氮磷钾肥对有柄石韦生理及绿原酸合成积累的影响
2024-06-27步园园潘志芳崔祥祥周笑陈建桦黄荣韶李良波
步园园 潘志芳 崔祥祥 周笑 陈建桦 黄荣韶 李良波

DOI: 10.11931/guihaia.gxzw202306012
步园园, 潘志芳, 崔祥祥, 等, 2024.
氮磷钾肥对有柄石韦生理及绿原酸合成积累的影响 [J].
广西植物, 44(5): 972-980.
BU YY, PAN ZF, CUI XX, et al., 2024.
Effects of nitrogen, phosphorus and potassium fertilizers on physiology and chlorogenic acid synthesis and accumulation of Pyrrosia petiolosa [J].
Guihaia, 44(5): 972-980.
摘 要: 为探讨氮磷钾3种养分对有柄石韦生理及有效成分绿原酸合成积累的影响,该研究以有柄石韦组培苗为材料,分别用低养分(不施肥:N0,P0,K0)、正常施肥(N:0.20 g·kg-1,P:0.15 g·kg-1,K:0.15 g·kg-1)和高养分(N1:0.40 g·kg-1,P1:0.30 g·kg-1,K1:0.30 g·kg-1)3个浓度梯度,设置7个处理分别为NPK、N0PK、N1PK、NP0K、NP1K、NPK0、NPK1,测定不同处理下有柄石韦的抗性生理指标、绿原酸(CGA)含量及其合成关键酶活性。结果表明:(1)氮磷钾肥对有柄石韦的抗性生理有显著的影响,超氧化物歧化酶(SOD)在高氮和低钾处理中活性显著增加,而3种养分的低浓度和高浓度处理均会导致过氧化氢酶(CAT)活性显著上升。(2)不同养分水平氮、磷和钾对有柄石韦CGA含量存在显著影响,正常施肥的CGA含量最高,达到12.92 mg·g-1;高钾施肥的CGA含量最低,为7.79 mg·g-1;钾肥对CGA含量影响最显著。(3)CGA合成关键酶活性在不同施肥处理中差异显著,CGA含量与奎宁酸羟基肉桂酰转移酶(HQT)和4-香豆酰辅酶A连接酶(4CL)活性呈显著正相关,与莽草酸羟基肉酰转移酶(HCT)活性显著负相关,HQT、4CL和HCT是导致CGA含量差异的关键因素。该研究结果为有柄石韦药材的人工栽培提供了理论依据。
关键词: 有柄石韦, 氮磷钾肥, 生理, 绿原酸, 积累
中图分类号: Q945
文献标识码: A
文章编号: 1000-3142(2024)05-0972-09
收稿日期: 2023-09-25 接受日期: 2023-10-19
基金项目: 国家自然科学基金地区项目(32160085); 广西中医药大学桂派中医药传承创新团队项目(2022B005); 广西中医药大学研究生创新计划项目(YCSY2023005); 江西红星药业有限公司委托项目。
第一作者: 步园园(1998—),硕士研究生,研究方向为中药资源开发与利用,(E-mail)1609299682@qq.com。
*通信作者: 李良波,博士,研究员,研究方向为道地药材活性成分积累机制及质量控制,(E-mail)llb100@126.com。
Effects of nitrogen, phosphorus and potassium fertilizers
on physiology and chlorogenic acid synthesis and
accumulation of Pyrrosia petiolosa
BU Yuanyuan1, PAN Zhifang1, CUI Xiangxiang1, ZHOU Xiao1,
CHEN Jianhua1,2, HUANG Rongshao1,2, LI Liangbo1,2*
( 1. College of Pharmacy, Guangxi University of Chinese Medicine, Nanning 530200, China; 2. Key Laboratory of Protection and
Utilization of Chinese Medicine Resources, Guangxi University of Chinese Medicine, Nanning 530200, China )
Abstract: To explore the effects of nitrogen, phosphorus and potassium on the physiology of Pyrrosia petiolosa as well as the synthesis and accumulation of chlorogenic acid (CGA), tissue culture seedlings of P. petiolosa were used as materials, and three concentration gradients of low nutrient ( no fertilization: N0, P0, K0 ), normal fertilization ( N: 0.20 g·kg-1, P: 0.15 g·kg-1, K: 0.15 g·kg-1 ) and high nutrient ( N1: 0.40 g·kg-1, P1: 0.30 g·kg-1, K1: 0.30 g·kg-1) were set up. Seven treatments were set up to NPK, N0PK, N1PK, NP0K, NP1K, NPK0 and NPK1, and the resistance physiological indexes, CGA contents and key enzyme activities of P. petiolosa under different treatments were determined. The results were as follows: (1) Nitrogen, phosphorus and potassium fertilizers had significant effects on the resistance physiology of P. petiolosa. The activities of superoxide dismutase(SOD) increased significantly under high nitrogen and low potassium treatments, but catalase (CAT) activities rose significantly under low and high treatments of three nutrients. (2) The contents of CGA in P. petiolosa was significantly affected by different nutrient levels of nitrogen, phosphorus, and potassium. The CGA level in normal fertilization was the highest, reaching 12.92 mg·g-1, while the CGA content in high potassium fertilization was the lowest, 7.79 mg·g-1. Potassium fertilizer had the most significant effect on CGA content. (3) The key enzyme activities of CGA synthesis were significantly different in different fertilization treatments. The content of CGA was positively correlated with the activities of quinate o-hydroxycinnamoyl transferase (HQT) and 4-coumaroyl coenzyme ligase (4CL), and negatively correlated with the activity of quinic acid shikimate o-hydroxycinnamoyl transferase (HCT). HQT, 4CL and HCT were the key factors leading to the difference of CGA content. The results of this study provide a theoretical reference for the artificial cultivation of P. petiolosa.
