干旱胁迫下哈茨木霉对烟草幼苗生理指标的影响
2024-06-03冯召杨盟权姚峰李先锋彭志良殷全玉
冯召 杨盟权 姚峰 李先锋 彭志良 殷全玉

摘要:为了探究干旱胁迫下哈茨木霉对烟草幼苗生理指标的影响,以烤烟品种K326为试验材料,采用蘸根法进行哈茨木霉处理,用不同浓度(10%、20%)聚乙二醇(PEG-6000)模拟不同程度(轻度、中度)的干旱胁迫,用沙培法研究不同程度干旱胁迫下哈茨木霉对烟草幼苗表型、叶片相对含水率、根系发育、光合色素含量、膜脂过氧化指标、渗透调节物质含量、保护酶活性的影响。结果表明,在正常清水处理条件下,哈茨木霉蘸根处理相比于清水条件更能够促进烟株生长,提高叶片相对含水率,促进根系发育,如总根长、投影面积、根表面积、分枝数、连接数等指标,增加烟叶中叶绿素a、叶绿素b含量,降低烟叶中过氧化氢、丙二醛含量,增加烟叶中脯氨酸、可溶性糖、可溶性蛋白含量,提升烟叶中过氧化氢酶、过氧化物酶、超氧化物歧化酶、抗坏血酸过氧化物酶活性。与清水对照相比,在轻度、中度干旱胁迫下,烟株变黄萎蔫,相对叶片含水率降低,总根长、投影面积、根表面积、连接数受到抑制,叶绿素a和叶绿素b含量减少,过氧化氢和丙二醛含量增加,脯氨酸、可溶性糖、可溶性蛋白含量增加,过氧化氢酶、过氧化物酶、超氧化物歧化酶、抗坏血酸过氧化物酶活性增加。在轻度和中度干旱的条件下,与同等程度干旱胁迫条件相比,哈茨木霉蘸根处理能够明显改善烟叶萎蔫变黄现象,增加叶片相对含水率,促进根系发育,如总根长、投影面积、根表面积、连接数,提高绿素a、叶绿素b、脯氨酸、可溶性糖、可溶性蛋白含量以及过氧化氢酶、过氧化物酶、超氧化物歧化酶、抗坏血酸过氧化物酶活性,降低过氧化氢和丙二醛含量。由研究结果可以看出,不同程度干旱胁迫均可对烟株造成不同程度的损伤,哈茨木霉蘸根处理可促进烟草发育、提高烟草的抗旱性,从而降低干旱胁迫对烟草造成的危害。
关键词:烟草;干旱胁迫;哈茨木霉;生理指标;根系;生长发育
中图分类号:S572.01 文献标志码:A
文章编号:1002-1302(2024)08-0086-08
收稿日期:2023-10-12
基金项目:河南省科技攻关项目(编号:232102110053);中国烟草总公司陕西省公司科技项目(编号:2022610700260183、2021611000270042)。
作者简介:冯 召(1999—),男,河南武陟人,硕士研究生,主要从事烟草栽培生理生化研究。E-mail:2467212544@qq.com。
通信作者:殷全玉,博士,副教授,主要从事烟草资源微生物利用与土壤保育研究。E-mail:quanyuy@126.com。
随着各国各行业的高速发展,全球气候受到影响,限制了各地农业生产[1]。干旱在气候影响中占有较大比例,加上各地降水不足、水资源匮乏,对农作物的生产产生了巨大影响[2-3]。干旱胁迫会抑制植物生长发育、破坏植物的正常生理代谢活动与植物的防御系统、减弱植物的光合作用、减少干物质的积累,从而极大地降低农作物的产量与质量[4]。王晶等研究发现,干旱可显著抑制沙芦草幼苗生长及根系发育,降低其光合色素含量,提高丙二醛含量[5]。杨云等研究发现,干旱可导致薏苡幼苗叶片干枯,降低叶片相对含水率,减弱光合作用[6]。刘硕等研究发现,干旱胁迫可提升云蔗08-1609、粤糖93-159 等甘蔗品种的抗氧化酶活性,从而清除过多的活性氧(ROS)[7]。孙晓梵等研究发现,在干旱胁迫下,狗牙根幼苗的电导率升高,渗透调节物质得到积累,从而使细胞内部环境保持稳定[8]。
