沉默减数分裂内切酶1(EME1)抑制肝癌细胞增殖的机制
2024-06-06陈椿王可欣贺梦雯李乐王春艳刘妍纪冬
陈椿 王可欣 贺梦雯 李乐 王春艳 刘妍 纪冬


摘要: 目的 探討减数分裂内切酶1 (EME1) 在肝癌组织中的表达及其对肝癌细胞生物学行为的影响。方法 筛选TCGA数据库肝癌样本中的差异表达基因。采用免疫组化和Western Blot分析EME1在肝癌组织中的表达丰度。通过短发夹RNA(shRNA) 构建慢病毒并感染BEL-7404细胞干扰EME1基因表达, 分为沉默组 (shEME1) 和对照组 (shCtrl)。通过实时荧光定量PCR法和Western Blot检测两组EME1 mRNA和蛋白表达水平, Celigo计数法及MTT活性检测细胞增殖率, 流式细胞术检测细胞周期, Caspase3/7活性检测细胞凋亡。两组间比较采用成组t检验。结果 TCGA结果显示EME1的mRNA表达水平在肝癌组织中是癌旁组织的18. 9倍 (114. 5±153. 0 vs 8. 0±7. 2, t=5. 00, P<0. 001); EME1的蛋白表达水平在肝癌组织中是癌旁组织的7. 0倍(免疫组化检测, 8. 4±2. 6 vs 1. 2±0. 4, t=7. 55, P<0. 001)和2. 5倍(Western Blot检测, 249. 0%±35. 5% vs 100. 0%±77. 8%, t=3. 02, P<0. 05)。慢病毒感染后, 相对于对照组, 沉默组EME1的mRNA表达水平下降了29. 9%(29. 9%± 0. 9% vs 100. 0%±3. 6%, t=32. 82, P<0. 001), 蛋白表达水平显著下降了35. 7% (35. 7%±14. 9% vs 100. 0%±28. 9%, t=3. 42, P<0. 05); 细胞计数下降了45. 1% (4 053±167 vs 8 988±477, t=16. 91, P<0. 001)、 细胞活性下降至66. 9% (0. 518±0. 046 vs 0. 774±0. 022, t=8. 74, P<0. 001) 及细胞克隆形成能力下降至29. 0% (75±6 vs 260±9, t=28. 92, P<0. 001)。与对照组比较, 沉默组G1期细胞 (49. 9% vs 44. 0%, t=8. 96, P<0. 001) 比例增多, G2/M期 (15. 9% vs 17. 9%, t=9. 13, P<0. 001) 与S期 (34. 2% vs 38. 1%,t=6. 91, P<0. 001)的细胞比例减少; Caspase3/7活性增强了1. 5倍(145. 8%±5. 9% vs 100. 0%±2. 3%, t=12. 50, P<0. 001)。结论 EME1在肝癌组织中高表达, 沉默EME1基因可抑制肝癌细胞增殖, 促进细胞凋亡。
关键词: 癌, 肝细胞; 减数分裂内切酶1; RNA, 小分子干扰; 细胞增殖
基金项目: 北京市自然科学基金面上项目 (7222173)
Silencing essential meiotic endonuclease 1 inhibits the proliferation of liver cancer cells: A study of related mechanisms
CHEN Chun1, WANG Kexin2, 3, HE Mengwen2, 4, LI Le5, WANG Chunyan2, LIU Yan5, JI Dong1, 2, 3, 4. (1. The Second Clinical Medical School of Southern Medical University, Guangzhou 510515, China; 2. Senior Department of Hepatology, The Fifth Medical Center of Chinese PLA General Hospital, Beijing 100039, China; 3. PLA 307 Medical College of Anhui Medical University, Hefei 230032, China;4. Peking University 302 Clinical Medical School, Beijing 100191, China;5. Senior Department of Infectious Diseases, The Fifth Medical Center of Chinese PLA General Hospital, Beijing 100039, China)
Corresponding authors: JI Dong, jidg302@126.com (ORCID: 0000-0001-8214-462X); LIU Yan, liuyan5360@163.com (ORCID: 0000-0003-1498-4314)