Key words: Pyrrosia petiolosa,nitrogen,phosphorus and potassium fertilizers, physiology, chlorogenic acid (CGA), accumulation
有柄石韦(Pyrrosia petiolosa)为水龙骨科石韦属植物,是2020版《中国药典》收录的石韦药材基原之一,以叶入药,味苦、甘,性微寒,具有利水通淋、清肺泄热、止血等功效(国家药典委员会,2020)。潘礼业等(2020)研究表明,有柄石韦含有酚酸类、黄酮类、三萜类等多种类型的有效成分,而绿原酸(chlorogenic acid, CGA)是其质量控制的指标成分。一方面,有柄石韦基本来源于野生资源,随着野生资源的过度采挖和临床需求量的不断增加,有柄石韦野生资源急剧减少,供需矛盾日益突出;另一方面,由于各地石韦药材质量参差不齐,无法保证稳定的临床疗效(赖海标等,2010)。因此,为满足石韦药材的市场需求和保证药材质量的稳定,开展有柄石韦的引种驯化和人工栽培已势在必行。
目前,关于有柄石韦的人工栽培刚刚开始,仍存在孢子萌发率低、出苗整齐度差和存活率低、自然生长缓慢、施肥配比不明等问题。施肥是药用植物人工栽培及规范化种植过程中的关键环节之一,直接影响药用植物的生长、产量和质量。根据笔者调查,目前未见关于有柄石韦人工栽培中科学施肥方法的文献报道。因此,如何通过施肥管理调控有柄石韦的生长和有效成分积累是实现有柄石韦人工栽培需要解决的关键问题之一。
大量研究表明,合理的施肥能有效促进药用植物的生长发育和有效成分的合成积累,而滥用肥料或缺肥会形成逆境胁迫环境,破坏体内的代谢平衡,导致减产或药材有效成分含量降低。韩建萍等(2004)研究了氮、磷肥对丹参的次生代谢产物的影响,发现氮素对丹参酮ⅡA的积累表现出负面效应,丹参酮ⅡA含量随着施氮量的增加而逐渐减少,磷的施用对丹参酮ⅡA和丹参素的积累则具有积极促进作用。曹欣欣等(2020)研究发现,氮肥对西洋参皂苷影响变化明显,适当的氮肥用量显著增加了三七皂苷Rb3含量,而氮肥施用过多则会导致含量降低。有柄石韦的有效成分为绿原酸,属于酚酸类成分,其在植物体内的合成途径为苯丙烷代谢途径(Clifford et al., 2017),该途径的前体物质为芳香族氨基酸苯丙氨酸和酪氨酸,氮供应量可通过影响芳香族氨基酸的含量从而影响苯丙烷次生代谢产物的含量(Fritz et al., 2006)。张宁洁等(2022)研究发现,金银花绿原酸(CGA)的含量与施氮量呈负相关,适量施肥后金银花中苯丙氨酸氨解酶基因(PAL)、肉桂酸-4-羟化酶基因(C4H)、4-香豆酰辅酶A连接酶基因(4CL)和羟基肉桂酰转移酶基因(HQT)等CGA合成关键基因表达水平上调。有研究指出,磷素也会影响酚酸类代谢产物的合成积累,参与酚酸和类黄酮生物合成途径的基因PAL、4CL在磷胁迫下的不同表达,在高磷处理下异绿原酸C、隐绿原酸、异绿原酸B等酚酸含量显著降低(Wang et al., 2023)。与氮和磷不同,钾不是有机化合物的一部分,但钾作为主要阳离子或各种酶的辅助因子在植物的生理和生化过程中起着重要作用,如酶活化、离子稳态、渗透调节和蛋白质合成等(Xue et al., 2022)。然而,目前尚未有任何关于施肥水平影响有柄石韦酚酸类活性成分合成积累的研究报道。
为了探讨有柄石韦对不同氮磷钾肥浓度的生理响应及绿原酸(CGA)合成积累的规律,本研究测定了不同浓度氮磷钾处理下有柄石韦的抗氧化酶SOD、CAT及渗透调节物质脯氨酸含量的变化,以及CGA合成关键酶苯丙氨酸转氨酶(PAL)、肉桂酸-4-羟化酶(C4H)、4-香豆酰辅酶A连接酶(4CL)、莽草酸羟基肉酰转移酶(HCT)、奎宁酸羟基肉桂酰转移酶(HQT)、香豆酸-3-羟化酶(C3H)的活性和CGA的含量,以期阐明有柄石韦对不同浓度氮磷钾肥的生理响应及CGA合成积累规律,为有柄石韦的引种驯化和人工栽培提供理论依据。
