18个玉米自交系主要性状配合力及遗传效应分析
2024-06-03张军刚冯晓曦郭海斌许海涛许波王成业
张军刚 冯晓曦 郭海斌 许海涛 许波 王成业
摘要:为评价新选育自交系的潜在利用价值,科学合理利用优良玉米自交系,指导玉米育种实践,用18个玉米自交系采用7×11不完全双列杂交设计配制77个组合,对单株产量等11个主要性状的配合力、遗传参数和相关性进行分析。一般配合力分析结果显示,M3、F1、M4、F2、F4等自交系产量及相关性状表现优良,M4、M10、F3、F6、M11等自交系宜机收性状表现优良;综合收获时籽粒含水量和单株产量的一般配合力表现,M4、F4、M11、M2、M9等自交系表现较好。特殊配合力分析结果显示,组合F2×M4单株产量的正向效应值最大,组合F3×M8收获时籽粒含水量的负向效应值最大;组合F2×M4、F1×M4、F4×M5、F1×M11的单株产量总配合力效应值较高,为优势组合,可进行进一步试验。遗传参数分析结果表明,株高、收获时籽粒含水量、雄穗分枝数和雄穗长可在早代选择;穗粗、穗位高可以早晚代结合选择;秃尖长、穗长、行粒数、单株产量应在晚代进行选择。相关性分析结果显示,单株产量与穗粗、穗长、穗位高和株高呈极显著正相关,与行粒数呈显著正相关,与收获时籽粒含水量呈正相关但不显著。研究结果可为玉米自交系利用及品种选育提供重要理论参考。
关键词:玉米;自交系;配合力;遗传参数;相关性
中图分类号:S513.032 文献标志码:A
文章编号:1002-1302(2024)08-0100-07
收稿日期:2023-05-29
基金项目:河南省玉米产业技术体系驻马店综合试验站项目(编号:HARS-22-02-Z6)。
作者简介:张军刚(1989—),男,河南清丰人,硕士,助理研究员,主要从事玉米遗传育种与栽培技术研究。E-mail:jungang39@163.com。
通信作者:王成业,研究员,主要从事玉米遗传育种与栽培技术研究。E-mail:13513865035@126.com。
在玉米育种中,配合力是评估自交系利用价值的重要指标,遗传力是确定目标性状选择世代的重要参数。通过对配合力和遗传力进行分析,明确亲本的遗传特性,根据育种目标选择优良亲本,可以显著提高育种效率[1]。贾恩吉等对13个玉米自交系的宜机收相关性状的配合力及遗传参数进行分析,选出机收性状配合力较好的自交系,探索各性状的遗传规律[2]。李文龙等对20份玉米双单倍体系种质的产量及相关农艺性状的配合力和遗传参数进行分析,筛选出产量性状配合力较高的3份玉米双单倍体系种质和5个玉米杂交组合[3]。