Abstract: Objective To investigate the expression of essential meiotic endonuclease 1 (EME1) in liver cancer tissue and its effect on the biological behavior of hepatoma cells. Methods The TCGA database was used to identify the differentially expressed genes between liver cancer tissue and paracancerous tissue. Immunohistochemistry and Western Blot were used to measure the expression abundance of EME1 in liver cancer tissue. A lentivirus was constructed by short hairpin RNA, and BEL-7404 cells were transfected with the lentivirus to interfere with the expression of the EME1 gene; the cells were divided into silencing group (shEME1 group) and control group (shCtrl group) . Quantitative real-time PCR and Western Blot were used to measure the mRNA and protein expression levels of EME1; Celigo Image Cytometer and MTT assay were used to measure cell proliferation rate; flow cytometry was used to observe cell cycle; Caspase 3/7 activity was used to measure cell apoptosis. The independent-samples t-test was used for comparison between two groups. Results TCGA results showed that the mRNA expression level of EME1 in liver cancer tissue was 18.9 times that in paracancerous tissue (t=5.00, P<0.001), and the protein expression level of EME1 in liver cancer tissue was 7.0 times (based on immunohistochemistry: 8.4±2.6 vs 1.2±0.4, t=7.55, P<0.001) or 2.5 times (based on Western Blot: 249.0%±35.5% vs 100.0%±77.8%, t=3.02, P<0.05) that in paracancerous tissue. After lentivirus infection, compared with the shCtrl group, the shEME1 group had an mRNA expression level of EME1 reduced by 29.9% (29.9%±0.9% vs 100.0%±3.6%,
t=32.82, P<0.001), a protein expression level of EME1 reduced by 35.7% (35.7%±14.9% vs 100.0%±28.9%, t=3.42, P<0.05), and a level of cell counting reduced by 45.1% (4 053±167 vs 8 988±477, t=16.91, P<0.001), as well as a level of cell activity reduced to 66.9% (0.518±0.046 vs 0.774±0.022, t=8.74, P<0.001) and a level of colony forming ability reduced to 29.0% (75±6 vs 260±9, t=28.92, P<0.001) . Compared with the shCtrl group, the shEME1 group had a significant increase in the proportion of cells in G1 phase (49.9% vs 