1 材料与方法
1.1 供试材料
本试验于2022年7—9月在广西中医药大学温室大棚内进行,供试植株为1年生有柄石韦组培苗,选取生长状态一致的有柄石韦植株,将根部冲洗干净后用1 000倍多菌灵浸泡根部15 min,移栽后置于温室大棚内培养。使用尿素(津欧博凯化工有限公司)、磷酸二氢钙(天津市大茂化学试剂厂)和氯化钾(天津市北辰方正试剂厂)分别对有柄石韦进行氮磷钾养分处理。
1.2 供试土壤
供试土壤为酸性红色土壤,土壤经灭菌、风干、打碎、混匀后用于盆栽试验。混合基质为红土∶育苗基质(有机质+腐殖酸)∶珍珠岩=2∶2∶1,混合基质理化性质为 pH 4.9、全氮0.10%、全磷0.12%、全钾2.50%、有机质 16.0 g·kg-1、水解性氮73.0 mg·kg-1、速效磷8.8 mg·kg-1、速效钾247 mg·kg-1。
1.3 试验设计
施肥使用分析纯的尿素、磷酸二氢钙和氯化钾,氮磷钾3种主要营养分别用低养分(不施肥)、正常养分和高养分3个浓度梯度,共设置7个处理组分别为NPK、 N0PK、 N1PK、 NP0K、 NP1K、NPK0、NPK1,其中氮处理为 N0PK、NPK、N1PK;磷处理为NP0K、NPK、 NP1K;钾处理为NPK0、NPK、NPK1。 N0、P0和K0为不施肥,正常养分施肥(N:0.20 g·kg-1,P:0.15 g·kg-1,K:0.15 g·kg-1),高养分施肥(N1:0.40 g·kg-1,P1:0.30 g·kg-1,K1:0.30 g·kg-1)。将有柄石韦植株移栽于栽培盆(长30 cm, 宽15 cm, 高10 cm)中,每个盆装2.8 kg混合基质。每个处理设置3次重复,每个重复15株苗。分两次施肥,0 d施一半肥,30 d施另一半肥,处理60 d,每隔3 d自来水浇灌1次,其他管理一致。
1.4 生理指标测定
超氧化物歧化酶(SOD)活性采用氯化硝基四氮唑蓝(NBT)光还原法(李合生,2000);用万分之一天平称取0.100 g的样品,加入1 mL提取液冷冻研磨,以8 000 r·min-1在4 ℃下离心10 min后取上清液,上清液处理后于酶标仪560 nm下测定样品的吸光度,当黄嘌呤氧化酶偶联反应体系中抑制百分率为50%时,反应体系中的SOD酶活力定义为一个酶活力单位。过氧化氢酶(CAT)活性采用分光光度法测定(赵亚华,2000),酶提取步骤同上,每克组织在反应体系中每分钟催化1 μmol H2O2定义为一个酶活单位。脯氨酸含量测定采用茚三酮法测定(高俊凤,2006)。
1.5 CGA含量测定
CGA含量的测定参照《中国药典》(国家药典委员会,2020)并进行稍作改动。将药材烘干粉碎后过筛,称取有柄石韦粉末0.2 g,置于50 mL离心管中,加入50%的甲醇15 mL,静置24 h后再加入50%的甲醇10 mL二次提取,药渣与提取液超声处理45 min后放冷至室温补足重量,10 000 r·min-1常温离心10 min后取上清液0.22 μm微孔滤膜过滤后待测。精密称定CGA标准品(北京索莱宝科技有限公司,货号:929 N022)2.17 mg,使用50%甲醇配置浓度为0.217 mg·mL-1的标准品溶液,分别稀释成浓度为0.021 7、0.043 4、0.086 8、0.130 2、0.173 6、0.217 0 mg·mL-1的标准曲线,以对照品浓度为横坐标x,峰面积为纵坐标y,线性回归方程为y=3.004 24×107x-329 43,R2=0.999 2。色谱柱:XBridge C18(4.6 mm × 250 mm,5 μm),检测波长326 nm,柱温25 ℃,流速1.0 mL·min-1,乙腈(C)~0.1%磷酸溶液(A)梯度洗脱,洗脱程序: 0~5 min,10%→15%C,90%→85%A;5~12 min,15%→19%C,85%→81%A;12~18 min,19%→27%C,81%→73%A;18~40 min,27%→80%C,73%→20%A;40~60 min,80%→10%C,20%→90%A。