烟草是我国重要的经济作物,在整个生育期对水分都有较高要求,缺少水分会对烟草根系发育及体内代谢活动产生严重影响[9]。我国不少烟区饱受水资源缺乏的困扰,使得烟叶的产量、品质大幅下降[10]。如今我国已有大量关于筛选抗旱品种烤烟的研究,这些研究一般通过对烤烟抗旱性指标进行比较分析,或在生产中大力推广抗旱烤烟品种,但是,目前通过微生物作用来提高烤烟抗旱性的研究尚少。
木霉菌广泛存在于植物体及根际土壤中,具有促进植物生长、提高抗性的能力。在植物根系定植的过程中,木霉菌丝缠绕在植物根系周围,形成附着胞状结构并能够存活较长时间[11]。它还能释放植物激素,明显促进种子萌发、幼苗生长、根系生长及分解利用土壤养分,从而提高植物的抗逆性[12]。殷全玉等研究发现,哈茨木霉可对烟草起到显著的促生作用,可以促进根系发育,提高抗病性[13]。李怿聪等研究发现,棘孢木霉可使玉米叶片叶绿素含量保持较高水平,增强光合作用,增加干物质的积累[14]。孙悦燕等研究发现,根际接种木霉可提高华北落叶松的渗透调节物质含量,提高其抗氧化酶活性,增強华北落叶松自身的抗性[15]。刘爱荣等研究发现,木霉可促进黄瓜根系发育,抑制黄瓜尖孢镰刀菌,从而提高黄瓜的抗病性[16]。
目前,哈茨木霉在作物领域已有大量研究,但关于哈茨木霉对烟草抗旱性影响的研究极少。因此,本研究选择在生产实践中应用较广泛的烤烟品种K326作为研究对象,通过聚乙二醇(PEG-6000)模拟干旱胁迫条件,检测干旱胁迫下哈茨木霉对烟草幼苗的根系发育、光合色素含量、膜脂过氧化指标、植物渗透调节物质和保护性酶活性的影响规律,揭示干旱胁迫下哈茨木霉对烟草的生理响应规律,以期为促进烟草生产发展提供借鉴。
1 材料与方法
1.1 供试材料
供试烟草品种为K326,种子由河南农业大学烟草学院育种实验室提供;供试菌株为哈茨木霉CGMCC2394,由河南农业大学烟草学院微生物实验室提供;PEG-6000购买于天津市科密欧化学试剂有限公司;改良型霍格兰营养液购于Coolaber公司。
1.2 试验设计
试验于2022年11月在河南农业大学许昌校区现代科教园区人工气候室内进行,将烟草种子播种于育苗盘中,并放入人工气候室中进行培养,培养条件如下:室温25 ℃,光照时间07:00—21:00,待长至3~4张真叶时,取长势一致的烟苗36株,将根部基质洗净后分为2组,每18株1组,分别放在清水、哈茨木霉孢子悬浮液(1×108 CFU/mL)中蘸根5 h,将每株烟苗移栽到装有石英砂、蛭石、珍珠岩体积比为1 ∶1 ∶1的塑料钵(上下边长和高分别为10、7、8 cm)中,将36个塑料钵分别放入盛有 1/4 霍格兰营养液(2 L)的6个塑料盆中,缓苗3 d,得到具有试验条件的烟苗,缓苗结束后更换营养液开始干旱处理,干旱处理期间使用1/2霍格兰营养液。
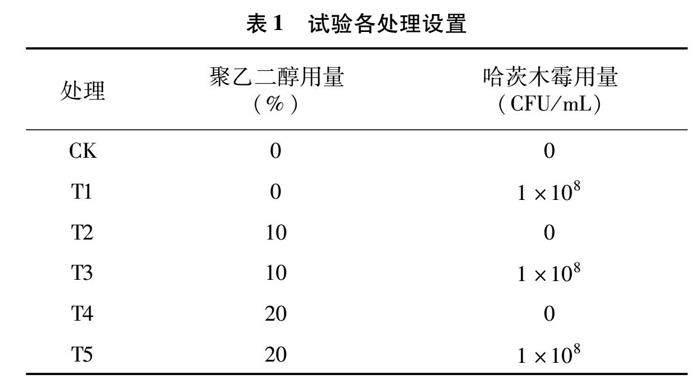