44.0%, t=8.96, P<0.001) and significant reductions in the proportion of cells in G2/M phase (15.9% vs 17.9%, t=9.13, P<0.001) and S phase (34.2% vs 38.1%, t=6.91, P<0.001), while Caspase 3/7 activity was enhanced by 1.5 times (145.8%±5.9% vs 100.0%±2.3%,t=12.50, P<0.001) . Conclusion EME1 is highly expressed in liver cancer tissue, and silencing the EME1 gene can inhibit the proliferation of hepatoma cells and promote cell apoptosis.
Key words: Carcinoma, Hepatocellular; Essential Meiotic Endonuclease 1; RNA, Small Interfering; Cell Proliferation
Research funding: Beijing Municipal National Science Foundation (7222173)
肝細胞癌是全球癌症相关死亡的第三大常见原因,对于晚期肝癌患者, 靶向治疗的效果仍不能满足临床需求[1-5] , 寻找新的基因靶点可进一步提升疗效及安全性,成为目前肝癌治疗领域的研究热点。减数分裂内切酶1(essential meiotic endonuclease 1, EME1) 编码的蛋白质可与MUS81形成核酸内切酶复合物, 作用于特定的DNA底物, 包括Holiday连接、 3′ 末端翘翼结构以及复制叉结构, 在DNA链间交联、 DNA双链断裂后修复过程中对重组中间产物的分解发挥着关键作用[6] 。既往研究[7-9] 发现, EME1高表达是膀胱癌、 乳腺癌及肾透明细胞癌不良预后的高危因素, 但其在肝癌中的表达和作用尚未见报道。本研究从TCGA数据库入手, 通过生物信息学寻找肝癌组织中的高表达基因EME1, 并在肝癌组织/癌旁组织中进行验证, 通过基因沉默探讨EME1对肝癌细胞生物学行为的影响, 以期鉴定新的治疗靶点。
1 材料与方法
1. 1 材料 人肝癌细胞BEL-7404/7402、 Hep3B及人胚肾细胞293T均为本单位感染病研究所保存。慢病毒载体购自上海吉凯基因公司。鼠源EME1单克隆抗体购自美国Santa Cruz公司, HRP标记山羊抗小鼠的二抗检测试剂盒购自北京中杉金桥公司。HiScript Ⅲ All-in one RT、 SuperMix Perfect for qPCR 逆转录试剂和 Taq Pro Universal SYBR qPCR Master Mix购自南京诺唯赞公司。RPMI 1640和MEM培养基购自美国Corning公司, 胎牛血清 (FBS) 购自美国Geniala公司, 四甲基偶氮唑盐 (MTT)购自北京鼎国昌盛公司, Caspase-Glo? 3/7 Assay试剂盒购自美国Promega公司, 碘化丙啶 (PI) 购自吉至生化公司。
1. 2 TCGA数据库分析 下载TCGA数据库中肝细胞癌样本的mRNA seq表达谱, 利用R软件中的 “edge” 包筛选癌及癌旁组织中高表达基因。
1. 3 细胞培养 BEL-7404/7402细胞在含10%FBS的RPMI 1640细胞培养液、 Hep3B细胞在含10%FBS的MEM细胞培养液中37 ℃、 5% CO2条件下培养。细胞传代使用0. 25%胰蛋白酶消化, 按照1∶3比例传代。
1. 4 免疫组化染色 石蜡切片经脱蜡、 水化、 抗原修复后, FBS封闭后加入EME1一抗, 4 ℃孵育过夜, 加入二抗37 ℃孵育20 min, DAB显色, 苏木精复染, 中性树胶封片, 镜检评分。染色结果进行半定量分析, 以染色强度评分 [阴性为0分, 淡黄色 (弱阳性) 为1分, 棕黄色 (阳性) 为2分, 棕褐色 (强阳性) 为3分] 和阳性细胞范围评分[≤25% 为 1 分, 26%~50% 计为 2 分, 51%~75% 为3分, >75%为4分] 乘积作为最终结果。
1. 5 EME1沉默慢病毒载体的构建 EME1 短发夹RN(shRNA) 干扰序列为CTGAGAAGACAGGAAAGAA, 两端分别添加AgeI (A^CCGGT) 和EcoRI (G^AATTC) 酶切位点黏性末端, 并在正链3′ 端添加终止信号 (TTTTT), 反链5端添加终止信号互补序列 (AAAAA), 合成DNA Oligo。双酶切hU6-MCS-CMV-EGFP慢病毒载体使其线性化, 将DNA Oligo与载体质粒相连后转化感受态细胞, 之后PCR扩增及测序鉴定 (表1)。提取鉴定正确的质粒, 利用第二代慢病毒包装系统, 穿梭质粒GV115、 包装质粒psPAX2、 包膜质粒pMD2. G共转染293T细胞, 转染后48 进行病毒收获, 浓缩与纯化慢病毒。