1.6 CGA合成关键酶活性测定
CGA合成关键酶均采用酶联免疫法,按照试剂盒(广西君奇生物科技有限公司,202301)方法步骤进行测定,精确称取有柄石韦鲜样0.1 g ,加入1 mL的PBS(pH为7.2~7.4)缓冲液,4 ℃冷冻研磨匀浆,10 000 r·min-1、4 ℃离心20 min,获得酶提取液。样品孔加入样品稀释液,空白孔不加,除空白孔外标准孔和样品孔分别加入辣根过氧化物酶(HRP)标记的检测抗体,恒温箱37 ℃反应60 min后取出,弃去废液加入洗涤液重复洗涤5次,拍干后每孔加入底物,37 ℃避光孵育15 min后每孔加入硫酸终止液,测定450 nm下各孔的OD值。关键酶活性定义为每克蛋白质在每分钟内降解1 μmol 四甲基联苯胺(TMB)为一个酶活性单位。
1.7 数据分析
用Microsoft Excel 2019软件处理数据(数据以平均值±标准差表示),用SPSS 25软件对数据进行方差分析、多重比较、Pearson相关性分析,Origin 2021软件作图。
2 结果与分析
2.1 氮磷钾肥对有柄石韦抗性生理的影响
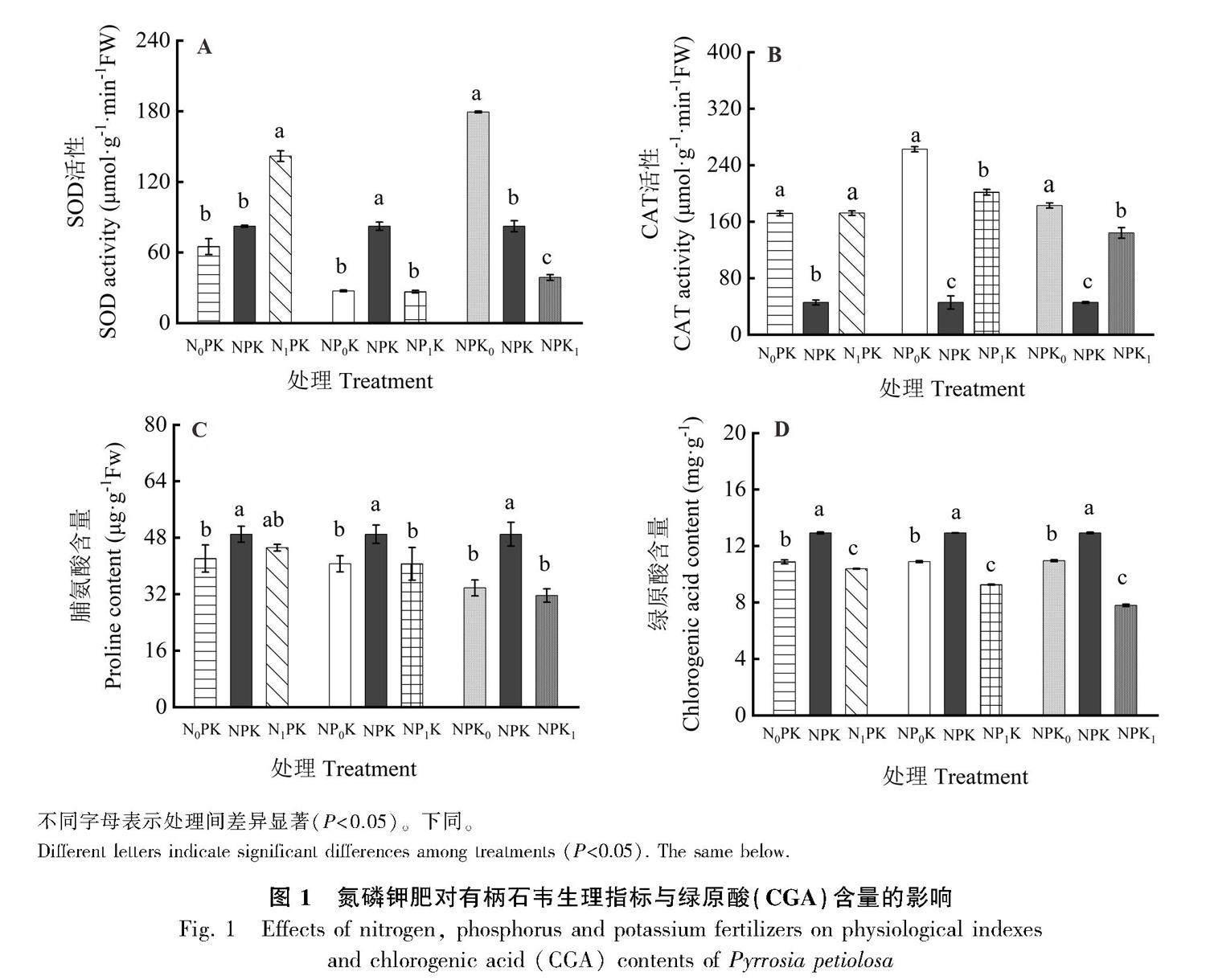
2.1.1 不同氮浓度对有柄石韦抗性生理的影响 由图1可知,高氮处理的SOD活性是低氮处理的2.18倍,正常施氮与低氮处理的SOD活性无显著性差异(图1:A)。正常施氮的CAT活性最低(图1:B),与低氮和高氮处理达到显著水平。低氮处理下脯氨酸含量最低,为42.09 μg·g-1,与正常氮处理达到显著水平,与高氮处理无显著差异。随着施氮量增加,脯氨酸含量呈现先升高后下降的趋势(图1:C)
2.1.2 不同磷浓度对有柄石韦抗性生理的影响 在磷处理试验中,正常施磷的SOD活性最高,而高磷处理与低磷处理的SOD活性无显著性差异(图1:A)。不同磷浓度下有柄石韦的CAT活性变化趋势与SOD活性变化趋势相反,正常磷浓度时CAT活性最低,低磷处理和高磷处理分别较正常磷升高了82.64%和77.41%,均达到显著水平(图1:B)。正常施磷时脯氨酸含量最高,高磷处理与低磷处理无显著性差异(图1:C)。
2.1.3 不同钾浓度对有柄石韦抗性生理的影响 不同钾浓度处理间有柄石韦SOD活性存在显著性差异,低钾处理SOD活性最高,高钾处理SOD活性最低(图1:A)。CAT活性在正常施钾肥情况下最低,低钾处理下CAT活性显著高于正常施钾和高钾处理。钾浓度对有柄石韦的脯氨酸含量的影响与氮、磷处理的影响一致,即正常施钾显著高于低钾处理和高钾处理,低钾处理与高钾处理间无显著性差异(图1:C)。
2.2 氮磷钾肥对有柄石韦CGA含量及合成关键酶活性的影响
2.2.1 氮磷钾肥对有柄石韦CGA含量的影响 由图1可知,不同水平氮、磷和钾肥对有柄石韦CGA含量存在显著影响。正常施氮CGA含量最高,达12.92 mg·g-1,低氮和高氮处理分别较正常施氮降低了15.87%和19.66%,均达到了显著水平。高磷和高钾处理不利于有柄石韦的CGA积累,与正常施磷和正常施钾肥相比分别下降了28.33%和39.71%,钾浓度对有柄石韦CGA含量影响更显著。
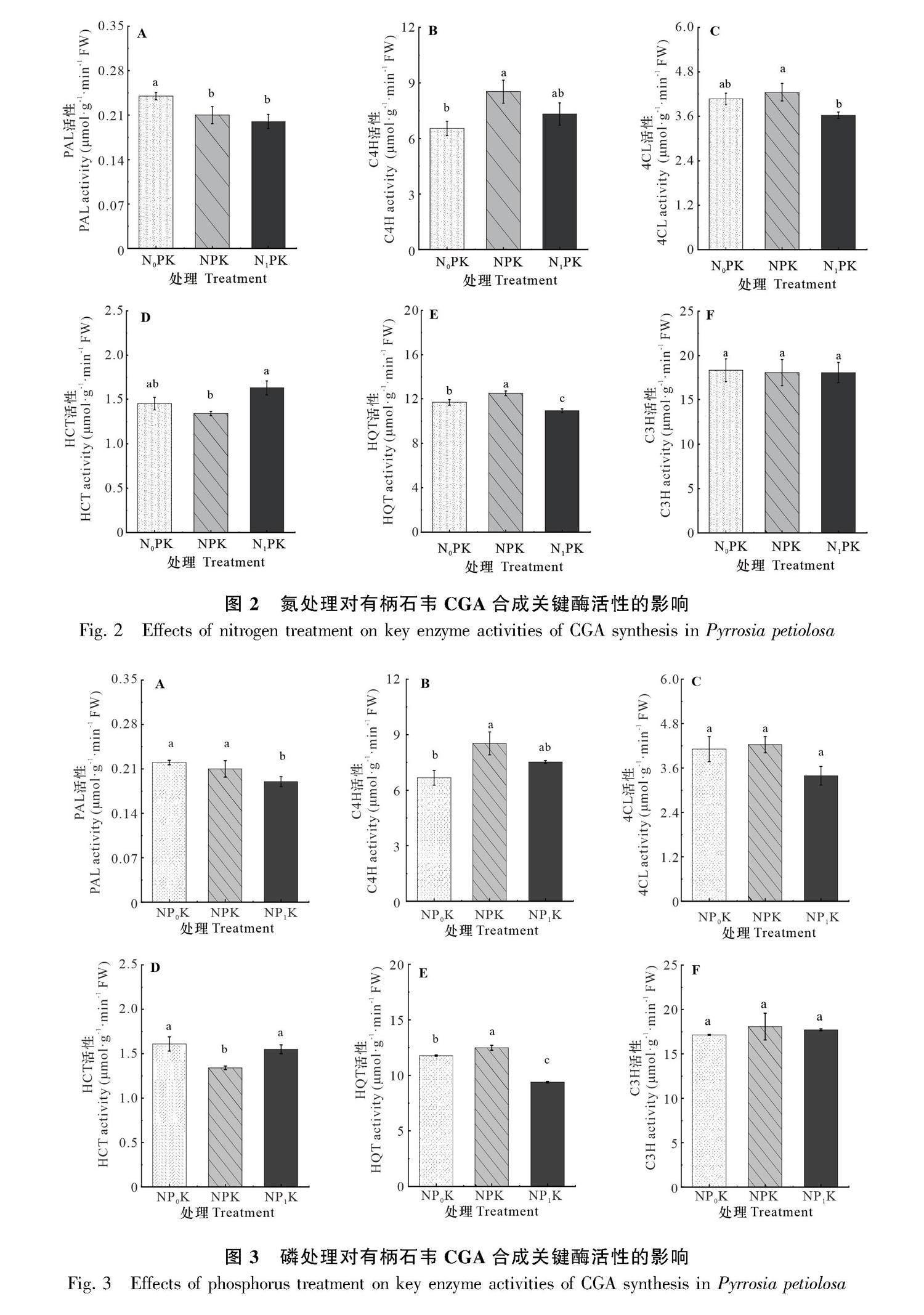
