外源赤霉素处理影响湖北梣种子萌发过程的生理机制
2024-06-25赵怡宁刘彦宏郑鹏丽周明芹
赵怡宁 刘彦宏 郑鹏丽 周明芹

收稿日期:2023-12-07
基金项目:国家自然科学基金资助项目(31200528)
作者简介:赵怡宁(1998-),女,湖南常德人,在读硕士研究生,研究方向为园林植物,(电话)19936731678(电子信箱)814601873@qq.com;通信作者,周明芹(1978-),女,湖北荆州人,教授,博士,主要从事园林植物种质资源的利用与创新、植物系统分类等方面研究,(电子信箱)500548@ yangtzeu.edu.cn。
赵怡宁,刘彦宏,郑鹏丽,等. 外源赤霉素处理影响湖北梣种子萌发过程的生理机制[J]. 湖北农业科学,2024,63(5):70-76,83.
摘要:以常温贮藏2年的湖北梣(Fraxinus hupehensis Chiú. Shang et Su)种子为研究对象,试验前去除种子胚根端被覆组织以露出胚根。以清水作对照,用150 mg/L赤霉素浸种24 h后,测定种子萌发过程中生理生化指标及内源激素含量的变化。结果表明,赤霉素浸种可以提高种子发芽率,赤霉素处理后种子发芽率为24.67%,对照为16.67%;萌发过程中,赤霉素处理后种子可溶性糖含量呈先上升再下降的变化趋势;可溶性蛋白质含量呈下降趋势,各时期均显著高于对照;超氧化物歧化酶(SOD)活性、过氧化物酶(POD)活性、过氧化氢酶(CAT)活性整体呈上升趋势;赤霉素处理后的种子内源茉莉酸甲酯(JA-Me)含量均显著低于对照,更有利于内源赤霉素(GA3)、玉米素核苷(ZR)含量的增加;生长素比脱落酸(IAA/ABA)与赤霉素比脱落酸(GA3/ABA)和玉米素核苷比脱落酸(ZR/ABA)分别在0.01和0.001水平上呈显著正相关;GA3/ABA与ZR/ABA在0.001水平上呈显著正相关。赤霉素浸种显著提高了湖北梣种子的发芽率,处理后的种子平衡保护酶活性系统较强,质膜受损延缓,抗逆能力较好。较低内源JA-Me含量和较高内源GA3、ZR有利于种子萌发的启动和休眠的解除。
关键词:湖北梣(Fraxinus hupehensis Chiú. Shang et Su); 种子; 生理生化; 萌发; 赤霉素
中图分类号:S792.41 文献标识码:A
文章编号:0439-8114(2024)05-0070-07
DOI:10.14088/j.cnki.issn0439-8114.2024.05.013 开放科学(资源服务)标识码(OSID):
Physiological mechanisms of exogenous gibberellin treatment affecting the seed germination process of Fraxinus hupehensis Chiú. Shang et Su
ZHAO Yi-ning, LIU Yan-hong, ZHENG Peng-li, ZHOU Ming-qin
(College of Horticulture and Gardening, Yangtze University, Jingzhou 434000, Hubei,China)
Abstract: The seeds of Fraxinus hupehensis stored at normal temperature for 2 years were taken as the research object. Before the experiment, the radicle end covering tissue was removed to expose the radicle. Taking water as a control, after soaking the seeds with 150 mg/L gibberellin solution for 24 h, the physiological and biochemical indexes and the contents of endogenous hormones in seeds during germination were measured. The results showed that gibberellin treatment could enhance seed germination rate, with a germination rate of 24.67% after gibberellin treatment compared to 16.67% in the control group. During germination, gibberellin-treated seeds exhibited a trend of initially increasing and then decreasing in soluble sugar content. Soluble protein content showed a decreasing trend but remained significantly higher than the control group at all stages. Superoxide dismutase (SOD), peroxidase (POD), and catalase (CAT) activities showed an increasing trend on the whole. The content of endogenous jasmonic acid methyl ester (JA-Me) in seeds after gibberellin treatment was significantly lower than that in the control, which favored an increase in endogenous gibberellin (GA3) and zeatin riboside (ZR) content. The ratios of gibberellin to abscisic acid (GA3/ABA) and zeatin riboside to abscisic acid (ZR/ABA) were significantly positively correlated with the ratio of indole-3-acetic acid to abscisic acid (IAA/ABA) at 0.01 and 0.001 levels, respectively, and GA3/ABA was significantly positively correlated with ZR/ABA at 0.001 level. Gibberellin immersion significantly increased the germination rate of Fraxinus hupehensis seeds, and the seeds treated with gibberellin exhibited a stronger balance of protective enzyme activity system, delayed membrane damage, and the resistance was better. The lower levels of endogenous JA-Me and the higher levels of endogenous GA3 and ZR favored the initiation of seed germination and the release of dormancy.
