,薛大虎,等. 基于开花物候期和花粉质量初步筛选库尔勒香梨适宜的授粉品种[J]. 天津农业科学,2023,29(12):32-38.
[25] 冯丹丹.一个控制拟南芥小孢子发育基因的定位和雄性不育基因启动子的克隆[D]. 上海:上海师范大学,2008.