2.2.2 氮处理对有柄石韦CGA合成关键酶活性的影响 低氮处理的PAL活性最高,而高氮处理与正常施氮的PAL活性无显著变化(图2:A)。低氮处理的C4H活性较正常施氮降低了23.39%,达到显著水平(图2:B)。高氮处理的4CL活性较正常施氮相比下降了13.23%,低氮处理和正常施氮间无显著性差异(图2:C)。高氮处理的HCT活性比正常施氮显著升高了21.05%,而正常施氮的HCT活性最低(图2:D)。高氮处理下HQT活性与正常施氮相比降低了12.6%,均达到显著水平(图2:E)。氮浓度对C3H活性无显著影响(图2:F)。
2.2.3 磷处理对有柄石韦CGA合成关键酶活性的影响 高磷处理下PAL活性最低,较正常施磷处理组降低了12.85%,而低磷处理较正常施磷PAL活性无显著变化(图3:A)。磷处理对C4H活性的影响与氮处理相似,呈先升高后降低的趋势(图3:B)。低磷处理4CL活性与正常施磷相比无显著变化,而高磷处理4CL活性较正常施磷降低了14.39%(图3:C)。正常施磷的HCT活性最低,低磷和高磷处理较正常施磷相比分别升高了20.15%和15.67%(图3:D)。HQT活性随磷处理浓度先上升后下降,高磷处理较正常磷处理降低了24.83%(图3:E)。磷处理对C3H活性无显著影响(图3:F)。
2.2.4 钾处理对有柄石韦CGA合成关键酶活性的影响 钾浓度对有柄石韦PAL活性无显著影响(图4:A)。低钾处理和高钾处理的C4H活性较正常施钾分别降低了23.44%和31.72%,均达到显著水平(图4:B)。低钾处理与正常施钾的4CL活性无显著性差异,而高钾处理较正常施钾下降了22.40%(图4:C)。高钾处理较正常施钾HCT活性升高了28.94%(图4:D)。高钾处理的HQT活性较正常施钾相比降低了22.45%,而低钾处理与正常施钾无显著性差异(图4:E)。C3H活性在不同钾处理下无显著变化(图4:F)。
2.3 氮磷钾肥对有柄石韦CGA含量及其合成关键酶的相关性分析
不同浓度的氮磷钾肥对有柄石韦CGA合成途径中关键酶活性与CGA含量相关性分析如表1所示,上游关键酶4CL与有柄石韦CGA含量呈极显著正相关,下游关键酶HQT活性与CGA含量呈显著正相关,而下游关键酶HCT活性与CGA含量呈显著负相关。
3 讨论与结论
施氮量不足和过多时都会对植物产生不同的胁迫作用,造成抗氧化酶活性变化(刘寒等,2020),植物能通过促进抗氧化酶系统加强清除活性氧的能力以维持正常生长(Nadarajah, 2020)。本研究中,高浓度氮会对有柄石韦生长造成胁迫环境,而植株则通过提高SOD、CAT活性以适应环境,与羌活在氮处理中抗氧化酶活性变化的研究一致(张燕等,2018)。抗氧化酶系统协同作用才能消除体内有害的自由基,植物通过提高CAT活性应对高磷和低磷胁迫造成的活性氧损害;而SOD活性随着钾浓度增高降低,这与低钾胁迫下孔雀草抗氧化系统变化规律较为一致(王宏宇等,2020)。整体而言,低浓度和高浓度施肥会对有柄石韦生长造成胁迫,但CAT变化趋势与SOD变化不一致,SOD活性主要响应于有柄石韦的高氮和低钾处理,而CAT活性则对3种养分的低浓度和高浓度处理都有显著响应。脯氨酸在植物发育过程中发挥着重要作用,其合成与代谢与许多细胞途径有关,如光合作用、线粒体呼吸、氧化还原平衡、能量稳态等(Alvarez et al., 2022)。研究表明,脯氨酸也是植物获得能量的来源,通过介导NADP+/NADPH比率的变化影响氧化磷酸戊糖途径(OPPP)的碳通量,提供了赤藓糖-4-磷酸形式的前体,以在应激条件下合成苯丙素或次级代谢产物(Kishor et al., 2005)。这可能是本研究中脯氨酸含量变化与CGA含量变化规律较为相似的原因。