Key words: Fraxinus hupehensis Chiú. Shang et Su; seed; physiology and biochemistry; germination; gibberellin
湖北梣(Fraxinus hupehensis Chiú. Shang et Su)为木犀科白蜡属的落叶乔木,原生于湖北省大洪山余脉的京山市、钟祥市一带,属于湖北省特有珍稀新品种保护植物和国家Ⅱ级珍稀濒危植物[1,2]。其树姿丰盈,耐修剪,观赏价值高,抗逆性强,在园林建设中应用广泛[2,3]。孔令雷等[4]的研究表明湖北梣提取物有抗癌作用,具有药用价值。有研究发现湖北梣种胚被覆组织机械阻碍了种子萌发[5]。杨湘虹等[6]认为休眠可能是几种因素共同作用的结果,内源抑制物的存在与胚生理发育未完全是造成湖北梣种子休眠的主要原因。湖北梣种子深度休眠的特性限制了其栽培与应用,而关于其种子萌发的生理机制研究却鲜有报道。
赤霉素(GA3)作为一种植物激素,影响植物生长和发育的许多方面[7],并已被广泛研究。GA3能打破种子休眠、促进萌发[8]。罗富成等[9]发现用赤霉素对种子进行处理,可提高其发芽率,类似的结论在蒺藜苜蓿种子[10]和海枣种子[11]的研究中也有体现。Song等[12]研究发现外源GA3提高了湖北梣种子的萌发率、活力和吸水率,改变了其内源激素含量。植物内源激素对种子的生长发育具有不同的效应,其含量变化也客观反映了种子的代谢活力,不同激素之间的协同、相互作用,共同调控种子萌发[13-15]。GA3在打破种子休眠后所引起的内源激素变化能反映种子萌发的程度,是评价种子萌发的关键指标[16-19]。综上,有关外源GA3处理在湖北梣种子萌发过程中的变化研究尚为鲜见。
本研究通过施用外源赤霉素,测定种子不同时间的生理生化指标和内源激素变化,探究内源激素在种子萌发中的作用,借以初步研究抑制湖北梣种子萌发的内部因素,以期提出切实有效的休眠破除方法,为后续湖北梣种子内物质代谢、内源激素调控及信号传导等方面研究做铺垫,为其繁育及开发应用提供理论依据。
1 材料与方法
1.1 材料及其处理
湖北梣种子采集于湖北省宜昌市三峡植物园,将种子保留果翅,常温贮藏。去翅处理后,各处理选取1 000粒果粒饱满的种子用于发芽试验。试验开始前,将种子用高锰酸钾(0.2%)消毒后去除胚根端被覆组织,以150 mg/L赤霉素浸种24 h,用清水作对照。
1.2 方法
用电导法测定种子的质膜透性变化,用蒽酮比色法测定种子可溶性糖含量[20],采用考马斯亮蓝法[21]测定可溶性蛋白质含量,采用氮蓝四唑法[22]测定超氧化物歧化酶(SOD)活性,采用愈创木酚显色法测定[19]过氧化物酶(POD)活性,采用紫外吸收法[23]测定过氧化氢酶(CAT)活性,用电导仪测定浸泡液和对照溶液的电导率。
取处理后的种子50粒均匀散布于铺有2层中速定性滤纸、直径9 cm的玻璃培养皿中,置于4 ℃冰箱中低温层积处理,每处理20个重复,随机选取3个重复标记为萌发试验处理,剩下17个重复用于内源激素含量测定。每3 d洒水1次,保证培养皿中滤纸处于湿润状态,每30 d取样1次,直至试验结束。