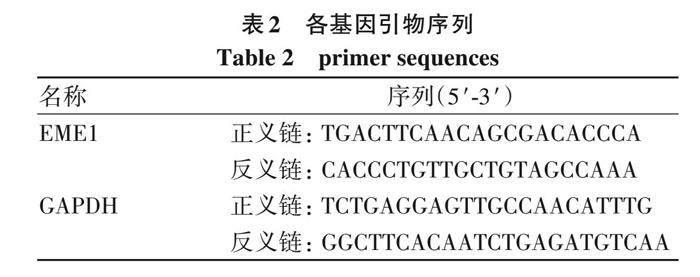
1. 6 EME1沉默慢病毒感染BEL-7404细胞 对数生长期的BEL-7404细胞接种于6孔板 (2×105个/孔), 培养至铺板量达30%后分别加入shRNA干扰表达慢病毒和阴性对照慢病毒, 感染12 h后更换完全培养基继续培养。感染72 h后, 荧光显微镜观察细胞感染效率。以GADPH为内参照, 进行实时荧光定量PCR (RT-qPCR) 检测 (表2) , 2?△△Ct法计算EME1相对表达量。
1. 7 Western Blot检测 制备样品, 每孔上样量为30 ?g,10%的SDS-PAGE电泳分离, 并转移到PVDF膜上, 室温封闭1 h, 一抗 (1∶800) 4 ℃孵育过夜, TBST洗膜。二抗(1∶5 000) 室温孵育1 h, TBST洗膜。化学发光法 (ECL法) 检测条带, 以GAPDH为内参照, 用Image J软件进行灰度值分析。
1. 8 Celigo与MTT检测 对数生长期细胞接种于96孔板 (2×103个/孔), 每天用Celigo检测读板。另设置一组进行MTT检测, 培养终止前4 h加入5 mg/mL的MTT溶液20 ?L, 继续培养4 h后吸取培养液, 每孔加入100 ?L二甲基亚砜, 振荡器振荡2 min, 检测490 nm处的吸光值。
1. 9 细胞克隆形成能力检测 对数生长期细胞接种于6孔板 (800个/孔), 持续培养10 d后使用4%多聚甲醛固定细胞30 min, PBS洗涤后加入500 ?L/孔吉姆萨染液(10 min), ddH2O洗涤细胞, 晾干后拍照并克隆计数。
1. 10 Caspase3/7检测 对数生长期细胞接种于96孔板(1×104个/孔), Caspase-Glo3/7缓冲液10 mL溶解Caspase-Glo3/7 冻干粉, 每孔加入 Caspase-Glo 反应液 100 ?L,500 r/min离心30 min, 室温孵育2 h后使用酶标仪测定信号强度。
1. 11 PI-FACS-周期检测 收集处于对数生长期的细胞, 用4 ℃预冷的D-Hanks洗涤细胞并沉淀, 用4 ℃预冷的75%乙醇固定细胞1 h。1 300 r/min离心5 min, 去固定液, 用4 ℃预冷的D-Hanks洗涤细胞沉淀1次。加入1 mL的细胞染色液重悬, 上机检测。
1. 12 统计学方法 采用R语言软件4. 2. 1进行统计分析, 每个实验均设置3个复孔, 数据以x ˉ±s表示, 两组间比较采用成组t检验。P<0. 05为差异有统计学意义。
2 结果
2. 1 人肝癌细胞中EME1 mRNA表达水平 TCGA数据库筛选出50对肝癌样本, EME1 mRNA在肝癌组织(114. 5±153. 0) 的表达量是癌旁组织 (8. 0±7. 2) 的18. 9倍 (t=5. 00, P<0. 001)(图1a)。RT-qPCR結果显示, 人肝癌细胞BEL-7404/7402和Hep3B中EME1 mRNA相对表达量 (ΔCT值) 分别为9. 25、 10. 32和10. 89, 均<12, 为高丰度表达 (图1b)。
2. 2 人肝癌细胞中EME1的蛋白表达水平 选取病理确诊的3例肝癌患者的手术切除癌/癌旁组织进行Western Blot分析, 结果显示肝癌组织EME1蛋白水平 (249. 0%± 35. 5%) 是癌旁组织 (100. 0%±77. 8%) 的2. 5倍 (t=3. 02,P<0. 05)(图2a、 b)。选取病理确诊的5例肝癌患者的手术切除肝癌组织/癌旁组织进行免疫组化分析, 结果显示肝癌组织EME1蛋白水平 (8. 4±2. 6) 是癌旁组织 (1. 2±0. 4) 的7. 0倍 (t=7. 55, P<0. 001)(图2c、 d)。
2. 3 EME1稳定沉默BEL-7404细胞系的构建 荧光倒置相差显微镜观察慢病毒感染72 h后的BEL-7404细胞,结果显示感染效率达到80%以上, 细胞状态正常 (图3)。RT-qPCR结果显示, 沉默组 (shEME1) EME1 mRNA表达水平 (29. 9%±0. 9%) 相较于阴性对照组 (shCtrl)(100. 0%± 3. 6%) 显著降低 (t=32. 82, P<0. 001)(图4a); Western Blot结果显示, shEME1组EME1蛋白表达水平 (35. 7%±14. 9%)相较于shCtrl组 (100. 0%±28. 9%) 显著降低 (t=3. 42, P<0. 05)(图4b), 表明EME1稳定沉默的肝癌细胞株BEL-7404构建成功。