本研究表明,氮磷钾肥对有柄石韦CGA积累具有显著影响。Kovacik等(2020)研究表明连翘在高氮处理下CGA含量减少,而正常施氮更利于CGA积累,可能与高氮处理下CGA合成相关酶基因表达下调有关(张婷婷等,2022)。同样,低磷胁迫会使参与苯丙烷次生代谢物生物合成的PAL和4CL基因表达下调(Zhao et al., 2023),这可能是导致低磷处理下有柄石韦CGA含量显著下降的原因。Sun等(2023)研究表明,苹果叶片在缺钾处理下PAL、C4H和4CL基因表达上调,在高浓度钾处理状态下PAL基因表达下调,类黄酮通路在对不同钾处理的反应中起着重要作用。本研究中,高钾处理下CGA含量处于最低水平,与付立忠等(2019)高钾处理不利于三叶青酚类物质积累的研究结果一致。推测可能是植株中钾与钙、镁等阳离子之间存在着过高拮抗作用,过量施钾会降低植株的Ca/K、Mg/K,导致植株养分不平衡,从而影响植物的次生代谢过程(王千等,2012)。PAL、4CL、C4H、HCT、C3H和HQT作为植物体内CGA合成关键酶已经在金银花(张静茹等,2016)、桑叶(Zhao et al., 2019)等植物中得到验证,但在合成途径中所占的重要性不同。本研究中,有柄石韦在不同的肥料处理下CGA含量及其合成关键酶活性存在显著性差异。相关性分析表明,CGA含量与上游关键酶4CL和下游限速酶HQT呈显著正相关,而与HCT活性呈显著负相关。HCT 是HQT的上游酶,两者之间的底物与产物存在联系,有着反馈调节和负反馈调节。可见,4CL、HQT和HCT是氮磷钾3种主要养分影响有柄石韦CGA积累的关键酶,但其影响机制还有待进一步研究。
综上所述,氮磷钾肥对有柄石韦的SOD和CAT两种抗氧化酶活性有显著的影响,SOD活性主要响应高氮和低钾处理,而CAT活性则对3种养分的低浓度和高浓度处理均有显著响应。不同水平氮、磷和钾浓度对有柄石韦CGA含量存在显著影响,正常施肥的CGA含量最高,达12.92 mg·g-1,高钾施肥的CGA含量最低,为7.79 mg·g-1,钾肥对CGA含量影响最显著。HQT、4CL和HCT是造成不同施肥处理有柄石韦CGA含量差异的关键酶。
参考文献:
ALVAREZ ME, SAVOURE A, SZABADOS L, 2022. Proline metabolism as regulatory hub [J]. Trends Plant Sci, 27(1): 39-55.
CAO XX, KONG FL, YANG FT, et al., 2020. Effects of different nitrogen levels on the accumulation of saponins in American Ginseng [J]. J NE Agric Sci, 45(6): 78-81. [曹欣欣, 孔凡丽, 杨粉团, 等, 2020. 不同氮素水平对西洋参皂苷积累的影响 [J]. 东北农业科学, 45(6): 78-81.]
CLIFFORD MN, JAGANATH IB, LUDWIG IA, et al., 2017. Chlorogenic acids and the acyl-quinic acids: discovery, biosynthesis, bioavailability and bioactivity [J]. Nat Prod Rep, 34(12): 1391-1421.
FRITZ C, PALACIOS-ROJAS N, FEIL R,et al., 2006. Regulation of secondary metabolism by the carbon-nitrogen status in tobacco: nitrate inhibits large sectors of phenylpropanoid metabolism [J]. Plant J, 46(4): 533-548.