进茜宁等对新选育的19个玉米自交系的10个农艺性状进行配合力、遗传参数和相关性分析,评估了所选自交系的应用价值[4]。罗黎明等分析了20个新选玉米自交系的产量和穗部性状的配合力,筛选到7份具有较高育种利用潜力的自交系[5]。孙轩等对玉米自交系京2416×京92改良后代收获时子粒含水量的配合力和遗传参数进行研究,筛选出优良杂交组合57个,认为收获时子粒含水量可以进行早代选择[6]。鉴于配合力和遗传力在育种亲本选择和杂交组合配制中的重要指导作用,研究玉米自交系的配合力和遗传效应,有助于认识自交系的应用价值,提高新品种的选育效率。
少数骨干玉米自交系在育种过程中的长期使用,导致我国玉米种质资源遗传基础贫乏,造成品种性状退化、抗逆性降低、稳产能力不足等[7],严重阻碍了我国玉米育种的进步。引入外源新种质并加以改良仍是玉米种质资源扩增的主要方式[8]。笔者所在课题组以美国、欧洲等国外种质资源为基础材料,经过改良选育出一批综合农艺性状优良的玉米自交系。本研究以新选育及外引的18个玉米自交系为试验材料,采用不完全双列杂交设计的方法组配77个杂交组合,对玉米单株产量及相关性状进行配合力、遗传参数以及相关性分析,筛选出表现优良的玉米自交系和优势杂交组合,以期为自交系的有效利用和改良提供一定的参考。
1 材料与方法
1.1 试验材料
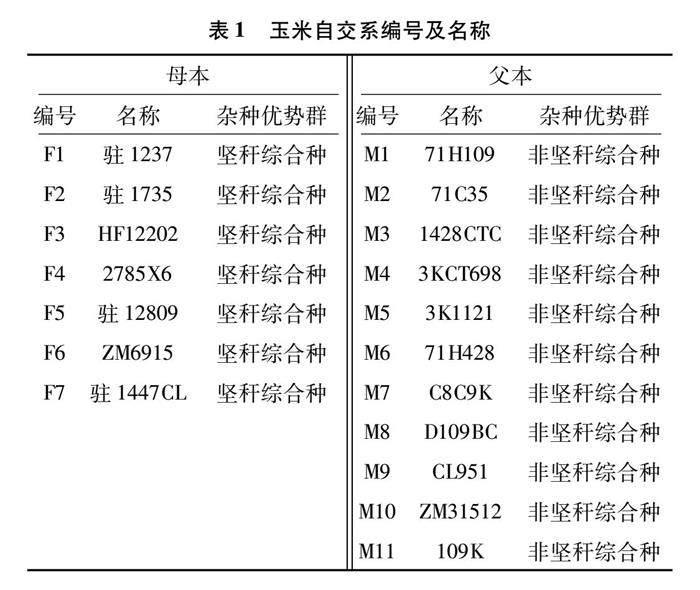
供试亲本材料为驻马店市农业科学院自选及外引的18个玉米自交系,其中母本材料7个,父本材料11个,名称及类群见表1。
1.2 试验方法
根据不完全双列杂交设计方法,2021年冬在海南省驻马店市农业科学院南繁基地进行杂交,组配77份杂交组合。2022年将77份杂交组合种植在驻马店市农业科学试验站(33°01′N,114°05′E)。试验采用完全随机区组设计,3行区,行长4 m,行距 0.6 m,3次重复,种植密度为7.5万株/hm2,田间管理同一般大田生产。
1.3 调查项目
在灌浆期选择具有代表性的植株调查株高、穗位、雄穗长及其分枝数;成熟期收获当天使用绿洲牌LDS-1G谷物水分测定仪直接测定籽粒含水量;收获小区中间1行连续10穗,自然风干后,对穗长、穗粗、秃尖长、行粒数、穗行数、单株产量(含水量为14%)进行室内考种。
1.4 数据处理方法
应用SAS 9.1软件对株高、穗长、单株产量等11个性状进行方差分析。根据黄远樟等的方法[9]计算一般配合力和特殊配合效应值,并计算遗传参数。采用SPSS 22.0软件进行相关性分析。