试验共6个处理,其中CK为正常清水处理,T1处理为正常清水+哈茨木霉处理,T2处理为轻度干旱处理,T3处理为轻度干旱+哈茨木霉处理,T4处理为中度干旱处理,T5处理为中度干旱+哈茨木霉处理(表1)。每3 d更换1次营养液,6 d后进行取样,每个处理随机选取3株烟苗,将根系清洗干净后进行分析,叶片用液氮速冻后保存于-80 ℃冰箱中,备用待测。每份烟叶样品分别测定叶片相对含水率、光合色素含量、膜脂过氧化指标、渗透调节物质含量和保护酶活性。
1.3 指标及测定方法
1.3.1 叶片相对含水率 叶片相对含水率采用张建锴等的方法[17]进行测定。取1张新鲜烟叶,称取鲜重(WF);再将烟叶浸水24 h,取出后用吸水纸擦干样品表面水分,称取饱和重(WT);最后将烟叶放入105 ℃烘干箱中,杀青15 min,然后于80 ℃下烘至恒重,称取干重(WD)。计算公式如下:
相对含水率=(WF-WD)/(WT-WD)×100%。
1.3.2 根系发育 将根系清洗干净后,用植物根系扫描仪(EPSON Expression 12000 XL)扫描根系外观形态,用根系专业化分析软件(WinRHIZO)分析投影面积、总根长、根表面积、分枝数、连接数。
1.3.3 光合色素含量 用乙醇提取比色法测定叶绿素a、叶绿素b、类胡萝卜素含量[18]。
1.3.4 膜脂过氧化指标 叶片过氧化氢含量的测定采用南京建成科技有限公司的过氧化氢检测试剂盒。丙二醛含量采用北京索莱宝科技有限公司的试剂盒进行测定。
1.3.5 渗透调节物质 脯氨酸、可溶性糖、可溶性蛋白含量均采用北京索莱宝科技有限公司的试剂盒进行测定。
1.3.6 保护酶活性 过氧化氢酶、过氧化物酶、超氧化物歧化酶、抗坏血酸过氧化物酶的活性采用北京索莱宝科技有限公司的试剂盒进行测定。
1.4 数据处理
数据用Excel 2019、SPSS 26.0进行方差分析,并检验数据的差异显著性,用Origin 2021作图。
2 结果与分析
2.1 干旱胁迫下哈茨木霉对烟草叶片相对含水率及表型的影响
由图1-A可知,正常清水+哈茨木霉处理(T1处理)的叶片相对含水率最高,为89.35%,比CK增加6.48%;轻度干旱处理(T2处理)、中度干旱处理(T4处理)的叶片相对含水率分别比CK降低12.75%、28.27%;轻度干旱+哈茨木霉处理(T3处理)的叶片相对含水率比T2处理提高8.51%,中度干旱+哈茨木霉处理(T5处理)的叶片相对含水率比T4处理提高12.36%。由图1-B、图1-C、图 1-D 可知,烟草受到干旱胁迫时,出现叶片失水萎蔫、叶片发黄、烟株发育受阻的现象;经过哈茨木霉蘸根处理的烟株在干旱条件下有较好的表型特征,多体现在叶面积增大、叶片发黄现象减少、烟株生长发育受到促进。受到轻度干旱胁迫的烟株出现叶片萎蔫、叶片偏小的现象,受到中度干旱胁迫的烟株出现明显叶片萎蔫,同时烟叶面积极大减小,表明哈茨木霉蘸根处理可明显减轻干旱胁迫对烟株造成的伤害。研究结果显示,哈茨木霉处理可以提高叶片相对含水率,使烟株更好地应对干旱胁迫,尤其对中度干旱胁迫的损伤有著更明显的缓解作用,可明显提高烟株的抗旱能力。
2.2 干旱胁迫下哈茨木霉对烟草根系发育的影响