取萌发试验的0、30、60 d种子各1 g研磨成粉末,加入40 mL 80%冰甲醇,4 ℃冰箱下浸提8 h以上,离心过滤,取上清液,用旋转蒸发仪将提取液浓缩至原有体积的1/3。加入0.5 mol/L Na2HPO4,将pH调至8.00,用石油醚萃取3次后加水、交联聚乙烯吡咯烷酮(PVPP)置于摇床上振荡30 min,过滤后加入2 mol/L柠檬酸调pH至3.00,乙酸乙酯萃取3次,酯相于真空浓缩仪中蒸干,残留物用3 mL pH=3.00的Na2HPO4溶解。将收集液过C18固相萃取小柱进一步纯化,并将收集的洗脱液于旋转蒸发仪上蒸干,用色谱纯甲醇溶解残留物于2 mL离心管中,定容至1 mL,过0.45 μmL微孔滤膜待测。测定内源激素有茉莉酸甲酯(JA-Me)、赤霉素(GA3)、玉米素核苷(ZR)、生长素(IAA)、芸苔素(BR)、脱落酸(ABA)。
1.3 数据分析
采用Excel软件计算数据,利用IBM SPSS Statistics 26、GraphPad Prism软件和R语言进行显著性及相关性分析并制图。
2 结果与分析
2.1 湖北梣种子萌发过程中的生理生化分析
2.1.1 湖北梣种子萌发过程中发芽率和电导率的变化 如图1A所示,外源赤霉素处理能够显著提高湖北梣种子的发芽率。种子经150 mg/L赤霉素溶液浸泡24 h后,发芽率为24.67%;与清水对照(发芽率为16.67%)相比,其发芽率显著提高了8个百分点。电导率反映种子活力的高低,如图1B所示,0~30 d,湖北梣种子的电导率均呈较低水平,赤霉素浸种后种子电导率均低于对照,随着时间的增加,电导率均呈增长趋势,60 d电导率均较0 d显著增加,分别为27.16 μS/(cm·g)和31.76 μS/(cm·g),赤霉素浸种处理与对照相比在各处理时间内均呈较低水平。
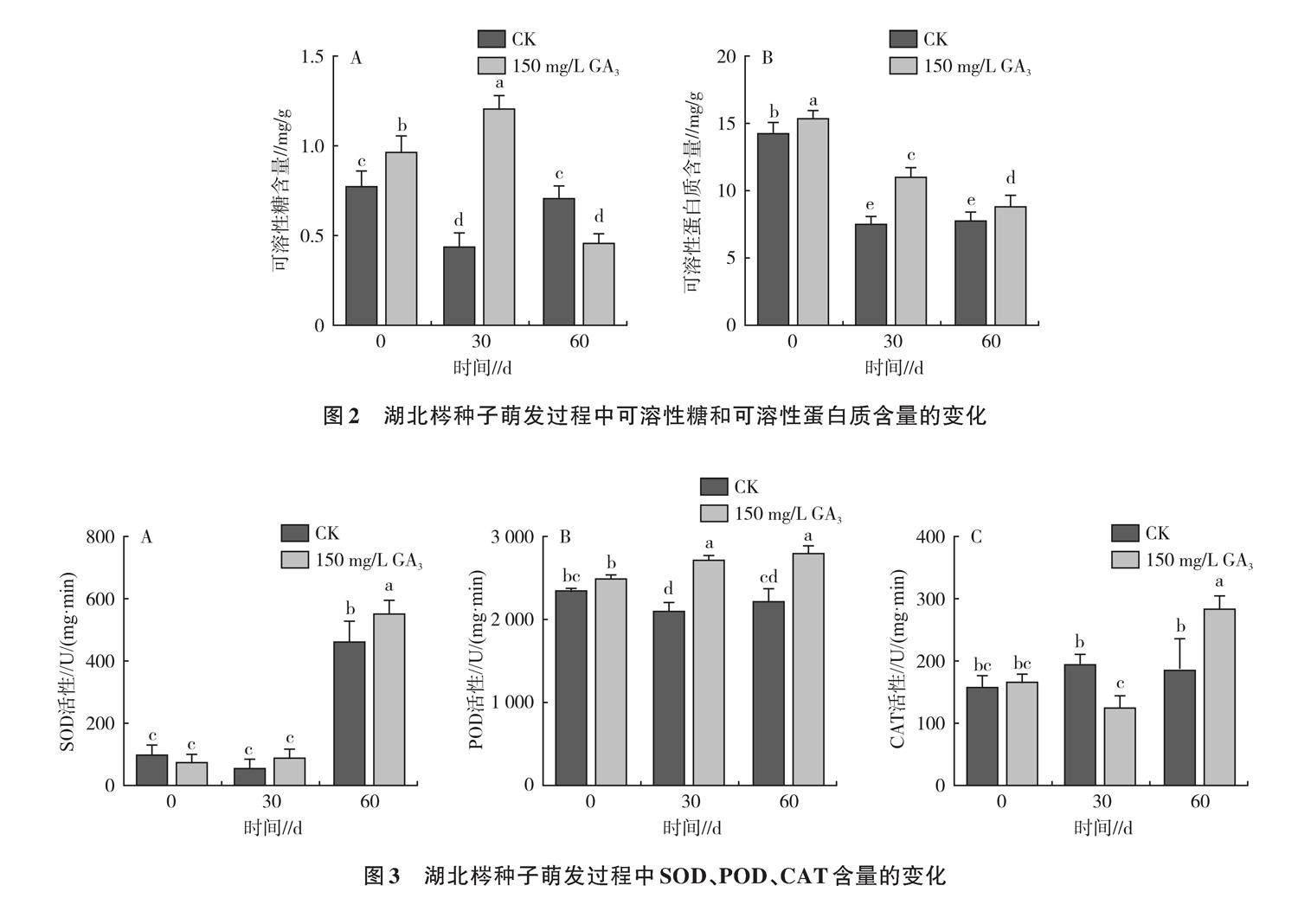
2.1.2 湖北梣种子萌发过程中可溶性糖和可溶性蛋白质含量的变化 可溶性糖含量能反映种子生理代谢过程中能量供应情况,如图2A所示,赤霉素处理的种子可溶性糖含量呈先升后降的趋势,与对照之间差异显著。试验30 d可溶性糖含量达到最大值,为1.21 mg/g,相比0 d增加了24.74%,60 d呈最低值,为0.46 mg/g,较0 d降低了52.58%。较低的可溶性蛋白质含量意味着种子内部代谢活动更为旺盛,如图2B所示,对照和赤霉素处理可溶性蛋白质含量的变化均呈下降趋势,0 d呈最大值(分别为14.81、15.36 mg/g),分别在30 d和60 d呈最低值(分别为7.53、8.87 mg/g),赤霉素处理可溶性蛋白质含量均显著高于对照。
2.1.3 湖北梣种子萌发过程中SOD、POD、CAT活性的变化 SOD维持活性氧代谢系统的平衡,如图3A所示,萌发0 d,各处理SOD处于较低水平;30 d时,对照和赤霉素处理SOD活性分别为53.47、88.42 U/(g·min);随后SOD活性显著上升,60 d时各处理SOD活性达到最高,赤霉素处理SOD活性显著高于对照。赤霉素处理后POD活性比SOD活性表现更加敏感,POD是种子贮藏过程中的重要保护酶,如图3B所示,赤霉素处理POD活性呈上升趋势,萌发30、60 d时POD活性显著高于对照。CAT可清除逆境下产生的过氧化氢,减弱植物所受的损伤,CAT活性的大小反映了细胞代谢的强度,如图3C所示,随着萌发时间的延长,赤霉素处理CAT活性随着萌发时间的延长呈先降低后升高的趋势,60 d时其活性为283.37 U/(g·min),较0 d增加了69.68%,与对照之间差异显著。