2 结果与分析
2.1 方差分析
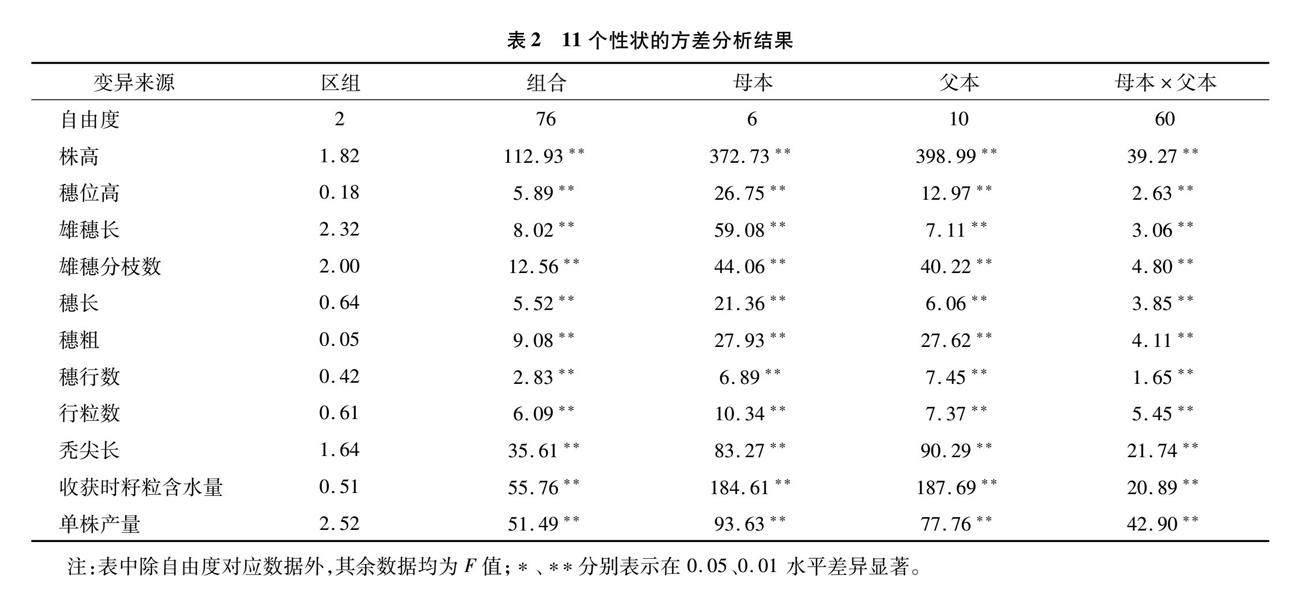
对77份杂交组合11个农艺性状进行方差分析,结果(表2)显示,11个农艺性状表型F值在区组间均未达到显著水平,在亲本及组合间均达到极显著水平,说明各性状的差异主要是由遗传差异造成的,可以进行下一步的配合力分析。
2.2 一般配合力效应分析
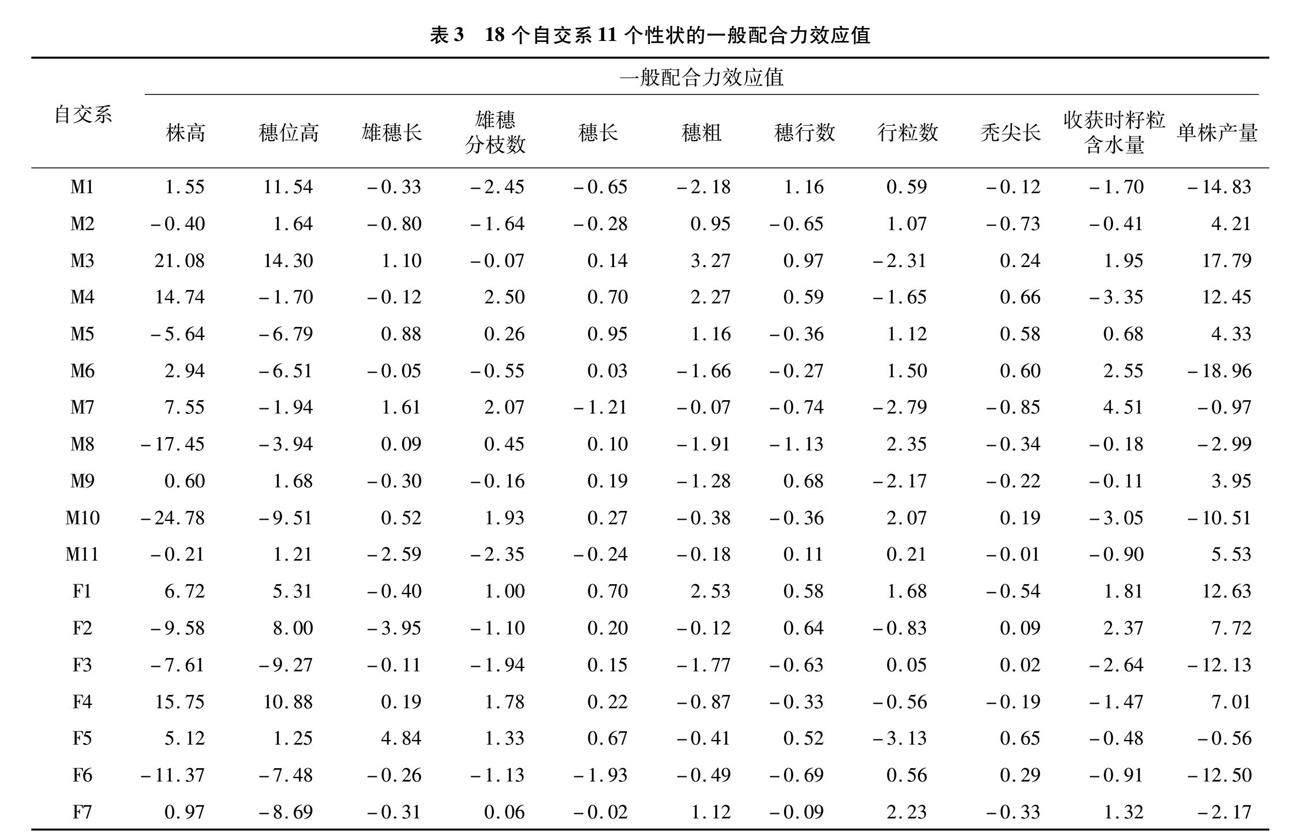
一般配合力指一个亲本与多个亲本杂交后,杂种后代在某个性状上的平均表现,主要由基因的加性效应决定,是可以稳定遗传的部分[10]。自交系一般配合力高,表明该系所含有的有利基因位点多,在育种实践中具有较高的利用价值,组配优良杂交种的概率大[11]。通过分析18个亲本自交系11个性状的一般配合力效应值(表3)可知,一般配合力效应值在同一自交系不同性状间和同一性状不同自交系间存在较大差异。单株产量一般配合力效应值变幅为-18.96~17.79,效应值为正值的有9个,排名前3的亲本为M3、F1、M4,有利于提高杂交后代的单株产量;株高一般配合力效应值变幅为 -24.78~21.08,负向效应值最高的3个亲本为M10、M8、F6,可用于降低雜交后代株高,提高抗倒性;穗位高一般配合力效应值变幅为-9.51~14.30,负向效应值最高的3个亲本为M10、F3、F7,有利于降低杂交后代的穗位高,提高抗倒性;雄穗长一般配合力效应值变幅为-3.95~4.84,正向效应值最高的3个亲本为F5、M7、M3,有利于增加花粉量,提高结实率;雄穗分枝数一般配合力效应值变幅为-2.45~2.50,负向效应值最高的3个亲本为M1、M11、F3,有利于减少养分消耗,提高产量;穗长一般配合力效应值变幅为-1.93~0.95,正向效应值最高的3个亲本为M5、M4、F1,有利于选育出穗长较长的杂交后代;穗粗一般配合力效应值变幅为-2.18~3.27,正向效应值最高的3个亲本为M3、F1、M4,可用于选育穗粗较粗的杂交后代;穗行数一般配合力效应值变幅为-1.13~1.16,正向效应值最高的3个亲本为M1、M3、M9,有利于选育穗行数较多的杂交后代;行粒数一般配合力效应值变幅为-3.13~2.35,正向效应值最高的3个亲本为M8、F7、M10,有利于增加杂交后代的行粒数;秃尖长一般配合力效应值变幅为-0.85~0.66,负向效应值最高的3个亲本为M7、M2、F1,有利于减小秃尖长,提高结实率;收获时籽粒含水量一般配合力效应值变幅为-3.35~4.51,负向效应值最高的3个亲本为M4、M10、F3,可用于选育籽粒含水量低的自交系或品种。