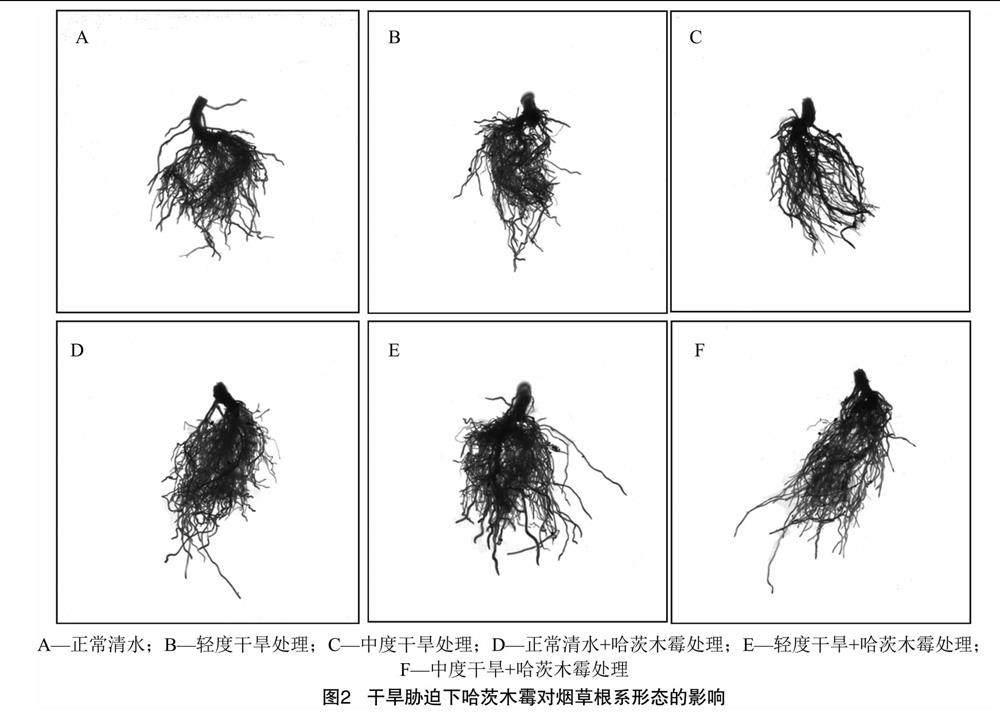
由图2、表2可知,T1处理的总根长、投影面积、根表面积、分枝数、连接数都处于最高水平,与CK相比分别增加49.12%、31.45%、32.83%、71.30%、65.23%;T2、T4处理的总根长、投影面积、根表面积、连接数与CK相比分别降低12.52%、4.19%、8.39%、29.11%和34.04%、27.00%、22.08%、36.14%;T3处理的总根长、投影面积、分枝数、连接数比T2处理分别增加6.16%、6.02%、3.58%、12.40%,T5处理的总根长、投影面积、根表面积、分枝数、连接数分别比T4处理增加30.28%、45.60%、43.43%、26.58%、25.71%。研究结果表明,干旱胁迫可明显抑制根系的生长,哈茨木霉蘸根处理可促进烟草根系发育并壮根, 缓解干旱胁迫的损伤,从而增强烟草根系吸收营养的能力。
2.3 干旱胁迫下哈茨木霉对烟叶光合色素含量的影响
由图3可知,T1处理的叶绿素a、叶绿素b含量与CK相比分别提高11.16%、14.50%,类胡萝卜素含量分别降低13.21%;T2、T4处理的叶绿素a、叶绿素b含量分别比CK降低10.36%、46.88%及6.80%、19.96%,类胡萝卜素含量分别提高30.58%、13.47%;T3处理的叶绿素a、叶绿素b含量分别比T2处理提高11.72%、2.94%,T5处理的叶绿素a、叶绿素b含量分别比T4提高18.99%、59.46%。说明哈茨木霉蘸根处理可使烟株在干旱的情况下仍可保持较高的叶绿素含量,增强烟株的光合作用,增加干物质的积累量,从而提高烟株的抗旱性。
2.4 干旱胁迫下哈茨木霉对烟叶膜脂过氧化指标的影响
由图4-A可知,T1处理的过氧化氢含量最低,比CK降低38.39%;T2、T4处理的过氧化氢含量分别比CK提高100.93%、147.57%;T3处理的过氧化氢含量比T2处理降低46.22%,T5处理的过氧化氢含量比T4处理降低49.41%。
由图4-B可知,T1处理的丙二醛含量最低,比CK降低21.47%;T2、T4处理的丙二醛含量分别比CK提高9.82%、91.82%;T3处理的丙二醛含量比T2处理降低5.96%,T5处理的丙二醛含量比T4处理降低41.16%。研究结果表明,随着干旱程度的加强,过氧化氢、丙二醛含量呈明显的增加趋势;哈茨木霉蘸根处理可清除烟株体内的过氧化物,使烟株在干旱条件下仍可保持较低的过氧化氢、丙二醛含量,减缓烟草膜脂过氧化,并且在中度干旱胁迫时效果最明显。
2.5 干旱胁迫下哈茨木霉对烟叶渗透调节物质的影响
由图5-A可知,与CK相比,T1处理的脯氨酸含量提高116.90%;T2、T4处理的脯氨酸含量分别比CK提高195.72%、628.74%;T3处理的脯氨酸含量比T2处理提高26.48%,T5处理的脯氨酸含量比T4处理提高25.45%。
由图5-B可知,与CK相比,T1处理的可溶性糖含量提高26.95%;T2、T4处理的可溶性糖含量分别比CK提高6.53%、12.04%;T3处理的可溶性糖含量比T2处理提高21.57%,T5处理的可溶性糖含量比T4处理提高6.31%。
