不同钾肥对烤烟叶片钾吸收规律的影响
2024-06-03卢平曾彦清马扬扬黄鹤鸣胡兴川陈思原吴志高
卢平 曾彦清 马扬扬 黄鹤鸣 胡兴川 陈思原 吴志高
摘要:为探究在水肥一体化技术下追施不同钾肥对烤烟叶片钾吸收规律的影响,为烤烟质量调控提供参考。以云烟99为研究对象,设置不同钾肥为试验因素,以当地常用钾肥(硫酸钾)作为对照(CK),设置磷酸二氢钾(T1)、焦磷酸钾(T2)、黄腐酸钾(T3)3个钾肥处理,研究不同处理对烤烟生育期叶片钾含量、叶片钾积累量与积累速率、钾通道基因表达量及烤后烟常规化学成分的影响。结果表明,与移栽后30 d相比,在烤烟移栽后90 d,CK、T1、T2、T3处理叶片钾含量分别降低了60.19%、52.75%、56.31%、48.54%,降低幅度表现为CK>T2>T1>T3。在烤烟移栽后45~60、75~90 d,T3处理烤烟叶片钾的积累量、积累速率高于其他处理。在烤烟移栽后50 d,T1处理NtKC1基因相对表达量为CK处理的5.15倍。在烤烟移栽后65 d,T1、T3处理的NtHAK基因相对表达量分别为CK处理的4.27、3.77倍,T3处理NtNHX2基因的相对表达量为CK处理的3.46倍。在烤烟移栽后90 d,T3處理NtKC1基因的相对表达量为CK处理的7.98倍。在T3处理下,烤后烟上、中部叶钾含量分别较CK处理提高了10.00%、33.33%,烤后烟中部叶钾氯比较CK处理提高了107.41%。在水肥一体化技术下追施黄腐酸钾可以显著降低烤烟生育后期叶片钾的流失量,提高叶片钾的积累量及积累速率,诱导NtHAK、NtNHX2基因表达,并提高烤后烟中上部叶钾含量、中部叶钾氯比。
关键词:水肥一体化技术;钾肥;吸收;钾通道基因相对表达量
中图分类号:S572.06 文献标志码:A
文章编号:1002-1302(2024)08-0080-06
收稿日期:2023-06-08
基金项目:福建中烟工业有限责任公司科技计划(编号:2021350000340469)。
作者简介:卢 平(1972—),男,福建福州人,农艺师,主要从事烟草原料学研究。E-mail:331222069@qq.com。
通信作者:吴志高,农艺师,主要从事烟草生产和管理研究。E-mail:84848383@qq.com。
钾离子是植物生长发育过程中必需的大量元素之一,其含量约占植物干物质量的0.3%~5.0%,植物生长发育过程和生理代谢过程的多个环节均有钾离子的参与和调控[1]。钾离子对烟草品质有重要影响,它不仅能改善烟叶的燃烧性、降低焦油含量,并且显著影响了烤后烟叶的可用性[2]。在低钾胁迫下,烟碱含量增加,烟叶身份过厚,细胞间隙增大,烟株矮小且生长缓慢,叶尖、叶缘有杂色斑点,严重时甚至导致烟株死亡[3-4]。王英锋等的研究结果表明,钙浓度为5 mmol/L的水培条件最大限度缓解了低钾胁迫造成的烟株生长缓慢及钾吸收抑制现象[5]。巫彬芳研究证实,滴灌施肥可以大幅提高烟叶钾含量[6],这与席奇亮的研究结果[7]一致。江文林的研究发现,环割、断根处理及运用降碱提钾制剂、植物生长调节剂等方式均达到了提升烟叶钾含量、改善烟叶品质的效果[8]。胡珑瀚通过喷施不同叶面肥和分段式增施钾肥的方式,也达到了烟叶提钾的效果[9]。