2.3 特殊配合力效应分析
特殊配合力是2个亲本通过组配产生的杂交种的平均水平,主要由基因间的显性或超显性以及上位性等非加性效应控制,在杂交育种中不能稳定地遗传给后代[10]。特定组合的特殊配合力高,表明该杂交组合性状表现优异,是杂交组合选配的重要参考依据。对测算的特殊配合力效应值进行归类,结果(表4)显示,秃尖长特殊配合力为负向效应值的组合个数较多,其他10个性状特殊配合力效应值为正值和负值的组合个数差异较小。组合F4×M11株高和穗位高的负向效应值最大,表明其降低组合株高和穗位高的效果突出。组合F1×M4雄穗长的正向效应值最大,组合F5×M1雄穗分枝数的负向效应值最大,组合F1×M11穗长的正向效应值最大,组合F4×M2穗粗的正向效应值最大,组合 F3×M2穗行数正向效应值最大,组合F4×M5行粒数的正向效应值最大,组合F5×M2秃尖长的负向效应值最大,组合F3×M8收获时籽粒含水量的负向效应值最大,组合F2×M4单株产量的正向效应值最大。
2.4 单株产量总配合力效应值分析
总配合力是杂交双亲一般配合力和组合特殊配合力效应值之和,能够充分反映组合的配合力效应和产量潜力水平,杂交组合某性状总配合力值越大,说明其亲本间的杂种优势越大,田间表现越优异[12-14]。77个杂交组合的单株产量总配合力效应值变幅为-74.71~59.91,其中效应值为正值的组合有40个。为深入了解优异杂交组合的综合特性,对总配合力排名前20位的组合进行分析(表5)。亲本一般配合力较高,其组配的杂交种特殊配合力不一定高,如F1×M3、F2×M3等;亲本一般配合力较低,其组配的杂交种特殊配合力却较高,如F7×M9、F4×M1等;实际测产结果与总配合力效应值排名完全一致,与特殊配合力排名不完全一致,说明总配合力更能体现杂交组合的实际表现。
总配合力效应值大的组合可以分为3类:(1)双亲一般配合力和组合特殊配合力效应值都高,如F2×M4、F1×M4、F4×M5等;(2)双亲之一一般配合力效应值较高且组合特殊配合力效应值较高,如F7×M3、F2×M7等;(3)双亲一般配合力效应值较高、组合特殊配合力效应值较低,如F2×M3、F1×M3。本研究中F2×M4、F1×M4、F4×M5、F1×M11这4个组合单株产量总配合力较高,表现突出,为优势组合,可进一步鉴定其丰产性、稳产性和适应性,挖掘其应用潜力。