FU LZ, ZHAO LM, L HQ, et al., 2019. Effects of potassium on biomass, chemical composition of stems and leaves and antioxidant activity of Tetrastigma hemsleyanum [J]. J Chin Med Mat, 42(12): 2751-2754. [付立忠, 赵利梅, 吕惠卿, 等, 2019. 钾对三叶青生物量及其茎叶化学成分、抗氧化活性的影响 [J]. 中药材, 42(12): 2751-2754.]
GAO JF, 2006. Plant physiology experiment instruction [M]. Beijing: Higher Education Press: 142-211. [高俊凤, 2006. 植物生理学试验指导 [M]. 北京: 高等教育出版社: 142-211.]
HAN JP, LIANG ZS, SUN Q, et al., 2004. Study on the characteristic of assimilating nitrogenous phosphorous fertilizer and the accumulation disciplinarian of total tanshinons of Salvia miltiorrhiza [J]. Chin J Chin Mat Med, 29(3): 19-23. [韩建萍, 梁宗锁, 孙群, 等, 2004. 丹参根系氮、磷营养吸收及丹参酮累积规律研究 [J]. 中国中药杂志, 29(3): 19-23.]
KISHOR PBK, SANGAM S, AMRUTHA RN, et al., 2005. Regulation of proline biosynthesis, degradation, uptake and transport in higher plants: its implications in plant growth and abiotic stress tolerance [J]. Curr Sci, 88(3): 424-438.
KOVACIK J, DRESLE S, PETERKOVA V, et al., 2020. Nitrogen nutrition modulates oxidative stress and metabolite production in Hypericum perforatum [J]. Protoplasma, 257(2): 439-447.
LAI HB, MEI QX, FAN WC, 2010. A review on chemical constituents, pharmacological effects, and clinical applications of Pyrrosia petiolosa [J]. Chin Med Her, 7(21): 9-11. [赖海标, 梅全喜, 范文昌, 2010. 石韦的化学成分、药理作用和临床应用研究进展 [J]. 中国医药导报, 7(21): 9-11.]
LI HS, 2000. Plant physiological and biochemical principles and techniques [M]. Beijing: Higher Education Press: 164-168. [李合生, 2000. 植物生理生化实验原理和技术 [M]. 北京: 高等教育出版社: 164-168.]