目前,一般认为植物对钾离子的吸收有2套机制,即高钾浓度下的低亲和性钾吸收机制及低钾浓度下的高亲和性钾吸收机制[10]。高亲和性钾吸收为逆浓度梯度运输的主动吸收过程,一般发生在外界钾离子浓度为1~200 μmol/L时,需通过细胞质膜上的载体协助完成,其载体被称为“离子泵”,低亲和性钾吸收是顺浓度梯度运输的被动吸收过程,主要发生在外界钾离子浓度为1~10 mmol/L时,其运输通道被称为“离子通道”[10-12]。植物吸收钾离子的过程需要多种蛋白质参与,从分子水平上看,KUP/HAK/KT、HKT和CPA家族的钾转运蛋白是钾离子主动运输过程的主要参与者,而Shaker、TPK、Kir-like家族的钾离子通道蛋白主要参与钾离子的被动运输过程[13-14]。
Shaker家族是在烟草钾吸收与转运中研究最早的转运蛋白,目前已报道的烟草Shaker钾通道基因共有12个,分别为圆锥烟草的NpKT1,普通烟草的NKT1、NKT2、NtKC1、NTORK1、NKT3、NKT4、NKT5、NtKAT3、NtSKOR,林烟草的NKT6和黄花烟草的NKC1[15-23]。其中,NKT1、NKT2、NtKC1为内向整流钾通道基因,与拟南芥的钾通道基因AKT1、AKT2、AtKC1同源,NTORK1为外向整流钾通道基因,与拟南芥的钾通道Group V同源。有研究发现,高浓度水杨酸和低浓度萘乙酸可以诱导NTORK1表达,从而促进烟草根系钾离子外流。NKT3、NKT5和拟南芥AtKC1基因同源[16-17,19]。NKT4主要在烟草的根部表达,与烟草的低钾、干旱胁迫有关[18]。NtKAT3基因在烟草的根、茎、叶及花中均有表达,与烟草的低钾、高盐胁迫有关[20]。与NTORK1一样,NtSKOR也为外向整流钾通道基因,可能在调节烟草的低钾、高盐、干旱、低温等非生物胁迫中发挥了重要作用[21]。NKT6与拟南芥KAT1/KAT2同源,主要定位于细胞膜和核膜附近的内质网,其在林烟草的茎、腋芽、萼片、叶、花、根中均有表达,在外源施加脱落酸处理与干旱胁迫下,该基因的表达量下降,可能与林烟草的气孔运动有关[22]。
目前对烟草TPK家族基因的研究较少,仅报道了NtTPK1、NtTPK2这2个TPK家族的钾通道基因。其中NtTPK1存在于液泡膜中,对K+有高度的选择性,在盐胁迫、高渗透压胁迫下,该基因的部分异构体表达量显著提高[23]。NtTPK2是从普通烟草红花大金元品种中克隆得到的,在烟草根、茎、叶、花中均有表达[24]。
钾转运体主要分为KUP/HAK/KT、HKT和CPA这3个家族。KUP/HAK/KT家族最早在大肠杆菌中被发现,在植物中的研究较晚,此类钾转运体广泛存在于拟南芥、水稻、玉米等植物体内[25-28]。2009年Guo等从黄花烟草中克隆得到了烟草中的首个钾转运基因——NrHAK1,其与烟草根部的钾吸收过程相关[29]。鲁黎明等在普通烟草中克隆得到基因NtHAK1,其在烟草K326品种中过表达后,烟株的富钾能力显著提高[30-31]。除此之外,还有主要在叶、茎、花中表达的烟草NtPOT10基因及在林烟草盛花期各组织中表达的HAK11基因等,各基因均在烟草的胁迫响应机制中扮演着重要角色[32-33]。HKT家族主要参与控制K+/Na+的选择性运输和K+的吸收,陆玉建等将AtHKT1启动子导入到本生烟草细胞中,其β-D-葡萄糖苷酸酶(β-glucuronidase,GUS)染色结果显示,本生烟草的叶片、根部及维管系统染色较深,表明这些区域AtHKT1表达量可能较高,有利于提高植物的耐盐性[34]。