由图5-C可知,与CK相比,T1处理的可溶性蛋白含量提高36.59%;T2处理、T4处理的可溶性蛋白含量分别比CK提高50.28%、74.20%;T3处理的可溶性蛋白含量比T2处理提高15.37%,T5处理的可溶性蛋白含量比T4处理提高15.37%。研究结果表明,在正常清水条件下,哈茨木霉蘸根处理可明显提高烟株的渗透调节物质含量,增加养分的积累量。干旱胁迫可明显提高烟株体内脯氨酸、可溶性糖、可溶性蛋白含量,使其依靠其自身防御系统应对干旱胁迫,而经哈茨木霉蘸根处理后,可进一步提高干旱胁迫下烟草的脯氨酸含量,增强调控水分的能力,增强抗旱能力。
2.6 干旱胁迫下哈茨木霉对烟叶保护酶活性的影响
由图6-A可知,T1处理的过氧化氢酶活性最高,比CK高5.43%;T2、T4处理的过氧化氢酶活性分别比CK低32.07%、54.89%;T3处理的过氧化氢酶活性比T2处理高21.60%,T5处理的过氧化氢酶活性比T4处理高44.58%。研究结果表明,干旱胁迫可明显降低烟草过氧化氢酶活性,且随着干旱程度的增强呈下降趋势;哈茨木霉蘸根处理可提高烟株在干旱条件下的过氧化氢酶活性,提高烟株的抗氧化能力,增强抗旱性。
由图6-B可知,T1处理的过氧化物酶活性比CK高66.82%;T2、T4处理的过氧化物酶活性分别比CK高59.35%、 80.84%; T3处理的过氧化物酶活性比T2处理高27.57%,T5处理的过氧化物酶活性比T4处理高27.65%。
由图6-C可知,T1处理的超氧化物歧化酶活性比CK高96.56%;T2、T4处理的超氧化物歧化酶活性分别比CK高94.56%、176.01%;T3处理的超氧化物歧化酶活性比T2处理高22.18%,T5处理的超氧化物歧化酶活性比T4处理高17.62%。
由图6-D可知,T1处理的抗坏血酸过氧化物酶活性比CK高53.53%;T2、T4处理的抗坏血酸过氧化物酶活性分别比CK高71.72%、95.74%;T3处理的抗坏血酸过氧化物酶活性比T2处理高11.35%,T5处理的抗坏血酸过氧化物酶活性比T4处理高4.97%。结果表明,在正常清水处理的条件下,经哈茨木霉蘸根处理可明显提高烟株的抗氧化酶活性;在干旱胁迫下,过氧化物及其他小分子大量增加,自身防御系统产生抗氧化酶以保护自身细胞膜,提高过氧化物酶、超氧化物歧化酶、抗坏血酸过氧化物酶活性;经哈茨木霉蘸根处理后,烟株在干旱条件下能保持更高的抗氧化酶活性。
3 讨论
干旱胁迫对烟草最直接的影响就是导致烟草叶片变黄萎蔫、叶片相对含水率降低,从而抑制叶片的生长。在本研究中, 干旱胁迫导致烟草叶片相
對含水率下降、叶片萎蔫卷曲,叶面积增长及生长发育受到抑制,该研究结果与张杰等的研究结果[19]一致。经哈茨木霉蘸根处理的烟草在干旱胁迫下可提高叶片相对含水率,保持叶片舒展,促进烟株生长发育,这与王秀丽等的研究结论[20]一致。
根系是植物吸收养分的关键器官,干旱胁迫可导致根系发育受阻,减少干物质积累,影响烟草的产量和质量[21]。良好的根系具有较大的总根长、根体积、分枝数和连接数,能够提高根系从土壤中获取养分资源的能力。本研究中,干旱胁迫可显著抑制根系的生长,其中适度的干旱可增加烟草根系的根尖数和分枝数,以提高根系吸收养分的能力,使其更加适应干旱胁迫,这与张旭东等的研究结论[22]一致;哈茨木霉蘸根处理可明显促进根的生长,增大根体积,形成健壮的根系,使其在干旱条件下仍可保持良好的根系状态,增强吸收养分的能力,更好地应对干旱胁迫,增强烟株的抗旱性。邓薇等的研究表明,木霉在干旱条件下可通过增强玉米幼苗根系的养分吸收能力,减少水分散失,使幼苗更好地利用土壤水分[23],本研究结论与之一致。
