不同施氮水平对冬油菜根际微生物群落特征的影响
2024-12-31刘东海张智梅亮贤乔艳刘东伟肖卓熙李菲胡诚
关键词: 氮肥; 冬油菜; 微生物群落结构; 功能预测
油菜对氮素需求量高[1],合理施用氮肥我国冬油菜可平均增产42.5%[2],但实际生产中依然存在施肥不足、过量或偏施的现象[3];氮肥过量施用降低了肥料利用率[4]、油菜产量和籽粒品质[5]。肥料被认为是导致土壤微生物群落改变的主要驱动因子。有报道指出,细菌和真菌多样性均随氮肥施用量增加呈降低趋势[6],且施氮量越高多样性的降低幅度越大[7]。张叶[8]的油菜盆栽试验结果则显示,不同施肥量对油菜根际真菌多样性和土壤细菌群落结构影响不显著。Peng 等[9]指出,施肥不仅显著影响油菜产量形成和养分供给,而且会导致根际微生物的优势种群和相对丰度发生变化,从而影响有机物矿化进程和养分供给水平。甘国渝等[10]指出,长期不同养分缺乏会显著改变冬油菜根际细菌和真菌群落的组成结构及其多样性。目前,关于施氮对微生物群落的具体影响尚未形成共识,因此在油菜主产区(武穴) 深入探究氮肥施用如何改变油菜根部周围土壤微生物群落的特征,具有重要科学意义。在植物根际微生态环境中,微生物显著影响着“土壤−植物”系统中矿质元素的生物有效性及植物生长环境的健康状况[11],同时根际微生物多样性差异反映了植物的健康和受胁迫程度[12]。Feng 等[13]分析了中国10 个长期(大于20 年)定位施肥试验,指出土壤中存在少量微生物类群对施肥的响应一致。这些物种或可作为指示生物来监测施肥对土壤肥力的影响。但是目前研究没有明确哪些物种可以作为土壤肥力的指示物种。因此,不同施氮肥条件下,油菜根际土壤中是否也存在关键物种可以作为指示土壤肥力的一个参考指标,从而指导施肥,有必要深入研究。
因此,本研究基于田间肥料定位试验来探明秸秆还田前提下,冬油菜根际细菌和真菌群落特征和功能对氮肥的响应,为冬油菜根际生态过程中的养分调控、土壤培肥和可持续发展提供依据。
1 材料与方法
1.1 试验地概况
试验地点位于湖北省武穴市(北纬29°54′17”,东经115°30′19′′),该地区属亚热带季风气候,年平均气温16.8℃,年平均降水量1409 mm,年平均日照1900 h。土壤类型为黄棕壤,属于世界粮农组织(World Food and Agriculture Organization, FAO) 分类中的漂白淋溶土(albic luvisol)。种植前0—20 cm 深度土壤基本性状为:土壤有机质30.8 g/kg,pH 5.5,碱解氮177.0 mg/kg,有效磷24.9 mg/kg,速效钾104.7 mg/kg。
1.2 试验设计
冬油菜–水稻轮作为该地区主要的种植制度,一年两熟。该地区灌溉排水条件良好。油菜试验从2019 年开始,每年9 上旬左右育苗,10 中下旬进行移栽,栽种密度为11.43 万株/hm2,次年5 月收获,秸秆全量还田。试验共5 个处理,分别为:不施氮肥(SN0)、N 120 kg/hm2 (SN120)、N 180 kg/hm2(SN180)、N 240 kg/hm2 (SN240) 和N 300 kg/hm2(SN300)。各处理重复 3 次,小区面积20 m2,随机区组排列。冬油菜氮肥50% 作为基肥,20% 在越冬期施用,30% 在抽苔期施用。磷、钾肥一次性基施,用量分别为P2O5 90 kg/hm2、K2O 120 kg/hm2。肥料种类分别为尿素(N 46%)、过磷酸钙(P2O5 12%)、氯化钾(K2O 60%)。统一其他田间管理。5 月20 日,油菜实际测产。