2. 4 EME1沉默后BEL-7404细胞增殖能力的变化 Celigo观察培养1~5天的BEL-7404细胞, 结果显示随着培养时间的延长, 相对于对照组, 沉默组细胞数量显著下降, 培养5天时沉默组为4 053±167, 对照组为8 988±477, 细胞数量下降了45. 1% (t=16. 91, P<0. 001)(图5a、 b)。MTT检测培养1~5天的BEL-7404细胞在490 nm处的吸光值,相对于对照组, 沉默组细胞吸光值显著下降, 培养5天时沉默组为0. 518±0. 046, 对照组为0. 774±0. 022, 细胞活性下降了66. 9% (t=8. 74, P<0. 001)(5c)。培养10天后,沉默组克隆数 (75±6) 较对照组 (260±9) 显著减少, 细胞克隆形成能力下降了29. 0% (t=28. 92, P<0. 001)(图5d、e)。表明沉默EME1降低BEL-7404细胞的增殖能力。
2. 5 EME1沉默后BEL-7404细胞周期变化 流式细胞术检测慢病毒感染5天后的BEL-7404细胞周期, 沉默组的G1期细胞比例显著高于对照组 (49. 9%±0. 8% vs 44. 0%±0. 9%, t=8. 96, P<0. 001), G2/M期细胞比例显著低于对照组(15. 9%±0. 2% vs 17. 9%±0. 7%, t=9. 13, P<0. 001), S期细胞比例显著低于对照组 (34. 2%±0. 6% vs 38. 1%±0. 5%, t=6. 91, P<0. 001)(图6)。
2. 6 EME1沉默后的BEL-7404细胞凋亡的变化 Caspase3/7活性分析显示感染shRNA慢病毒5天后, 沉默组细胞的Caspase3/7 活性(145. 8%±5. 9%)较对照组组(100. 0%± 2. 3%) 显著升高1. 5倍 (t=12. 50, P<0. 001)(图7) 。
3 讨论
EME1对于哺乳动物细胞中 DNA 代谢的各个方面都很重要, Guo等[10]研究发现EME1与胃癌细胞AGS和MGC-803的增殖及侵袭能力密切相关, 沉默EME1基因表达可通过抑制蛋白激酶B活化, 降低糖原合酶激酶-3β、 细胞周期蛋白D1, 从而抑制癌细胞增殖和侵袭, 促进细胞凋亡。基于中国广西人群的研究[11] 发现EME1的Glu69Asp错义多态性与肝癌风险增加显著相关。EME1的另一外显子Ile350Thr变异会提高乳腺癌的易感性[12] 。
本研究发现沉默EME1的表达可降低肝癌的增殖能力。正常细胞中同源重组 (homologous recombination, HR)相关蛋白受到高度的调控, 对DNA序列具有相当高的特异性, 一旦失调就会导致基因组的不稳定性。MUS81/EME1在异常的时间节点被激活, 可能会导致细胞周期的阻滞、 DNA向子细胞的不对称分布以及非整倍体。相反,过早的磷酸化会刺激Holiday连接交叉。HR途径的过表达可能引起DNA重排, 导致癌基因活化或者抑癌基因的杂合性的缺失[13] 。HR介导基因表型的持续变化为肿瘤细胞提供生长优势, 恶性表型的不断发展以及耐药性的产生。有研究[14] 发现, 多发性骨髓瘤细胞和患者样本中的HR酶活性升高密切相关, 抑制HR活性可显著减少新基因表型的产生, 上调HR活性会增加基因突变的数量。持续获得HR介导的新基因, 可能会导致癌基因的激活, 促进肿瘤或耐药表型的获得。笔者推测高表达水平的EME1蛋白可能通过催化DNA-RNA杂交的形成导致基因组不稳定, 或通过促成同源性驱动的易出错过程 (如单链退火、 复制模板切换和非等位HR) 来促进突变。
结果表明, 沉默EME1表达后Caspase3/7活性显著增高, 有效促进细胞凋亡。细胞凋亡主要由Caspase家族蛋白酶执行, Caspase3/7负责对特定的细胞底物进行关键的切割, 导致最终的凋亡细胞死亡。癌前病变的DNA损伤可通过诱导细胞凋亡来清除潜在的有害细胞从而阻断肿瘤生长。这一死亡过程的失控与异常的细胞增殖、 肿瘤的发生和进展有关, 细胞凋亡的失调被认为是癌症的标志之一[15] 。本研究同时发现沉默EME1表达后G1期细胞比例高于shCtrl组, 表明抑制EME1表達能有效阻滞细胞周期, 抑制肿瘤细胞生长。EME1虽然在G2/M期活性受到严格的调控, 但其在细胞全周期中都能检测到。有研究[16] 证明有丝分裂中Cdk1活性的失调, 会过早激活MUS81/EME1的有丝分裂同源重组,可能会诱导DNA损伤应答缺陷相关的细胞周期改变。
蛋白激酶2 (CK2) 通过磷酸化调控MUS81/EME1复合物的活性, 从而在有丝分裂和DNA复制应激后发挥不同的功能。在细胞有丝分裂过程中, CK2通过磷酸化MUS81/EME1增强其活性, 有助于维持基因组的稳定性。但在细胞受到DNA复制应激后, CK2过度活化MUS81/EME1复合物会导致DNA损伤和染色体不稳定性[17]。事实上, 已经在许多肿瘤中发现CK2过表达[18-19] , 因此可以推测CK2可通过提高MUS81/EME1的生物学功能, 促进人类基因组的不稳定性, 还需要进一步研究确定肝癌中CK2的表达水平与MUS81/EME1的关系。在检查点激酶缺陷1的细胞中, 由于核苷酸的缺乏不足以维持有丝分裂期DNA合成, MUS81/EME1复合物切割有丝分裂期间合成的新生DNA[20] 。这种机制并不会导致细胞的死亡, 而是造成子细胞遗传了受损的DNA, MUS81/EME1复合物对晚期复制产物的异常处理会导致基因组不稳定。在FIBP敲低的肺腺癌中, EME1的过表达逆转了其对肺腺癌细胞增殖的抑制作用[21] 。该研究结果显示,EME1的过表达显著减少了 γ-H2AX 病灶形成, 表明EME1 参与肺腺癌细胞增殖和放射耐药性。FIBP 与STAT3互作通过增强其磷酸化从而提高转录活性, 并诱导靶基因EME1的表达。外源性STAT3的过表达也提高了FIBP缺陷肺腺癌细胞中的EME1表达水平, 这一结果在另一项研究[22] 中也得到证实。此外, 肿瘤细胞中的某些特异性的细胞因子常通过结合EME1的启动子上调其表达。