光合色素在植物中起到光能转化并起到固碳作用,植物的光合能力及光能利用转化能力可以用光合色素含量来反映。干旱胁迫可破坏叶绿体的结构,降解叶绿素,导致光能无法被充分利用,提高类胡萝卜素含量。本研究中,干旱胁迫明显导致了叶绿素a含量的下降,并不同程度地影响叶绿素b和类胡萝卜素含量,这与张环纬等的研究结论[24]一致;经哈茨木霉蘸根处理的烟株在干旱胁迫下保持较高水平的光合色素含量,表明哈茨木霉可能减轻了干旱对叶绿素的损伤,从而增加烟草的光合色素含量,增强烟株的光合作用,这与王建等的研究结论[25]一致。
植物在遭受逆境胁迫时会产生大量的ROS,破坏细胞膜系统,可导致烟草体内活性氧平衡被打破,生成大量的过氧化氢,释放大量的中间产物丙二醛,引起膜脂过氧化。即过氧化氢和丙二醛作为膜脂过氧化指标,一定程度上可以反映植物的保护能力。本研究中干旱胁迫可导致烟株体内过氧化氢和丙二醛含量显著增加,说明干旱导致了膜脂过氧化,这与丁丹阳等的研究结果[26]一致;经哈茨木霉蘸根处理的烟株在干旱条件下可降低过氧化氢和丙二醛的含量,这说明哈茨木霉可减缓膜脂过氧化程度,降低干旱胁迫对细胞膜的危害,这与王秀丽等的研究结论[27-28]一致。
脯氨酸、可溶性糖和可溶性蛋白作为渗透调节物质在烟草体内可起到调节渗透势的作用,可增强烟株调控水分的能力,从而提高烟株的抗旱性。本研究中,干旱胁迫可通过自身防御系统减少物质的消耗,积累渗透调节物质,应对干旱胁迫,这与李鹏辉等的研究结果[29]一致;哈茨木霉蘸根处理可提高烟草脯氨酸、可溶性糖和可溶性蛋白含量,增强烟草的保水性,保持较高的抗旱性,这与Racic等的研究结果[30]一致。
过氧化氢酶、超氧化物歧化酶、过氧化物酶和抗坏血酸过氧化物酶作为抗氧化酶,当烟草受到干旱胁迫时会产生大量的过氧化物,在过氧化物清除中,抗氧化酶起到至关重要的作用,降低干旱胁迫对烟株造成的损伤。本研究结果表明,干旱胁迫可提高烟草体内超氧化物歧化酶、过氧化物酶和抗坏血酸过氧化物酶活性,降低过氧化氢酶活性,这可能是因为过氧化氢酶在抗氧化过程中没有起到主要作用或调节能力有限,这与邵惠芳等的研究结论[31-32]一致;而哈茨木霉蘸根处理可进一步提高烟草的过氧化氢酶活性,清除体内过氧化物,增强烟草的抗氧化性,这与杨静雅等的研究结论[33]一致。
4 结论
本研究通过干旱胁迫对烟草生长的影响以及哈茨木霉对烟草抗旱性的影响,测定烟草根系发育情况,叶片的光合色素含量、膜脂过氧化指标、渗透调节物质含量和保护酶活性,通过分析指标变化规律。结果表明,在干旱胁迫下,烟草的生长发育受阻,叶片相对含水率下降,光合色素和渗透调节物质含量降低,膜脂过氧化加重,酶活性提高;哈茨木霉在正常培养条件下可促进烟草生长发育;干旱胁迫下,哈茨木霉蘸根处理可显著改善烟草表型,提高叶片相对含水率,提高光合色素含量,缓解膜脂过氧化,增加渗透调节物质,提高保护酶活性,增强烟草自身的防御能力。本研究说明哈茨木霉可提高烟草抗旱性,可为哈茨木霉在大田生产中提高烟草抗旱性提供一定的理论参考。