1.3 样品采样与测定
1.3.1 土壤样品采集及指标测定 2019 年10 月移栽油菜,共进行了4 年试验,于 2023 年4 月冬油菜盛花期采集土壤样品,各小区选取长势均匀的5 株油菜连根拔起,抖掉附着的松散土,用刀片采集根际土壤,作混合样。标记后装入无菌密封袋,放入有冰袋的冰盒带回实验室。一部分样品风干磨细过0.25 mm 筛,保存。测定土壤全氮、碱解氮、有效磷、速效钾、pH 和有机碳;以上测定方法均参考《土壤农化分析》[14]。另一部分放入−80℃ 冰箱保存,用于土壤微生物测序。
1.3.2 土壤微生物多样性分析 由上海美吉生物医药科技有限公司完成根系样品总 DNA 提取、PCR 扩增和序列测定。细菌16S rDNA V3~V4 区扩增采用前引物序列338F (5'-ACTCCTACGGGAGGCAGCAG-3')和后引物序列806R (5'-GGACTACNNGGGTATCTAAT-3'),真菌 ITS 扩增采用前引物序列 ITS1F (5'-CTTGGTCATTTAGAGGAAGTAA-3') 和后引物序列ITS2R(5'-GCTGCGTTCTICATCGATGC-3')。目标条带胶回收纯化后进行文库构建,利用 IlluminaMiSeqPE250平台进行高通量测序。
1.4 数据处理
测序原始数据去除引物接头序列和各低质量碱基后进行拼接,舍弃长度短于 160 bp 的序列,去除非特异性扩增序列及嵌合体后,得到每个样本的有效序列数据。以 97% 为规定阈值,对 16S 和 ITS 序列划分可操作分类单元(OTU)。采用 QIIME,调用UCLUST 序列对比工具按 97% 的序列相似度进行归并,并选取每个 OTU 度最高的序列作为该OTU 的代表序列。针对细菌 16S rRNA 和真菌 ITS rRNA 分别采用 Greengenes 数据库和 Silva 数据库作为 OTU分类地位鉴定的模板序列。
1.5 统计分析
试验数据采用 Excel 2019、IBM SPSS Statistics20 统计软件进行单因素方差分析,平均数据以“平均数±标准差(SD)”表示,采用 LSD 法检验 0.05 水平上的差异显著性。利用OriginPro 2019b 进行计算和作图。微生物群落组成和多样性以及环境因子和细菌或真菌群落的关联网络分析,利用上海美吉生物医药科技有限公司的I-sanger 云数据分析平台进行在线数据分析。
2 结果与分析
2.1 冬油菜根际土壤理化特征及产量
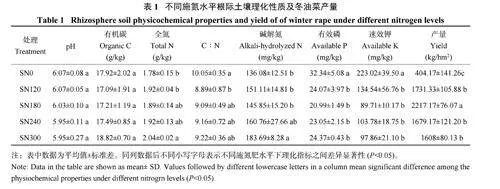
根际土壤理化性质分析结果(表1) 显示,与SN0 处理相比,随着施氮量的增加,pH 呈下降趋势。与SN0 处理相比,SN120、SN180、SN240 和SN300 处理根际土壤有效磷含量分别显著降低了25.27%、35.10%、28.73% 和24.63%,SN120、SN180、SN240 和SN300 处理速效钾含量分别显著降低了39.67%、59.77%、53.47% 和56.12%,全氮含量提高了6.18%~14.60% 和碱解氮含量提高了7.18%~34.99%。SN300 处理碱解氮含量显著提高了34.99%和全氮含量显著提高了14.61%。施氮肥降低了土壤碳氮比,却对有机碳的影响不显著。与SN0 处理相比,随着施氮量的增加,产量先增加后减少,SN180处理产量显著提高了448.57%,达到2217.17 kg/hm2。