综上, EME1在肝癌组织中高表达, 并在细胞功能学层面证明了沉默EME1基因可抑制肝癌细胞的增殖, 阻滞细胞周期, 诱导肝癌细胞的凋亡, 为临床治疗提供了新思路。此外本研究对EME1在肝癌中发生发展限于细胞的功能性实验, 后续还需对动物模型和分子机制进行下一步的探索。
伦理学声明: 本研究于2020年7月14日通过解放军总医院第五医学中心伦理委员会审批, 批号: 2020055D。
利益冲突声明: 本文不存在任何利益冲突。
作者贡献声明: 纪冬负责课题设计及拟定写作思路; 陈椿、 王可欣、 贺梦雯进行实验研究及撰写论文; 王春艳、刘妍、 纪冬指导撰写文章并最后定稿。
参考文献:
[1] SUNG H, FERLAY J, SIEGEL RL, et al. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries[J]. CA Cancer J Clin, 2021, 71(3): 209-249. DOI: 10.3322/caac.21660.
[2] LLOVET JM, KELLEY RK, VILLANUEVA A, et al. Hepatocellular car?cinoma[J]. Nat Rev Dis Primers, 2021, 7: 6. DOI: 10.1038/s41572-020-00240-3.
[3] CHEN SH, WANG CY, GUO C, et al. Establishment of a nomogram model for predicting the survival of hepatitis B virus-related hepato?cellular carcinoma[J]. J Clin Hepatol, 2022, 38(7): 1566-1571. DOI: 10.3969/j.issn.1001-5256.2022.07.020.
陈松海, 王春艳, 郭畅, 等. 预测HBV相关肝细胞癌生存的列线图模型的建立[J]. 临床肝胆病杂志, 2022, 38(7): 1566-1571. DOI: 10.3969/j.issn.1001-5256.2022.07.020.
[4] WANG JJ, WANG KX, CHEN C, et al. Survival analysis and development of a prognostic nomogram for patients with hepatitis B virus-associated hepatocellular carcinoma[J]. Heliyon, 2023, 9(10): e20850. DOI: 10.1016/j.heliyon.2023.e20850.
[5] JI D, CHEN Y, BI JF, et al. Entecavir plus Biejia-Ruangan compound reduces the risk of hepatocellular carcinoma in Chinese patients with chronic hepatitis B[J]. J Hepatol, 2022, 77(6): 1515-1524. DOI: 10.1016/j.jhep.2022.07.018.
[6] GWON GH, JO A, BAEK K, et al. Crystal structures of the structure-selective nuclease Mus81-Eme1 bound to flap DNA substrates[J]. EMBO J, 2014, 33(9): 1061-1072. DOI: 10.1002/embj.201487820.
[7] HAN YY, ZHENG QY, TIAN Y, et al. Identification of a nine-gene panel as a prognostic indicator for recurrence with muscle-invasive bladder cancer[J]. J Surg Oncol, 2019, 119(8): 1145-1154. DOI: 10.1002/jso.25446.