2.5 各性状遗传效应和遗传参数估计
为进一步认识玉米各性状的遗传效应,本试验对11个性状的遗传参数进行了研究,结果见表6。从各性状的配合力方差看,株高、穗位高、雄穗长、雄穗分枝数、穗粗、穗行数和收获时籽粒含水量的一般配合力方差明显高于特殊配合力方差,表明这7个性状的遗传由加性效应主导。穗长和秃尖长的特殊配合力方差较高,且与一般配合力方差差异不大,表明这2个性状在后代遗传中非加性效应和加性效应均比较重要。单株产量、行粒数的一般配合力方差明显低于特殊配合力方差,表明这2个性状由非加性效应主导。
从各性状的遗传力看,除穗行数外,其他10个性状的广义遗传力均大于60%,狭义遗传力大于50%的性状依次为株高、收获时籽粒含水量、雄穗分枝数和雄穗长,这些性状遗传比较稳定,可在早代选择;穗粗、穗位高的狭义遗传力分别为47.49%、44.25%,可以早晚代结合选择;秃尖长、穗长、行粒数、单株产量狭义遗传力较低,这些性状适合在晚代进行选择。穗行数广义遗传力较低,说明该性状受环境影响较大,遗传不稳定。
2.6 各性状表型的相关性分析
分析77个玉米杂交组合11个性状间的相关性,结果(表7)显示,单株产量与穗粗、穗长、穗位高和株高呈极显著正相关,与行粒数呈显著正相关;收获时籽粒含水量与穗粗呈显著正相关,与秃尖长呈显著负相关;株高与穗位高、穗长、穗粗和穗行数呈极显著正相关;穗位高与穗行数呈极显著正相关,与穗长呈显著正相关;雄穗长与雄穗分枝数呈极显著正相关;穗长与行粒数呈极显著正相关,与穗粗呈显著正相关;穗粗与穗行数呈极显著正相关;行粒数与秃尖长呈显著负相关。
3 结论与讨论
玉米育种成功的关键在于选择优良的亲本自交系,但杂交后代的优劣不仅取决于自交系农艺性状的好坏,还取决于自交系配合力的高低[5,15]。通过对玉米自交系进行配合力分析可以鉴别自交系的利用价值[16]。本研究以7个母本和11个父本为亲本,组配77个杂交组合,对11个主要农艺性状进行配合力分析,综合产量及相关性状的一般配合力表现可知,M3、F1、M4、F2、F4等表现较好,可以作为组配高产杂交种的亲本,在组配杂交组合过程中利用性状的遗传互补实现选育优良组合的目的[17];综合株高、穗位高、收获时籽粒含水量等宜机收性状的一般配合力表现,效应值较低的有M4、M10、F3、F6、M11,在适宜机械收获的品种或自交系选育中有较大的利用价值。综合单株产量和收获时籽粒含水量的一般配合力表现,M4、F4、M11、M2、M9等自交系表现较好,在选育高产、宜机收玉米品種及自交系时的应用潜力较大。在育种工作中,根据具体的育种目标筛选适当的自交系作为亲本,有针对性地配制杂交组合,才有可能选育出优良的玉米品种[18]。