2.2 冬油菜根际微生物多样性
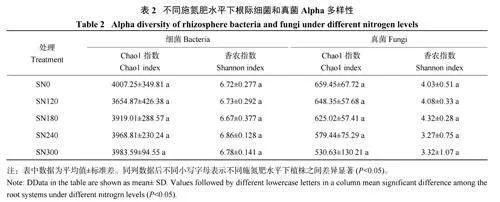
与SN0 处理比较,施氮处理的根际细菌Chao1指数和Shannon 指数虽没有显著变化,但细菌Chao1指数呈现降低趋势,其中SN120 处理的Chao1 指数最小,降低了8.79%;各施肥处理间,随着施氮量的增加,细菌Chao1 指数呈递增趋势。与SN0 处理相比,施氮处理根际土壤真菌Chao1 指数和Shannon指数虽无显著变化,但真菌Chao1 指数呈现降低趋势,SN300 处理的真菌Chao1 指数和Shannon 指数较小,S N 1 8 0 处理真菌S h a n n o n 指数最高,较SN0 处理提高7.20% (表2)。
2.3 冬油菜根际微生物群落组成
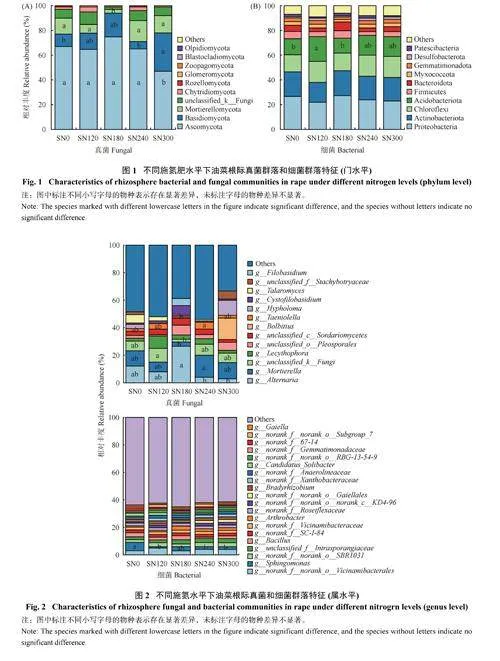
由图1 可知,不同施氮肥水平下,真菌的优势菌群主要是子囊菌门(Ascomycota)、担子菌门(Basidiomycota) 和被孢霉门(Mortierellomycota),三者占总体菌群相对丰度的84%~92%。与SN0 处理相比,SN180 处理提高了子囊菌门相对丰度10.45%,显著降低了被孢霉门相对丰度76.92%;SN300 处理显著降低了子囊菌门相对丰度30.0%,显著提高了担子菌门相对丰度210.0%。
细菌的优势菌群是变形菌门(Proteobacteria)、放线菌门(Actinobacteriota)、绿弯菌门(Chloroflexi)、酸杆菌门(Acidobacteriota)、厚壁菌门(Firmicutes) 和拟杆菌门(Bacteroidota),占总体菌群相对丰度的82%~86%,其中酸杆菌门作为油菜根际土壤中的优势菌门,占到了总菌门的16%~20%。与SN0 相比,施氮没有显著改变细菌群落组成,但SN120 处理显著提高了酸杆菌门的相对丰度53.85%,SN240和SN300 处理则分别提高了其相对丰度23.08% 和23.08%,SN180 处理则降低了其相对丰度7.69%。与SN0 处理相比,SN180 处理的优势菌群相对丰度基本相同,提高了拟杆菌门的相对丰度 (图1)。
由图2 可知,细菌属水平方面,与SN0 处理比较,SN120、SN180、SN240 和SN300 处理分别降低了鞘氨醇单胞菌属(Sphingomonas) 相对丰度75.01%、45.91%、67.48% 和66.29%,分别降低了芽孢杆菌属(Bacillus) 相对丰度36.20%、10.77%、56.07%和45.86%;SN180 处理减缓了鞘氨醇单胞菌属和芽孢杆菌属的降低趋势。施氮肥提高了节杆菌属(Arthrobacter) 相对丰度,其中SN180 和SN300 处理分别提高了122.35% 和82.08%;SN180、SN240 和SN300 处理分别降低了Candidatus_Solibacter 相对丰度28.53%、15.00% 和12.81%。SN120、SN240 和SN300处理分别提高了慢生根瘤菌属(Bradyrhizobium) 相对丰度26.43%、23.29% 和41.38%,但是SN180 降低了7.0%。