目前已有研究证实,水肥一体化技术可以促进烟株对钾离子的吸收[35]。本研究在水肥一体化技术的基础上设计追施不同钾肥的处理,研究不同钾肥对烤烟叶片钾吸收规律及钾通道基因表达量的影响,拟筛选出对烤烟提钾效果最佳的钾肥品种,以提高烤烟叶片钾含量,提升烟叶品质,并将其应用在大田生产上,指导大田生产活动。
1 材料与方法
1.1 试验地概况
试验于2022年在云南省大理白族自治州祥云县开展,土壤类型为黄褐土,土壤pH值为6.80,碱解氮含量为51.45 mg/kg,速效磷含量为9.67 mg/kg,速效钾含量为92.00 mg/kg,氯含量为66.03 mg/kg,有机质含量为11.35 g/kg。
1.2 试验设计
选用当地主栽品种云烟99作为供试材料。各处理均在移栽前1周每666.67 m2施入29 kg复合肥,复合肥选用当地烟草专用肥,N、P2O5、K2O的施用比例为12 ∶10 ∶25,采用水肥一体化技术追肥和灌水。根据钾肥品种不同,试验共设置4个处理,其中CK追施当地常用钾肥品种硫酸钾,T1处理追施磷酸二氢钾,T2处理追施焦磷酸钾,T3处理追施黄腐酸钾。追肥时,为了保证各处理在整个生育期施入的N、P、K总量相同,在移栽后30、45 d,T1、T2、T3处理使用自制液体专用肥。每666.67 m2的具体追肥方案见表1。
1.3 测定项目及方法
1.3.1 烤烟生育期叶片钾含量的测定 在烤烟移栽后30、45、60、75、90 d,随机选取各处理烤烟各3株,分别取上、中、下3个叶位的新鲜叶片3张,每次采样时,每株采摘相同叶位的鲜叶片,鲜叶片在105 ℃杀青30 min后转于60 ℃烘干至恒重,研磨后过0.25 mm(60目)筛网,用质量分数4%的盐酸浸提-火焰光度计法测定钾含量[36]。
1.3.2 烤烟叶片钾积累量及积累速率的测定 分别计算在烤烟移栽后30~45、45~60、60~75、75~90 d烤烟叶片钾的积累量及积累速率,其中,钾离子积累量=相邻2次取样时间间隔钾离子含量之差,钾离子积累速率=钾离子积累量/相邻2次取样时间间隔。
1.3.3 钾通道基因相对表达量的测定 在烟叶移栽后50、65、90 d,各个处理选取具有代表性的烟株3株,每株选取自下而上第3、10、15叶位的3 g叶片并混合,在-80 ℃超低温条件下保存,用荧光定量PCR法测定钾通道基因的相对表达量。
1.3.4 烤后烟常规化学成分的测定 分别选取各处理上、中部叶烤后样1 kg,用近红外法检测叶片的常规化学成分含量。
1.4 数据处理与分析
数据用Excel 2013、SPSS 26.0进行处理分析,绘图用Excel 2013。
2 结果与分析
2.1 追施不同钾肥对烤烟生育期叶片钾含量的影响
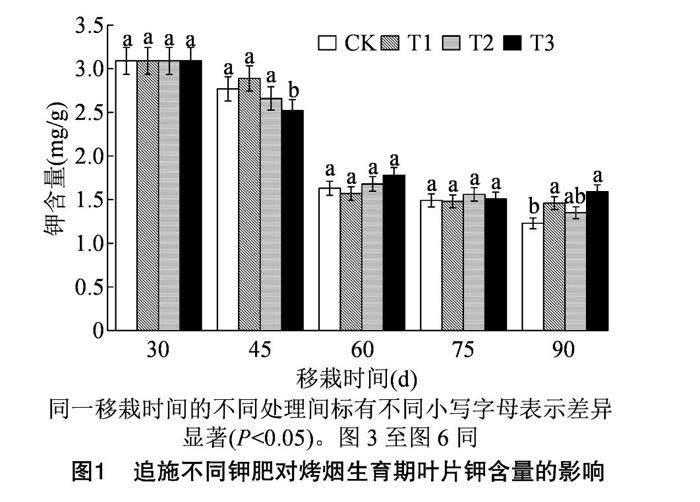