在不完全双列杂交遗传设计中,今后的研究重点应放在目标性状的总配合力效应上,通过总配合力对目标性状做出全面判断[19]。通过分析不同组合的单株产量总配合力效应值发现,大多总配合力效应值高的组合,至少有1个亲本的一般配合力效应值高,且杂交组合特殊配合力效应值也较高,但也有个别总配合力效应值高的组合,双亲的一般配合力效应值高,而杂交组合特殊配合力效应值却很低。因此,在组配杂交组合时,至少选用1个一般配合力高的亲本,在此基础上,选择特殊配合力高的组合,可以提高优良组合的选择效率,这与前人的观点[20-22]一致。单株产量总配合力更能反映杂交组合的实际单株产量,可以作为评价杂种优势的指标[23-24]。本研究中,单株产量总配合力效应值最高的4个组合依次为F2×M4、F1×M4、F4×M5、F1×M11,下一步可对这4个优良组合进行多年多点种植鉴定,以确定其应用价值。
遗传力反映了亲本性状传递给后代的能力大小,可分为广义遗传力和狭义遗传力,根据性状遗传力的高低可预测目标性状的有效选择世代[25-27]。杨彦忠等根据各性状的遗传力认为,株高、穗长、雄穗分枝数、穗行数和穗粗等性状可在早代选择;秃尖长可以在早、晚代结合选择;单株产量、穗位高、行粒数适合在高代进行选择[28]。邹成林等研究认为,穗位高、穗粗、穗行数、秃尖主要由加性效应决定,株高、穗长、行粒数和单株产量主要由非加性效应决定,穗位高和秃尖可以进行早代选择,穗粗、穗行数、株高、穗长、行粒数和单株产量宜在晚代进行选择[18]。彭林等研究认为,株高及收获时含水量宜早代选择,穗位高及雄穗分枝数宜晚代选择[29]。闫海霞等研究认为,穗长、穗行数可进行早代选择;穗粗、行粒数、单株粒重等受环境影响较大,不宜进行早代选择[30]。本研究中,株高、收获时籽粒含水量、雄穗分枝数和雄穗长等性状可在早代进行选择;穗粗、穗位高应早晚代结合选择;秃尖长、穗长、行粒数、单株产量适合在晚代进行选择;穗行数广义遗传力为39.44%,受环境影响较大。株高、穗位高、穗粗、穗行数、雄穗长、收获时籽粒含水量和雄穗分枝数主要受加性效应影响;穗长和秃尖长的非加性效应和加性效应均比较重要;单株产量、行粒数主要由非加性效应主导。本研究结果与前人研究既有相同之处,又有所不同,产生的差异可能是由材料选择、地理条件、生产条件、气候条件的不同引起的。