真菌属水平方面 (图2),与SN0 处理相比,SN180处理显著提高了链格孢属(Alternaria) 相对丰度116.67%,其他施氮处理提高不显著。SN180 中线黑粉酵母属(Filobasidium) 丰度最高(5.02%)。SN0 处理中篮状菌属(Talaromyces) 相对丰度最高(6.05%),SN300 处理中Bolbitius 相对丰度最高(16.14%)。
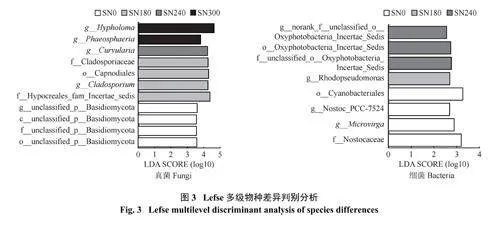
2.4 冬油菜根际微生物群落结构差异
由图3 可知,细菌方面,SN240 处理富集蓝菌门中的Oxyphotobacteria_Incertae_Sedis 目及其未知的科属。SN180 富集变形菌门中的红假单胞菌属(Rhodopseudomonas)。SN0 富集蓝菌门中蓝细菌目(Cyanobacteriales),念珠藻科(Nostocaceae) 和Nostoc_PCC-7524。真菌方面,SN0 富集担子菌门的一些未知菌类,SN240 富集弯孢霉属(Curvularia)。SN300富集球腔菌属(Phaeosphaeria) 和韧伞属(Hypholoma)。SN180 富集Hypocreales_fam_Incertae_sedis 科、枝孢属(Cladosporium)、煤炱目(Capnodiales) 和枝孢菌科(Cladosporiaceae)。
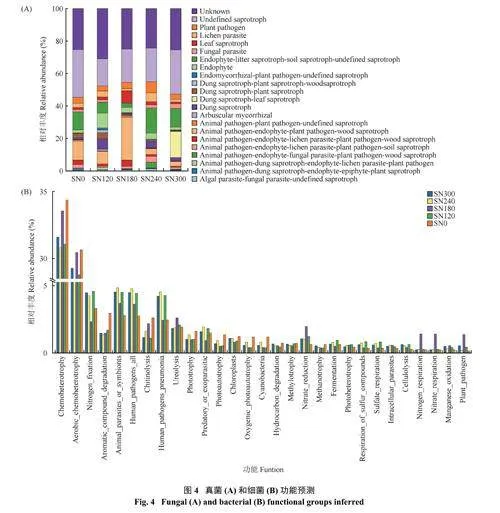
2.5 细菌和真菌功能预测
本研究借助FUN Guild 在线数据平台(https://github.-com/UMNFuN/FUNGild) 展开真菌功能预测分析(图4A)。主要营养型为腐生菌, 其次为病原菌和共生菌。与SN0 处理相比,SN180 处理显著提高了Animal pathogen-endophyte-plant pathogen-woodsaprotroph 菌群相对丰度124.47%,其他施氮处理则降低了34.33%~75.56%;SN240 处理显著提高了Plant Pathogen 菌群相对丰度80.55%,其他施氮处理变化不显著。SN120、SN180、SN240 和SN300 分别降低了Undefined Saprotroph 相对丰度43.17%、30.37%、29.67% 和7.34%,随着施氮肥用量的增加,Undefined Saprotroph 丰度呈现增加趋势。与SN0 比较,SN180 处理显著降低了Fungal parasite 相对丰度76.18%,SN300 处理显著提高了Fungal parasite相对丰度96.37%。与SN0 比较,SN120、SN180 和SN240 分别提高了Endophyte 相对丰度368.28%、66.54% 和90.18%,SN300 降低了Endophyte 相对丰度12.59%。