2.2 湖北梣种子萌发过程中内源激素含量的动态变化
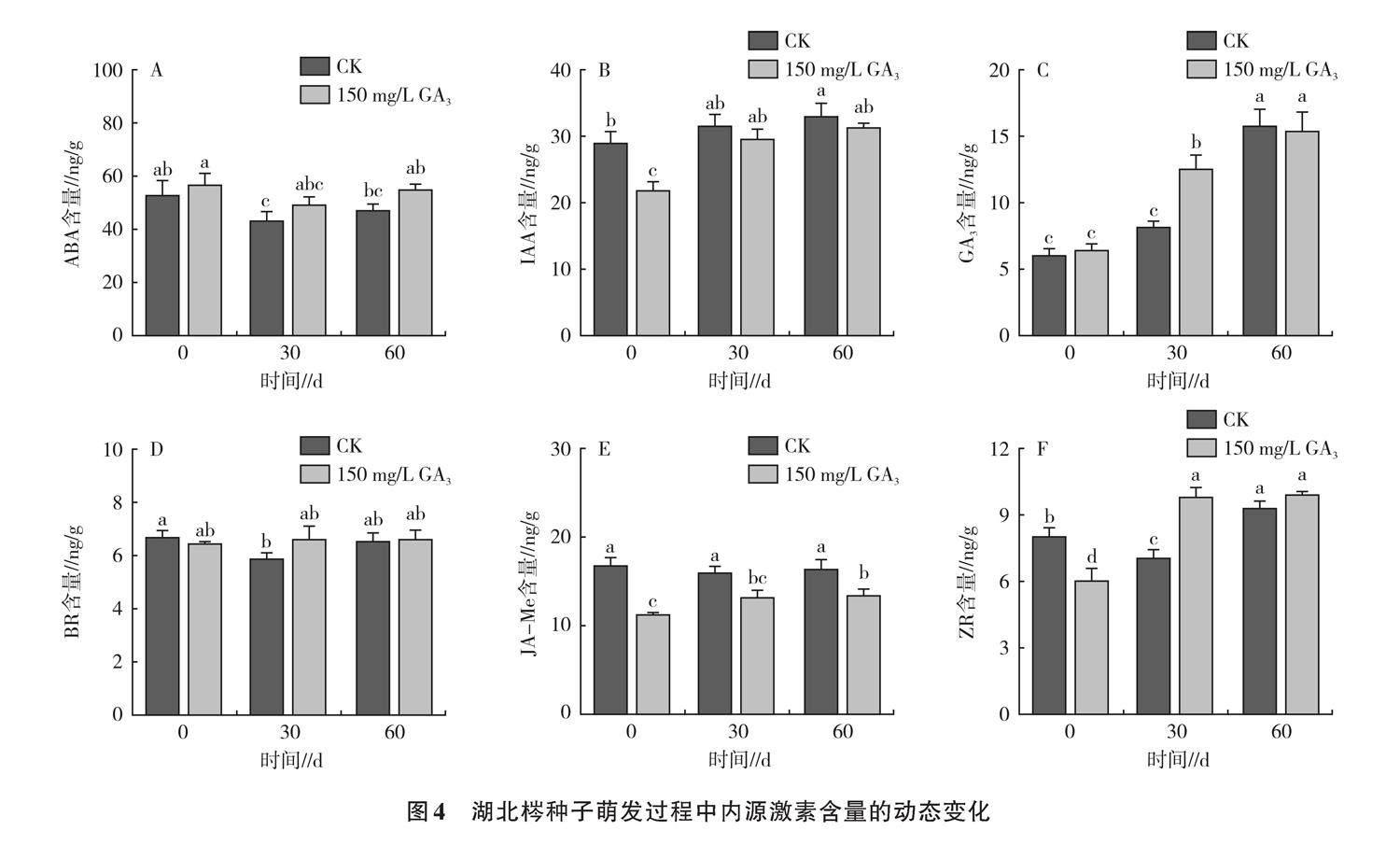
植物内源激素对植物的生理影响不是孤立发生的,二者之间存在复杂的关系网络[24]。如图4A所示,湖北梣种子内源ABA含量随着萌发时间的推移,呈先减后增的趋势,赤霉素处理ABA含量高于对照,在30 d时呈最低值(49.40 ng/g),60 d时赤霉素处理与对照ABA含量分别为55.32、47.24 ng/g,较0 d分别降低了2.76%和10.92%。表明试验前期种子ABA合成受到抑制,一定程度上利于破除种子休眠,而试验后期ABA含量的增加可能导致种子贮存合成物质增加,种子萌发受阻。此外,对内源激素进行相关性分析(图5)得出,ABA与JA-Me和IAA分别在0.01和0.001水平上呈显著负相关,相关系数(r)分别为-0.42和-0.48。
各处理种子IAA含量呈不断上升趋势,且赤霉素处理含量在每个试验阶段均低于对照(图4B)。0 d时赤霉素处理IAA含量显著低于对照,60 d时赤霉素处理和对照的IAA含量分别较0 d显著增加了42.67%、13.67%,这表明随着时间的推移,种子中的IAA含量在赤霉素处理下增加更为显著,但较高含量的IAA可能对解除种子休眠起到了负面影响。从图5可以看出,IAA与JA-Me在0.01水平上呈显著正相关(r=0.69),与IAA/ABA在0.001水平上呈显著正相关(r=0.87),与发芽率在0.01水平上呈显著负相关(r=-0.48)。
如图4C所示,各处理随着萌发时间的增加,GA3含量不断增加。30 d时,赤霉素浸种后种子GA3含量(12.61 ng/g)显著高于对照(8.27 ng/g);60 d时,GA3含量在各处理间无显著差异。说明外源赤霉素浸种可增加种子萌发过程中内源GA3含量,促进种子萌发。GA3与GA3/ABA、ZR/ABA分别在0.001和0.01水平上呈显著正相关(图5)。
芸苔素(BR)作为世界第六大激素,研究表明,其兼具赤霉素、细胞分裂素和生长素的综合功效,在平衡其他植物激素的同时还可以打破休眠、促进种子发芽[25]。赤霉素处理BR呈上升趋势,对照呈先下降后上升的趋势(图4D)。表明采用赤霉素浸种较采用清水浸种更有利于增加种子在萌发过程中的BR含量。BR与IAA/ABA在0.05水平上呈显著负相关(r=-0.45),与发芽率在0.05水平上呈显著正相关(r=0.27)(图5)。
如图4E所示,0 d时,赤霉素处理JA-Me含量显著低于对照32.6%,表明采用150 mg/g赤霉素浸种后种子质量更佳,具有较强抗逆性,而采用清水处理后种子损伤更大,品质更差,种子抗逆反应加强,JA-Me含量更高。各处理JA-Me含量随着试验时间推进不断上升,赤霉素处理试验各阶段JA-Me含量均显著低于对照,这可能是种子抗逆性逐渐降低,种子增加JA-Me含量来增强保护酶活性的原因,是种子对逆境做出的反应,赤霉素浸种后的种子更适应萌发环境。从图5可以看出,JA-Me与种子发芽率在0.001水平上呈显著负相关(r=0.90)。