玉米很多性状间都存在相关性,有的为正相关,性状间相互促进,有的为负相关,此消彼长。玉米各性状之间的相关性分析结果可为玉米育种实践提供依据。本研究相关性分析结果表明,单株产量与株高呈极显著正相关,表明高大的植株更易获得较高的产量。单株产量与收获时籽粒含水量呈正相关但不显著,相关系数为0.211,说明生育期长、晚熟品种易获得较高的产量。单株产量与穗粗、穗长呈极显著正相关,表明粗穗型、长穗型品种更易获得较高的产量。秃尖长与行粒数、收获时籽粒含水量呈显著负相关,表明秃尖越长,行粒数越少,收获时籽粒含水量越低。玉米机械粒收是我国玉米机械收获的发展方向和今后玉米生产方式的重点[31]。收获时籽粒含水量低、早熟耐密是宜粒收玉米品种的关键指标[32],因此,在宜机收玉米品种选育中,应尽量寻找单株产量与株高、收获时籽粒含水量等性状间的平衡点。
参考文献:
[1]桑晓慧,陈 伟,赵云雷,等. 陆地棉16份材料配组的产量和纤维品质性状配合力分析[J]. 棉花科学,2021,43(4):24-31.
[2]贾恩吉,张东升,李雨泽,等. 玉米自交系宜机收的农艺性状配合力分析[J]. 江苏农业科学,2022,50(20):107-110.
[3]李文龙,龙 舟,任国鑫,等. 20份玉米DH系配合力及遗传参数研究[J]. 黑龙江八一农垦大学学报,2022,34(3):35-40.
[4]进茜宁,李 威,王平喜,等. 玉米自交系的配合力及相关性分析[J]. 江苏农业科学,2021,49(6):68-72.
[5]罗黎明,蒋辅燕,高连彰,等. 20个新选玉米自交系产量和穗部性状配合力及其相关性研究[J]. 西南农业学报,2021,34(10):2084-2092.
[6]孙 轩,吉玉龙,张如养,等. 玉米自交系京2416×京92改良后代收获时子粒含水量配合力分析[J]. 玉米科学,2021,29(3):1-7,16.
[7]庹美琴,高 翔,高雪飞,等. 我国玉米种质资源研究现状及思考[J]. 新疆农垦科技,2021,44(4):68-69.
[8]赵 璞,温之雨,董文琦,等. 我国玉米资源研究现状及发展展望[J]. 中国种业,2019(10):8-11.
[9]黄远樟,刘来福. 作物数量遗传学基礎——六、配合力:不完全双列杂交[J]. 遗传,1980(2):43-46.
[10]史秀秀,毕晓静,马守才,等. 黄淮麦区杂交小麦亲本的杂种优势和配合力分析[J]. 麦类作物学报,2013,33(6):1111-1118.