参考文献:
[1]陈兰兰,王 丽,吴亚娟,等. 植物响应干旱胁迫的分子和微生态机制[J/OL]. 分子植物育种,(2023-04-07)[2024-04-24]. http://kns.cnki.net/kcms/detail/46.1068.S.20230406.1634.006.html.
[2] Yang X Y,Lu M Q,Wang Y F,et al. Response mechanism of plants to drought stress[J]. Horticulturae,2021,7:50.
[3]武慧敏,吕爱锋,张文翔. 巴音河流域水文干旱对气象干旱的响应[J]. 南水北调与水利科技(中英文),2022,20(3):459-467.
[4]莫言玲,郑俊鶱,杨瑞平,等. 不同西瓜基因型对干旱胁迫的生理响应及其抗旱性评价[J]. 应用生态学报,2016,27(6):1942-1952.
[5]王 晶,伏兵哲,李淑霞,等. 外源褪黑素对干旱胁迫下沙芦草幼苗生长和生理特性的影响[J]. 应用生态学报,2023,34(11):2947-2957.
[6]杨 云,周 宇,班秀文,等. 干旱胁迫对薏苡幼苗形态和生理特征的影响[J/OL]. 分子植物育种. (2023-07-07)[2024-04-24]. http://kns.cnki.net/kcms/detail/46.1068.S.20230706.1405.006.html.
[7]刘 硕,樊 仙,李如丹,等. 4个甘蔗主栽品种对干旱胁迫的生理响应[J]. 热带作物学报,2022,43(9):1812-1823.
[8]孙晓梵,张一龙,李培英,等. 不同施氮量对干旱下狗牙根抗氧化酶活性及渗透调节物质含量的影响[J]. 草业学报,2022,31(6):69-78.
[9]梁 栋,刘光亮,王 永,等. 干旱胁迫对烟草不同部位细胞程序化死亡的影响[J]. 中国烟草科学,2020,41(4):33-40.
[10]朱晓萌,张泽中,袁义杰,等. 基于水分亏缺率的贵州地区烟草干旱指标[J]. 水利水电科技进展,2021,41(5):34-40,88.
[11]李 阔,王红阳,郭秀芝,等. 木霉属真菌诱导根及根茎类中药材抗根腐病的研究及应用进展[J]. 中国中药杂志,2023,48(18):4942-4949.
[12]Rush T A,Shrestha H K,Gopalakrishnan M M,et al. Bioprospecting Trichoderma:a systematic roadmap to screen genomes and natural products for biocontrol applications[J]. Front Fungal Biology,2021,2:716511.
[13]殷全玉,匡志豪,王 景,等. 黑胫病不同抗性烤烟品种对哈茨木霉的生理响应[J]. 河南农业科学,2022,51(9):88-98.