由图1可知,在烤烟移栽后45 d,T3处理葉片钾含量显著低于CK处理;施入钾肥后,在烤烟移栽后的60、75 d,各处理叶片钾含量无显著差异;在烤烟移栽后的90 d,T1、T3处理烤烟叶片钾含量显著高于CK处理;与移栽后30 d相比,在烤烟移栽后90 d,各处理烤烟叶片钾含量分别降低了60.19%、52.75%、56.31%和48.54%,其中T1、T3处理叶片钾的损失量显著低于CK处理。上述结果表明,追施磷酸二氢钾、黄腐酸钾显著降低了烤烟叶片钾的损失量。
2.2 追施不同钾肥对烤烟生育期叶片钾积累量及积累速率的影响
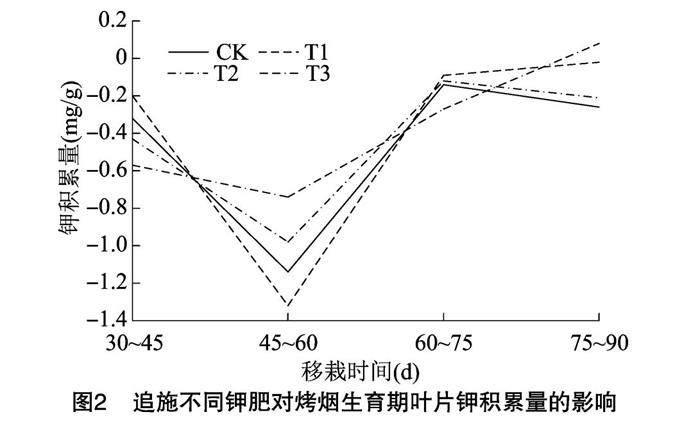
由图2可知,叶片钾积累速率的变化规律与叶片钾积累量的变化规律一致。在烤烟移栽后 45~60 d,各处理烤烟叶片中钾的积累量及积累速率均达到最低值,在这一时期各处理烤烟叶片中钾的积累量及积累速率表现为T3>T2>CK>T1;T1、T3处理烤烟叶片中钾的积累量及积累速率在烤烟移栽后75~90 d达到高峰,CK、T2处理烤烟叶片中钾的积累量及积累速率在烤烟移栽后60~75 d达到高峰。因此可见,追施黄腐酸钾可能促进了烤烟移栽后45~60、75~90 d叶片中钾的积累量及积累速率的提高,从而促进了烤烟叶片对钾离子的吸收。
2.3 追施不同钾肥对烤烟叶片钾通道基因表达量的影响
追施不同钾肥对烤烟叶片钾通道基因表达量的影响见图3至图6。NtHAK基因是KUP/HAK/KT
家族中调节钾素吸收的钾离子转运蛋白基因。由图3可知,在烤烟移栽后的65 d,T1、T2、T3处理NtHAK基因的相对表达量显著高于CK处理,分别为CK处理的4.27、2.25、3.77倍,表明在水肥一体化技术下,追施不同钾肥均促进了烤烟移栽后65 d NtHAK基因的表达,其中磷酸二氢钾、黄腐酸钾的效果显著优于焦磷酸钾。
CPKs基因在植物中广泛存在,已有研究证实,CPKs家族的NtCPK4、NtCPK5基因为钙依赖型的蛋白激酶基因[37]。由图4可知,在烤烟移栽后50 d,T1、T2、T3处理的NtCPK基因相对表达量显著低于CK处理,但在烤烟移栽后65、90 d,T1、T2处理叶片NtCPK基因相对表达量显著高于CK处理,在移栽后65 d分别为CK处理的1.92、1.73倍,在移栽后90 d分别为CK处理的2.15、1.77倍。这表明水肥一体化技术下追施不同钾肥均促进了烤烟移栽后65、90 d NtCPK基因的表达。
NHX常作为抗盐方面的基因被研究,但相关研究发现,该基因也可以促进植物大量吸收、积累钾离子[38]。由图5可知,在烤烟移栽后50 d,T1处理NtNHX2基因的相对表达量显著高于CK处理,为CK处理的1.69倍;在烤烟移栽后65 d, T1、 T3处理NtNHX2基因的相对表达量显著高于CK处理,
分别为CK处理的1.67、3.46倍。上述结果表明,在水肥一体化技术下追施的磷酸二氢钾、黄腐酸钾诱导了烤烟移栽后65 d叶片中NtNHX2基因的表达,对烤烟叶片中钾的积累产生有利影响。