[11]丰 光,赵洪绪,王孝杰,等. 玉米自交系郑58×PH6WC二环选系的配合力研究[J]. 玉米科学,2021,29(5):22-27.
[12]卢秉生,高洪敏,姚永祥,等. 10个玉米自交系穗部性状的配合力和遗传参数分析[J]. 作物杂志,2017(2):23-28.
[13]张艳茹,王 伟,唐 兰,等. 52份热带玉米和温带玉米配合力及杂种优势分析[J]. 四川农业大学学报,2021,39(4):467-476.
[14]鲁俊田,任丽丽,赵洪绪,等. Iodent玉米种质改良旅大红骨选系配合力及杂种优势利用研究[J]. 玉米科学,2020,28(6):18-24.
[15]张 征,张雪丽,莫博程,等. 籼型杂交水稻农艺性状的配合力研究[J]. 作物学报,2017,43(10):1448-1457.
[16]左淑珍,王光申,汤金涛,等. 14份新育玉米自交系应用潜力分析[J]. 玉米科学,2013,21(2):31-35.
[17]Wang B B,Hou M,Shi J P,et al. De novo genome assembly and analyses of 12 founder inbred lines provide insights into maize heterosis[J]. Nature Genetics,2023,55:312-323.
[18]邹成林,谭 华,黄开健,等. 24份热带玉米自交系主要农艺性状的遗传效应分析[J]. 西南农业学报,2022,35(7):1500-1508.
[19]韩登旭,邵红雨,阿布來提·阿布拉,等. 几个早熟玉米自交系配合力和遗传参数分析[J]. 新疆农业科学,2010,47(8):1541-1545.
[20]余世权,苟才明,黄 宁,等. 17个玉米地方种质选系的育种潜势分析[J]. 中国农学通报,2010,26(22):81-86.
[21]苟才明,余世权,黄 宁,等. 17个玉米地方种质选系的配合力分析[J]. 华北农学报,2015,30(2):175-182.
[22]韩登旭,杨 杰,阿布来提·阿布拉,等. 骨干玉米自交系种子活力性状配合力及遗传参数分析[J]. 西北农业学报,2021,30(9):1290-1300.
[23]崔 超,高聚林,于晓芳,等. 18个玉米自交系氮效率相关性状的配合力分析[J]. 作物学报,2014,40(5):838-849.
[24]高旭东,周旭梅,高洪敏,等. 欧洲玉米种质BRC选系主要农艺性状的配合力及杂种优势分析[J]. 玉米科学,2015,23(3):28-33.
[25]袁 凯,逯腊虎,杨 斌,等. 不同粒重小麦品种主要农艺性状的配合力和遗传力分析[J]. 麦类作物学报,2018,38(6):661-667.
[26]史民芳,安林利,丁国庆,等. 矮秆超大穗小麦亲本与多抗丰产小麦主要农艺性状的配合力评价及遗传分析[J]. 中国农学通报,2012,28(36):50-56.
[27]焦宇馨,张宇翔,杨文艳,等. 结合辅助性状的玉米全基因组选择预测力评估[J]. 江苏农业学报,2023,39(2):313-320.
[28]杨彦忠,周玉乾,连晓荣,等. 玉米自交系KA105和91227的配合力分析[J]. 中国种业,2021(2):45-50.
[29]彭 林,慈佳宾,杨 巍,等. 宜机收玉米品种选育及配合力分析[J]. 分子植物育种,2021,19(6):2073-2080.
[30]闫海霞,柳家友,付家锋,等. 10个自选玉米自交系的穗部性状配合力及遗传参数分析[J]. 山东农业科学,2018,50(10):30-32.
[31]《玉米科学》编辑部. 加强机械粒收研究推动玉米生产方式转型[J]. 玉米科学,2018,26(4):62.
[32]王元东,赵久然,张华生,等. 玉米宜粒收品种的选育及种质创新策略[J]. 分子植物育种,2020,18(10):3455-3460.