LIU H, JIN SY, YANG LX, et al., 2020. Effect of nitrogen application on antioxidant enzyme system of Acanthopanax senticosus [J]. Chin J Exp Tradit Med Form, 26(4): 157-161. [刘寒, 金司阳, 杨立学, 等, 2020. 施氮量对刺五加幼苗抗氧化酶系统的影响 [J]. 中国实验方剂学杂志, 26(4): 157-161.]
NADARAJAH KK, 2020. ROS homeostasis in abiotic stress tolerance in plants [J]. Int J Mol Sci, 21(15): 5208.
National Pharmacopoeia Commission, 2020. Pharmacopoeia of the Peoples Republic of China: Part I [S]. Beijing: China Pharmaceutical Science and Technology Press: 91-92. [国家药典委员会, 2020. 中华人民共和国药典: 一部 [S]. 北京: 中国药科技出版社: 91-92.]
PAN LY, SUO CX, QIU YJ, et al., 2020. Study on UPLC specific chromatogram of Pyrrosia petiolosa (Christ) Ching [J]. J Guangdong Pharm Univ, 36(6): 784-790. [潘礼业, 索彩仙, 邱韵静, 等, 2020. 有柄石韦药材UPLC特征图谱研究 [J]. 广东药科大学学报, 36(6): 784-790.]
SUN T, ZHANG J, ZHANG Q, et al., 2023. Transcriptional and metabolic responses of apple to different potassium environments [J]. Front Plant Sci, 14: 1131708.
WANG HY, ZHANG YL, ZONG XC, 2020. Effect of low potassium stress on defensive enzyme activity of Tagetes patula [J]. Hubei Agric Sci, 59(5): 110-112. [王宏宇, 张彦丽, 宗宪春, 2020. 低钾胁迫对孔雀草防御性酶活性的影响 [J]. 湖北农业科学, 59(5): 110-112.]
WANG Q, YI YL, ZHANG SX, 2012. Effects of different potassium on phenolic metabolism of tomato seedlings [J]. J Plant Nutr Fertil, 18(3): 706-716. [王千, 依艳丽, 张淑香, 2012. 不同钾肥对番茄幼苗酚类物质代谢作用的影响 [J]. 植物营养与肥料学报, 18(3): 706-716.]
WANG Z, MA L, CHEN C, et al., 2023. Effects of phosphorus stress on the growth and secondary metabolism of Artemisia argyi [J]. J Plant Res. DOI: 10.1007/s10265-023-01479-z.
XUE Y, ZHU S, SCHULTZE-KRAFT R, et al., 2022. Dissection of crop metabolome responses to nitrogen, phosphorus, potassium, and other nutrient deficiencies [J]. Int J Mol Sci, 23(16): 9079.
ZHANG JR, WU ML, LI WD, et al., 2016. Regulation of honeysuckle HQT gene to chlorogenic acid biosynthesis in eukaryotic plant cells of Lonicerae Flos [J]. Chin Tradit Herb Drugs, 47(20): 3683-3687. [张静茹, 吴敏琳, 李卫东, 等, 2016. 金银花HQT基因在真核植物细胞中对绿原酸生物合成的调控 [J]. 中草药, 47(20): 3683-3687.]
ZHANG NJ, LI JJ, WANG CL, et al., 2022. Effects of the balanced fertilization of N, P and K on the yield, quality and related gene expression in Lonicera japonica [J]. J Gansu Agric Univ, 57(4): 88-95. [张宁洁, 李金娟, 王春霖, 等, 2022. N、P、K平衡施肥对金银花产量和品质及相关基因表达的影响 [J]. 甘肃农业大学学报, 57(4): 88-95.]
ZHANG TT, MENG LL, LIU XR, et al., 2022. Response of nitrogen metabolism of potato to low nitrogen stress and transcriptome analysis [J]. J NW A & F Univ (Nat Sci Ed), 50(8): 15-26. [张婷婷, 孟丽丽, 刘晓蕊, 等, 2022. 马铃薯氮代谢对低氮胁迫的响应及转录组分析 [J]. 西北农林科技大学学报(自然科学版), 50(8): 15-26.]
ZHANG Y, WANG HL, JIANG SY, et al., 2018. Effect of different nitrogen concentrations on growth and antioxidant enzymes activity in Notopterygium incisum seedling in cultivation matrix [J]. Chin J Exp Tradit Med Form, 24(7): 38-44. [张燕, 王红兰, 蒋舜媛, 等, 2018. 不同氮浓度对羌活幼苗生长及抗氧化酶系统的效应 [J]. 中国实验方剂学杂志, 24(7): 38-44.]
ZHAO L, WANG D, LIU J, et al., 2019. Transcriptomic analysis of key genes involved in chlorogenic acid biosynthetic pathway and characterization of MaHCT from Morus alba L. [J]. Protein Expr Purif, 156: 25-35.
ZHAO X, CHEN KK, WANG LT, et al., 2023. Transcriptome analysis provides insights into the response of Lotus corniculatus roots to low-phosphorus stress [J]. Front Plant Sci, 14: 1089380.
ZHAO YH, 2000. Biochemical experiment technology tutorial [M]. Guangzhou: South China University of Technology Press: 151-154. [赵亚华, 2000. 生物化学实验技术教程 [M]. 广州: 华南理工大学出版社: 151-154.]
(责任编辑 周翠鸣)