玉米素核苷属于细胞分裂素,具有促进细胞分裂、抑制器官老化、诱导花芽分化、打破种子休眠的作用。如图4F所示,0 d时,采用赤霉素浸种种子的ZR含量低于采用清水的处理,可能是由于在试验前清水浸种使种子内部生理反应更为活跃。30 d时,赤霉素处理种子的ZR含量显著高于对照,较0 d增加62.68%,表明赤霉素处理更有利于促进萌发过程中ZR含量的增加。从图5可以看出,ZR与GA3、GA3/ABA、ZR/ABA均在0.001水平上呈显著正相关,r分别为0.81、0.78、0.87。
2.3 内源激素含量比例的变化
种子的萌发不仅与植物内源激素的绝对含量有关,还与各类激素之间的相互调节有关,特别是促进生长的激素与抑制生长的激素之间的比例及平衡有关[26]。本研究将湖北梣种子的IAA、GA3、BR、JA-Me、ZR测定值与ABA含量相比较,如图6所示,随着试验时间的推进,对照GA3/ABA、BR/ABA、ZR/ABA呈逐渐上升的趋势,IAA/ABA、JA-Me/ABA呈倒“V”形变化;赤霉素处理IAA/ABA、ZR/ABA、BR/ABA、JA-Me/ABA呈倒“V”形变化,GA3/ABA呈逐渐上升的趋势。各处理的ZR/ABA与GA3/ABA及IAA/ABA与JA-Me/ABA变化趋势基本相似,其比值在整个试验期间以GA3/ABA变化最为明显。从图5可以看出,IAA/ABA与GA3/ABA、ZR/ABA分别在0.01和0.001水平上呈显著正相关,相关系数分别为0.59、0.72;GA3/ABA与ZR/ABA在0.001水平上呈显著正相关(r=0.82)。
3 讨论与小结
湖北梣种子的萌发受到多重因素的综合影响,外源赤霉素处理湖北梣种子可以显著提高发芽率。种子萌发是物质和能量转化的过程,膜系统调节着各种物质的运输及各种酶的活性[27,28]。种子的抗逆性使得60 d可溶性蛋白质含量低于0 d,随着萌发时间的推进,细胞膜功能受损,种子通过消耗内部渗透物质维持其生理机能和萌发,致使60 d时各处理种子渗透调节物质含量低于0 d,此结果与魏朝阳[29]、杨万霞[30]的研究结论一致。SOD、POD、CAT能清除逆境条件下种子产生的活性氧,保护膜质系统[31]。在种子萌发的过程中,SOD活性表现出先下降后上升的趋势。在萌发初期(0 d),各处理组的SOD活性都较低。随着时间的推移,SOD活性逐渐上升,且在60 d时达到最高水平。赤霉素处理组的SOD活性在萌发30 d和60 d时均高于对照组。赤霉素处理后,种子中POD活性呈持续上升的趋势。在30 d和60 d时,赤霉素处理组的POD活性显著高于对照组,表明赤霉素处理能够显著提高种子中POD的活性,降低过氧化物对种子细胞膜的伤害,从而增强种子对逆境的抵抗能力,保持种子活性[32]。赤霉素处理后,种子中CAT活性的变化呈先降低后上升的趋势。在60 d时,赤霉素处理组的CAT活性显著高于对照组,表明赤霉素处理能够促进CAT活性的提高,这可能与种子萌发过程中细胞代谢的强度和逆境应对能力有关。综上所述,赤霉素处理对湖北梣种子萌发过程中SOD、POD和CAT活性产生了显著影响,表明赤霉素处理能够调节这些抗氧化酶的活性,从而有效降低种子在萌发过程中产生的活性氧,促进种子萌芽,与何秀燕等[32]在油樟中的研究相一致。