[8] V?QUAUD E, DESPLANQUES G, J?Z?QUEL P, et al. Survivin con?tributes to DNA repair by homologous recombination in breast can?cer cells[J]. Breast Cancer Res Treat, 2016, 155(1): 53-63. DOI: 10.1007/s10549-015-3657-z.
[9] PENG L, LIANG J, WANG Q, et al. A DNA damage repair gene sig?nature associated with immunotherapy response and clinical prog?nosis in clear cell renal cell carcinoma[J]. Front Genet, 2022, 13: 798846. DOI: 10.3389/fgene.2022.798846.
[10] GUO ZG, LIANG EB, LI W, et al. Essential meiotic structure-specific endonuclease1 (EME1) promotes malignant features in gastric can?cer cells via the Akt/GSK3B/CCND1 pathway[J]. Bioengineered, 2021, 12(2): 9869-9884. DOI: 10.1080/21655979.2021.1999371.
[11] WANG YX, HUANG XL, SU ZH, et al. The Glu69Asp polymorphism of EME1 gene is associated with an increased risk of hepatocellular carcinoma in Guangxi population, China[J]. Int J Gen Med, 2022, 15: 7855-7866. DOI: 10.2147/IJGM.S383261.
[12] ZHAO JW, LIU L, ZHANG AQ, et al. Effect of EME1 exon variant Ile350Thr on risk and early onset of breast cancer in southern Chinese women[J]. J Biomed Res, 2013, 27(3): 193-201. DOI: 10.7555/JBR.27.20130013.
[13] SUZUKI T, YASUI M, HONMA M. Mutator phenotype and DNA double-strand break repair in BLM helicase-deficient human cells[J]. Mol Cell Biol, 2016, 36(23): 2877-2889. DOI: 10.1128/MCB.00443-16.
[14] SHAMMAS MA, SHMOOKLER REIS RJ, KOLEY H, et al. Dysfunc?tional homologous recombination mediates genomic instability and progression in myeloma[J]. Blood, 2009, 113(10): 2290-2297. DOI: 10.1182/blood-2007-05-089193.
[15] FULDA S. Tumor resistance to apoptosis[J]. Int J Cancer, 2009, 124(3): 511-515. DOI: 10.1002/ijc.24064.
[16] SZAKAL B, BRANZEI D. Premature Cdk1/Cdc5/Mus81 pathway acti?vation induces aberrant replication and deleterious crossover[J]. EMBO J, 2013, 32(8): 1155-1167. DOI: 10.1038/emboj.2013.67.
[17] PALMA A, PUGLIESE GM, MURFUNI I, et al. Phosphorylation by CK2 regulates MUS81/EME1 in mitosis and after replication stress[J]. Nucleic Acids Res, 2018, 46(10): 5109-5124. DOI: 10.1093/nar/gky280.
[18] GUO YG, ZHU ZH, HUANG ZY, et al. CK2-induced cooperation of HHEX with the YAP-TEAD4 complex promotes colorectal tumorigen?esis[J]. Nat Commun, 2022, 13(1): 4995. DOI: 10.1038/s41467-022-32674-6.
[19] KWON J, ZHANG JM, MOK B, et al. CK2-mediated phosphorylation up?regulates the stability of USP13 and promotes ovarian cancer cell prolif?eration[J]. Cancers, 2022, 15(1): 200. DOI: 10.3390/cancers15010200.
[20] PITRODA SP, PASHTAN IM, LOGAN HL, et al. DNA repair pathway gene expression score correlates with repair proficiency and tumor sensitivity to chemotherapy[J]. Sci Transl Med, 2014, 6(229): 229ra42. DOI: 10.1126/scitranslmed.3008291.
[21] XU YH, LI J, ZHU KK, et al. FIBP interacts with transcription factor STAT3 to induce EME1 expression and drive radioresistance in lung adenocarcinoma[J]. Int J Biol Sci, 2023, 19(12): 3816-3829. DOI: 10.7150/ijbs.83134.
[22] WEINANDY A, PIROTH MD, GOSWAMI A, et al. Cetuximab induces eme1-mediated DNA repair: A novel mechanism for cetuximab resis?tance[J]. Neoplasia, 2014, 16(3): 207-220, 220. e1-220. e4. DOI: 10.1016/j.neo.2014.03.004.
收稿日期:2023-10-11; 录用日期:2023-12-11
本文编辑:王莹