根据16S 序列的分类注释结果,采用FAPROTAX工具对细菌群落功能注释后展开分析(图4B)。相对丰度较高的功能分组前8 位分别为化能异养(chemoheterotrophy 和aerobic chemoheterotrophy)、固氮(nitrogen_fixation)、人体致病菌(human_pathogens_all 和human_pathogens_pneumonia)、动物寄生菌(animal_parasites_or_symbionts),尿素分解(ureolysis)、芳香化合物降解(aromatic_compound_degradation),几丁质分解(chitinolysis) 和光合营养(phototrophy) 功能类细菌,分别占总细菌群落的61.80% (32.28% 和29.52%)、3.80%、7.60% (4.00% 和3.60%)、4.07%、2.07%、1.80%、1.74% 和1.21%。与SN0 处理相比,施氮降低了化学异养、固氮、芳香化合物降解和几丁质分解功能菌群的丰度;提高了人体致病菌(human_pathogens_all 和human_pathogens_pneumonia)和动物寄生菌(animal_parasites_or_symbionts) 丰度。SN180 处理提高了硝酸呼吸作用(nitrate respiration)、氮呼吸作用(nitrogen respiration)、硝酸还原作用(nitrate-reduction) 和尿素分解(ureolysis),降低了固氮作用(nitrogen_fixation)。
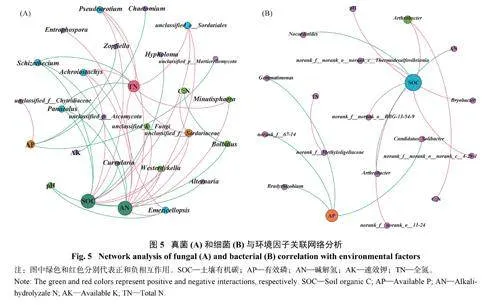
2.6 环境因子与土壤真菌和细菌优势菌属的关联网络分析
环境因子与土壤真菌、细菌优势菌属之间进行关联网络分析(Plt;0.02,|R2|gt;0.5),结果(图5) 显示,环境因子与真菌群落关联网络的平均度(2.89)、网络直径(6)、图密度(0.11) 和平均路径长(2.33) 均大于环境因子与细菌群落关联网络。环境因子与真菌群落关联网络分析(图5A) 得出,土壤有机碳(SOC)、碱解氮(AN)、全氮(TN)、有效磷(AP)、pH、C∶N 和速效钾(AK) 的平均度分别是11、11、8、4、2、2 和1。SOC 含量与Bolbitius、Hypholoma、Curvularia、Schizothecium 和Panaeolus 丰度呈显著正相关,与Pseudeurotium、Westerdykella、Achroiostachys以及3 个未分类真菌呈显著负相关。AN、TN 含量均与Schizothecium 和Panaeolus 相对丰度呈显著正相关,均与Minutisphaera、Pseudeurotium、Emericellopsis和Achroiostachys 等呈显著负相关;AN 含量还与Westerdykella、Entrophospora 以及未分类的真菌菌属呈显著负相关。AP 含量与Chaetomium 和Zopfiella呈显著正相关。pH 与Alternaria、Emericellopsis 呈显著正相关。C∶N 与Bolbitius 呈显著正相关。