[14]李怿聪,黄凤麟,吴少彤,等. 棘孢木霉对盐碱农田玉米根际土壤及叶片光合特性的影响[J/OL]. 分子植物育种. (2023-04-24)[2024-04-24]. http://kns.cnki.net/kcms/detail/46.1068.S.20230424.0919.002.html.
[15]孙悦燕,王秀丽,高润梅,等. 干旱胁迫下华北落叶松幼苗接种木霉的生理变化[J]. 应用生态学报,2021,32(3):853-859.
[16]刘爱荣,陈双臣,陈 凯,等. 哈茨木霉对黄瓜尖孢镰刀菌的抑制作用和抗性相关基因表达[J]. 植物保护学报,2010,37(3):249-254.
[17]張建锴,曹红霞,潘小燕,等. 基于产量和品质的陕北苹果滴灌水量和追施氮量优化研究[J]. 干旱地区农业研究,2020,38(5):143-152.
[18]李合生. 植物生理生化实验原理与技术[M]. 北京:高等教育出版社,2000:134-137.
[19]张 杰,马晓寒,陈 彪,等. 外源硫化氢对干旱胁迫下烟草幼苗生理生化特性的影响[J]. 中国农业科技导报,2018,20(11):112-119.
[20]王秀丽,柳昱旻,高润梅. 外生菌根真菌对干旱条件下油松幼苗生长的影响[J]. 中国水土保持科学,2019,17(2):70-76.
[21]李 佳,刘 涛,马菊莲,等. 烟草响应干旱胁迫与抗旱遗传育种研究进展[J]. 江苏农业科学,2023,51(8):34-43.
[22]张旭东,王智威,韩清芳,等. 玉米早期根系构型及其生理特性对土壤水分的响应[J]. 生态学报,2016,36(10):2969-2977.
[23]邓 薇,张祖衔,曹宇航,等. 绿色木霉缓解干旱胁迫对玉米幼苗根系生长的影响[J]. 山东农业科学,2022,54(2):40-45.
[24]张环纬,陈 彪,温心怡,等. 外源硅对干旱胁迫下烟草幼苗生长、叶片光合及生理指标的影响[J]. 生物技術通报,2019,35(1):17-26.
[25]王 建,赵 单. 丛枝菌根真菌与绿色木霉对干旱胁迫下苹果树苗生长、生理特征及水分利用的影响[J]. 江苏农业科学,2023,51(4):164-170.
[26]丁丹阳,张艺洁,王凯悦,等. 喷施拉肖皂苷C提高烟草幼苗抗干旱胁迫能力的生理机制[J]. 中国农业科技导报,2018,20(12):36-44.
[27]王秀丽,高润梅. 根际接种木霉对干旱胁迫下华北落叶松幼苗生长的影响[J]. 东北林业大学学报,2019,47(9):81-84.
[28]陈 赛,倪敏姿,崔 峰,等. 海洋生境棘孢木霉TCS007缓解蚕豆干旱胁迫的研究[J]. 农药学学报,2023,25(3):657-667.
[29]李鹏辉,向金友,王 林,等. 干旱胁迫下外源褪黑素对烟草幼苗生理特性的影响[J]. 中国农业科技导报,2019,21(5):41-48.
[30]Raciá G,VukeliáI,ProkiáL,et al. The influence of Trichoderma brevicompactum treatment and drought on physiological parameters,abscisic acid content and signalling pathway marker gene expression in leaves and roots of tomato[J]. Annals of Applied Biology,2018,173(3):213-221.
[31]邵惠芳,陈 征,许嘉阳,等. 两种烟草幼苗叶片对不同强度干旱胁迫的生理响应比较[J]. 植物生理学报,2016,52(12):1861-1871.
[32]刘元玺,吴俊文,王丽娜,等. 云南松幼苗生长及生理生化特性对持续干旱胁迫的响应[J/OL]. 分子植物育种. (2023-05-19)[2024-04-24]. http://kns.cnki.net/kcms/detail/46.1068.S.20230518.1319.004.html.
[33]杨静雅. 木霉菌对菘蓝促生抗旱效应研究[D]. 保定:河北大学,2021.