内源激素对种子萌发调控是一个复杂的生理生化过程[33-35],是各激素间的相互平衡、制约、协同[36]。Nambara等[37]的研究表明赤霉素浸种能增加种子抗逆性,增强保护酶活性,降低萌发内源抑制物含量,加快种子的萌发。本研究中,试验前赤霉素浸种的种子内源JA-Me含量显著低于对照,萌发过程中,IAA、ZR、BR含量增加幅度显著高于对照,对萌发抑制物的拮抗作用使种子内部生理生化反应保持活跃状态,为种子萌发做充分准备。IAA参与种子休眠的调控,促进GA3的合成[38],但高含量的IAA会抑制种子萌发。种子萌发时较高的IAA含量能提高代谢水平[39],但也使种子活力加速衰弱,类似研究在杨立学等[40]对紫椴的研究中也有所报道。Li等[36]对水稻种子的研究表明,BR参与反馈调节并抑制ABA产生,其含量与种子营养含量成正比,能提高发芽率和发芽势,缩短萌发时间。试验初期IAA、ZR、GA3与ABA较低的比值说明种子内较高的ABA可能是种子休眠的主要原因,GA3/ABA变化最为显著,综合表明IAA、ZR、GA3在调节种子萌发过程中存在一定的促进作用,可能是通过GA3与ABA的拮抗作用或各激素间互相拮抗与促进来调控。ABA不是惟一控制种子休眠与萌发的内源激素[41],ABA与GA3的关系可能为拮抗或者与两者绝对含量升降无关,种子萌发时ABA含量较高,而试验过程中ABA增长幅度显著低于GA3,减轻了对GA3的抑制作用,缩短了种子萌发时间。苏海兰等[42]的研究表明出苗前ABA含量持续上升,可能是为了阻止胚的过早萌发和淀粉萌发物的水解,该结果在浦梅等[43]的研究中得到了证实。与“三因子学说”一致,本研究中,较高的GA3与ABA比值水平表明,ABA并未影响GA3的合成或分解,萌发中ZR含量的增加可能解除了ABA对GA3的抑制作用,类似发现在矮金莲花种子[44]中得以体现。赤霉素浸种后,可能是通过改变种子透性,提高种子的呼吸作用,促使种子与外界气体交换,以解决生理后熟现象,同时调节种子内部萌发抑制物和促进物的相对水平,类似研究在二乔玉兰种子上得以运用[45]。
综上所述,用赤霉素浸种可以显著提高湖北梣种子发芽率,降低种子萌发过程中电导率;可溶性蛋白质含量呈下降趋势,各时期均显著高于对照;SOD活性、POD活性、CAT活性呈上升趋势,60 d时均显著高于对照,表明处理后的种子平衡保护酶活性系统较强,质膜受损延缓,抗逆能力较好。种子内源JA-Me含量均显著低于对照,更有利于内源GA3、ZR含量的增加, 较高的JA-Me含量对种子萌发的启动和休眠的解除产生了抑制作用,BR、ZR、GA3含量的增加对萌发的启动具有一定的正调控作用。相关性分析表明,IAA/ABA与GA3/ABA和ZR/ABA分别在0.01和0.001水平上呈显著正相关;GA3/ABA与ZR/ABA在0.001水平上呈显著正相关,IAA、ZR、GA3在调节种子萌发过程中存在一定的促进作用,可能是GA3与ABA的拮抗作用或各激素间互相拮抗与促进共同调节控制。
参考文献:
[1] 张 建.对节白蜡研究概况及展望[J].湖北林业科技,2015, 44(2):27-29.