环境因子与细菌群落关联网络分析(图5B) 得出,SOC、AP、AN、TN、AK 含量和C∶N 的平均度分别是6、4、1、1、1 和1。SOC 与Nocardioides显著正相关,与Anaeromyxobacter 以及4 个不确定的细菌属显著负相关。AP 含量与Bradyrhizobium、G e m m a t i m o n a s 等4 个属呈显著正相关。pH 与Bryobacter 呈显著负相关。C∶N 与Anaeromyxobacter呈显著负相关。
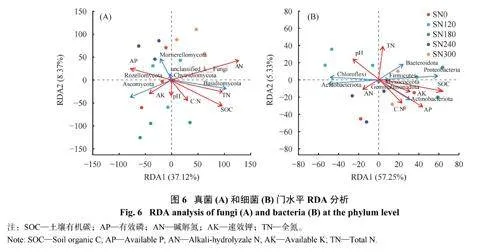
2.7 土壤真菌和细菌群落的RDA 分析
环境因子与真菌和细菌群落的冗余分析结果见图6 。真菌RDA 分析结果(图6A) 显示,第一轴和第二轴解释度合计45.49%,SOC、AN、TN 和AP是影响真菌群落的主要环境因子。细菌RDA 分析结果(图6B) 显示,SOC 和AP 是影响细菌群落的主要环境因子。
3 讨论
3.1 土壤理化特征和产量对施氮肥的响应
与SN0 比较,施氮肥未显著降低pH,随着氮肥用量的增加,pH 呈现下降趋势,与唐贤等[15]研究的结论一致。与SN0 比较,随着施氮量的增加,产量先增加后减少,SN180 产量显著提高了448.57%,产量最高,作物从土壤中吸收的磷钾元素大于其他施氮处理, 因此, 与其他施氮处理比较, S N 1 8 0处理的土壤中有效磷和速效钾含量最低。从产量角度而言,SN180 是适宜的氮肥用量。
3.2 根际微生物对施氮肥的响应
Chao1 和Shannon 指数越大,说明样品的物种丰富度和多样性越高。Yang 等[16]指出,施氮降低了不同生态系统土壤细菌多样性(−11%) 和真菌多样性(−17%)。靳玉婷等[17]也指出,秸秆还田配施化肥油菜季土壤细菌的Chao1 指数和Shannon 多样性指数有所降低。本研究中,与SN0 处理比较,施氮肥均减少了细菌Chao1 指数,差异不显著,随着氮肥用量的增加,细菌的Chao1 指数成递增趋势。Zhou 等[18]指出,长期施用氮肥和磷肥降低了真菌多样性,且随着化肥用量的增加,对真菌群落的影响加大。本研究中,与SN0 处理比较,SN180 提高了真菌Shannon指数7.20%,其他氮肥处理真菌Shannon 指数呈下降趋势,说明合理的氮肥用量能够维持真菌多样性。
Wang 等[19]指出,施用无机肥降低了子囊菌门丰度。过量氮肥对子囊菌门是有害的[20]。本研究SN180提高了子囊菌门相对丰度10.45%,SN120、SN240和SN300 处理降低了子囊菌门相对丰度。子囊菌门大多数为腐生菌,可以分解难降解性有机质,促进土壤碳的吸收利用[21]。酸杆菌门作为油菜根际土壤中的细菌优势菌门,占总体菌群相对丰度的16%~20%;Kielak 等[22]和Han 等[23]研究发现,土壤酸杆菌能够进行复杂的碳降解,可能有助于秸秆的降解。本研究中,SN120、SN240 和SN300 处理显著提高了酸杆菌门的相对丰度,SN180 处理降低了酸杆菌门的相对丰度。Dai 等[24]通过对全球数据的荟萃分析指出,长期施氮肥并不一定增加酸杆菌门的相对丰度,与施用的养分投入有关。许多鞘氨醇单胞菌具有固氮、溶解磷酸盐、产生植物生长激素等功能,能够促进植物生长和提高植物抗逆性[25−26];芽孢杆菌属也能促进植物生长[27]。在各施氮处理中,SN180 处理减缓了根际土壤鞘氨醇单胞菌和芽孢杆菌丰度的降低。