[2] 杨艺涛,袁龙义,向君妍.乡土树种对节白蜡在城市景观空间设计中的应用[J].世界林业研究,2023,36(3): 147.
[3] 郑鹏丽,程建如,周明芹.濒危植物对节白蜡遗传多样性的cpSSR分析[J].西南农业学报,2020,33(7): 1399-1403.
[4] 孔令雷,胡金凤,陈乃宏.香豆素类化合物药理和毒理作用的研究进展[J].中国药理学通报,2012, 28(2):165-168.
[5] 刘雅琦,张 建,李 刚,等.对节白蜡种子发芽特性的研究[J].种子,2016,35(10):27-31.
[6] 杨湘虹,赵荣秋,刘乐承.对节白蜡的应用与繁殖研究进展[J].长江大学学报(自然科学版),2016,13 (27):4-7.
[7] GUAN Y R,XUE J Q,XUE Y Q,et al. Effect of exogenous GA3 on flowering quality, endogenous hormones, and hormone- and flowering-associated gene expression in forcing-cultured tree peony (Paeonia suffruticosa)[J]. Journal of integrative agriculture,2019, 18(6):1295-1311.
[8] MA H Y,ZHAO D D,NING Q R,et al. A multi-year beneficial effect of seed priming with gibberellic Acid-3 (GA3) on plant growth and production in a perennial grass, Leymus chinensis[J].Scientific reports,2018,8(1):13214.
[9] 罗富成,郭轶敏,彭 健,等.外源激素对纳罗克非洲狗尾草种子休眠的破除效果[J].草业科学,2015, 32(3):406-412.
[10] 刘文瑜,魏小红,许可成,等.蒺藜苜蓿种子休眠机制及其破除方法研究[J].草地学报,2015,23(2):358-365.
[11] DOAIGEY A R,AL-WHAAIBI M H,SIDDIQQUI M H,et al. Effect of GA3 and 2,4-D foliar application on the anatomy of date palm (Phoenix dactylifera L.) seedling leaf[J].Saudi journal of biological sciences,2013,20(2):141-7.
[12] SONG Q,CHENG S,CHEN Z,et al. Comparative transcriptome analysis revealing the potential mechanism of seed germination stimulated by exogenous gibberellin in Fraxinus hupehensis[J].BMC Plant Biol,2019,19(1):199.
[13] FENG R W,LEI L,SU J M,et al. Toxicity of different forms of antimony to rice plant: Effects on root exudates, cell wall components, endogenous hormones and antioxidant system[J].Science of the total environment,2020,771:134589.
[14] QU Z M,QI X C,LIU Y L,et al. Interactive effect of irrigation and polymer-coated potassium chloride on tomato production in a greenhouse[J].Agricultural water management,2020,235:106149.
[15] ZHAO X H,YU H Q,WEN J,et al. Response of root morphology, physiology and endogenous hormones in maize (Zea mays L.) to potassium deficiency[J].Journal of integrative agriculture,2016,15(4):785-794.
[16] JIANG H K,SHUI Z W,XU L,et al. Gibberellins modulate shade-induced soybean hypocotyl elongation downstream of the mutual promotion of auxin and brassinosteroids[J].Plant physiology and biochemistry,2020,150:209-221.
[17] SONG S W,LEI Y L,HUANG X M,et al. Crosstalk of cold and gibberellin effects on bolting and flowering in flowering Chinese cabbage[J].Journal of integrative agriculture,2019,18(5):992-1000.
[18] YANG L,LIU D C,HU W,et al. Fruit characteristics and seed anatomy of ‘Majia pomelo pollinated with cobalt-60?gamma-ray-irradiated pollen[J].Scientia horticulturae,2020,267(1):109335.
[19] CHEN Y,SONG X H,GUO Y Y,et al. Stalk length should be considered for storage quality of broccoli heads based on the investigation of endogenous hormones metabolism[J].Scientia horticulturae,2020,267:109338.
[20] 陈爱葵,韩瑞宏,李东洋,等.植物叶片相对电导率测定方法比较研究[J].广东教育学院学报,2010,30(5): 88-91.
[21] CHENG Y F,LIU Y L,WEI H M,et al. Quantitation of low concentrations of polysorbates 80 in protein formulations by Coomassie brilliant blue[J].Analytical biochemistry,2019,573:67-72.
[22] CHENG C W,CHEN L Y,CHOU C W,et al. Investigations of riboflavin photolysis via coloured light in the nitro blue tetrazolium assay for superoxide dismutase activity[J].Journal of photochemistry and photobiology B: Biology,2015,148:262-267.
[23] 李合生.植物生理生化试验原理和技术[M].北京:高等教育出版社, 2000.
[24] 李冬林,张亚楠,金雅琴,等.毛梾种子低温层积过程中内源激素的变化及其与发芽的关系[J].经济林研究,2014,32(3):36-41.
[25] 李振华.外源生长素和赤霉素信号调控烟草种子休眠与萌发的机理[D].北京:中国农业大学,2017.
[26] KHAN A A. Primary, preventive and permissive roles of hormones in plant systems[J].Botanical review,1975,41:391-420.
[27] LI Q F,ZHOU Y,XIONG M,et al. Gibberellin recovers seed germination in rice with impaired brassinosteroid signalling[J].Plant science,2020,293:110435.
[28] KIM S Y,WARPEHA K M,HUBER S C. The brassinosteroid receptor kinase, BRI1, plays a role in seed germination and the release of dormancy by cold stratification[J].Journal of plant physiology,2019,241:153031.
[29] 魏朝阳.冬青种子休眠机理及解除方法研究[D].南京:南京林业大学,2007.
[30] 杨万霞.青钱柳种子休眠原因及萌发生理的初步研究[D].南京:南京林业大学,2004.
[31] SHAH K,NAHAKPAM S. Heat exposure alters the expression of SOD, POD, APX and CAT isozymes and mitigates low cadmium toxicity in seedlings of sensitive and tolerant rice cultivars[J].Plant physiology and biochemistry,2012,57:106-113.
[32] 何秀燕,倪婷婷,刘孟婕,等.不同浓度赤霉素对油樟种子生理及萌芽的影响[J].四川林业科技,2023,44(3):110-115.
[33] DOAIGEY A R,AL-WHAIBI M H,SIDDIQUI M H,et al. Effect of GA3 and 2,4-D foliar application on the anatomy of date palm (Phoenix dactylifera L.) seedling leaf[J].Saudi journal of biological sciences,2013,20(2):141-147.
[34] WEN T,DONG L J,WANG L,et al. Changes in root architecture and endogenous hormone levels in two Malus rootstocks under alkali stress[J].Scientia horticulturae,2018,235:198-204.
[35] ZHANG Y J,LI A,LIU X Q,et al. Changes in the morphology of the bud meristem and the levels of endogenous hormones after low-temperature treatment of different Phalaenopsis cultivars[J].South African journal of botany,2019,125:499-504.
[36] LI Q,YANG A,ZHANG W H. Higher endogenous bioactive gibberellins and α-amylase activity confer greater tolerance of rice seed germination to saline-alkaline stress[J].Environmental and experimental botany, 2019,162:357-363.
[37] NAMBARA E,MARION-POLL A. ABA action and interactions in seeds[J].Trends in plant science,2003, 8:213-217.
[38] NOOR A,ZAIF K,Amjad M,et al. Synthetic auxins concentration and application time modulates seed yield and quality of carrot by altering the umbel order[J].Scientia horticulturae,2020,262:109066.
[39] 廖文燕.金钱松种子贮藏过程中的生理生化变化[D].南京:南京林业大学,2011.
[40] 杨立学,王海南,张 琳,等.低温层积过程中紫椴种子内源激素含量的变化[J].经济林研究,2012,30(2): 15-18.
[41] 李建军,连笑雅,王 兰.忍冬花蕾延迟开花与内源激素调控研究[J].园艺学报,2019,46(7):1399-1408.
[42] 苏海兰,周先治,李 希,等.七叶一枝花种子萌发不同阶段贮藏物质及淀粉酶活性变化[J].福建农业学报,2017,32(10):1145-1149.
[43] 浦 梅,孙永玉,高成杰,等.滇重楼种子内源激素含量与种胚长度和萌发的关系[J].林业科学研究,2016, 29(2):268-273.
[44] 张雁明.矮金莲花种子萌发的内源激素调控及其环境响应规律[D].兰州:兰州大学,2018.
[45] 沈 超. 二乔玉兰种子萌发特性及促萌因素解析[D].南京:南京林业大学, 2013.