施肥均提高了真菌的链格孢属丰度,链格孢属是引起植物病害的重要真菌类群之一[28],可能会增加患植物病害的风险。SN180 处理根际土壤富集线黑粉酵母属 (Filobasidium);Keszthelyi 等[29]发现Filobasidium 能够分泌一种针对致病真菌Cryptococcusneoformans 的毒素,可作为生物防治剂。SN180 处理富集变形菌门中的红假单胞菌属,其属于非共生固氮菌[30],可提高土壤固氮潜力和固氮菌抗逆性[31]。因此,SN180 处理可能存在致病菌和拮抗菌的某种平衡,还需深入研究。慢生根瘤菌属是兼性固氮菌[32],来自土壤和非豆科植物的菌株缺乏共生固氮需要的nif 和nod 基因[33],因此施肥促进了慢生根瘤菌属的营养生长,而不是固氮[34]。SN120、SN240 和SN300处理分别提高了土壤慢生根瘤菌属(Bradyrhizobium)相对丰度,但是SN180 处理降低其相对丰度。这可能有两个原因:1) 与氮肥投入量有关;2) 氮肥的适宜用量促进油菜生长和提高产量,加大了油菜对土壤养分的吸收,导致土壤有效磷含量较低,本研究中慢生根瘤菌属(Bradyrhizobium) 与有效磷(AP) 存在显著正相关。
3.3 环境因子与群落关联网络分析
环境因子与群落关联网络分析和RDA 分析均显示,SOC、AN、TN 和AP 是真菌群落的主要影响因子,SOC 和AP 是细菌群落的主要影响因子。无色穗孢属(Achroiostachys) 与SOC 负相关,与AN 正相关。无色穗孢属均为土壤致病真菌,对作物生长具有不利影响,并降低作物产量[35]。Emericellopsis 与AN和TN 显著负相关,与pH 显著正相关。Emericellopsis属的很多物种被认为是生物活性代谢物的重要来源,例如Emericellopsis cladophorae 能够产生一系列抗真菌衍生化合物等[36]。说明过量施氮肥可能促进有害菌的生长,提高土壤有机碳能够抑制有害菌的生长。
厌氧粘细菌缺乏反硝化作用中的nirS 和nirK 基因, 只能还原NO3−-N 生成NH4+-N 或者还原N2O 生成N2 来获得生长所需的能量,因此可能不利于土壤中氮素的固存[37]。类诺卡氏菌属是我国黄土高原地区的优势固碳微生物[38],可通过3-羟基丙酸双循环与还原性三羧酸循环实现大气CO2 的固定[39];本研究发现SOC 与类诺卡氏属(Nocardioides) 显著正相关,与厌氧粘细菌属(Anaeromyxobacter) 显著负相关。因此,秸秆还田下合理的氮肥投入对微生物固氮和固碳过程有一定影响。
4 结论
冬油菜施 N 180 kg/hm2 显著提高了油菜籽产量,减少磷、钾养分在土壤中的富集,同时提高了真菌Shannon 指数,减缓了有益菌(鞘氨醇单胞菌属和芽孢杆菌属) 下降趋势,富集了非共生固氮菌(红假单胞菌属) 和线黑粉酵母属(Filobasidium)。但是根际土壤中微生物群落复杂,目前研究方法存在局限性,关键物种难以作为土壤肥力的一个参考指标。有机碳、碱解氮、全氮和有效磷是真菌群落的主要环境因子,有机碳和有效磷是细菌群落的主要影响因子。施氮显著影响油菜根际微生物群落特征,过量施用氮肥导致病害微生物丰度增加,综上所述,施N 180 kg/hm2 是种植油菜比较合理的施氮量。