NtKC1基因是Shaker家族的一种内向整流型钾离子通道基因。由图6可知,在烤烟移栽后的 50 d,T1、T2处理烤烟中NtKC1基因的相对表达量显著高于CK处理,分别为CK处理的5.15、3.09倍;在烤烟移栽后的65 d,T3处理烤烟中NtKC1基因的相对表达量为CK处理的2.56倍,显著高于CK处理;在烤烟移栽后的90 d,T2、T3处理烤烟中NtKC1基因的相对表达量显著高于CK处理,分别为CK处理的7.98、2.15倍。上述结果表明,在水肥
一体化技术下追施黄腐酸钾促进了烤烟移栽后65、90 d NtKC1基因的表达,追施焦磷酸钾促进了烤烟移栽后90 d NtKC1基因的表达,对烤烟叶片钾的积累产生有利影响。
2.4 追施不同钾肥对烤后烟常规化学成分的影响
由表2可知,追施不同钾肥后,各处理烤烟各叶位的化学成分含量均表现为总植物碱含量偏低、两糖含量适宜、 总氮含量为最适范围。在T1、T3处理下,上部烤烟叶片中的钾含量分别较CK高6.00%、10.00%,T1、T2、T3处理烤烟中部叶的钾含量分别较CK高25.49%、20.59%、33.33%;T2处理烤烟上部叶的氯含量较CK低20.00%,T2、T3处理烤烟中部叶的氯含量分别较CK低21.43%、35.71%;T2处理烤烟上部叶的钾氯比较CK高25.00%,T1、T2、T3处理烤烟中部叶钾氯比分别较CK高3.29%、53.50%、107.41%。除T2处理上部叶外,与CK处理相比,追施不同钾肥均提高了烤后烟上、中部叶的钾含量,效果表现为黄腐酸钾>磷酸二氢钾>焦磷酸钾,追施不同钾肥均提高了烤后烟中部叶的钾氯比,效果表现为黄腐酸钾>焦磷酸钾>磷酸二氢钾。
3 讨论与结论
钾离子是烟草生长及品质形成的关键物质。本研究在水肥一体化技术下追施不同钾肥,研究烤烟生育期叶片钾离子的变化规律,钾离子的积累规律以及钾通道基因相对表达量的变化规律,并检测了烤后烟的常规化学成分。研究结果表明,追施磷酸二氢钾和黄腐酸钾可以显著降低烤烟生长发育期间叶片钾的损失量,与移栽后30 d相比,在烤烟移栽后90 d,CK、T1、T2和T3处理叶片钾含量分别降低了60.19%、52.75%、56.31%和48.54%,降低幅度表现为CK>T2>T1>T3;在烤烟移栽后45~60 d,各处理叶片钾的积累量及积累速率均达到最低峰,在这一时期T3处理钾积累量及积累速率明显高于其他处理,在烤烟移栽75 d后,CK、T2处理叶片钾积累量及积累速率开始下降,但T1、T3处理仍保持上升趋势,且T3处理升高更为明显,追施黄腐酸钾可能提高了烤烟移栽后45~60、75~90 d叶片钾的积累量及积累速率。
對钾通道基因表达量的研究结果显示,追施不同钾肥在烤烟移栽后的50、65、90 d均对烤烟叶片钾通道基因表达量产生了不同程度的影响,其中T1处理在烤烟移栽后50 d NtNHX2基因和NtKC1基因相对表达量分别为CK处理的1.69、5.15倍,在烤烟移栽后65 d,NtHAK、NtCPK、NtNHX2基因相对表达量分别为CK处理的4.27、1.92、1.67倍,在移栽后90 d,NtCPK基因相对表达量为CK处理的2.15倍;T2处理在烤烟移栽后50 d,NtKC1基因相对表达量为CK处理的3.09倍,在烤烟移栽后65 d,NtHAK、NtCPK基因相对表达量分别为CK处理的2.25、1.73倍,在烤烟移栽后90 d NtCPK、NtKC1基因相对表达量分别为CK处理的1.77、7.98倍;T3处理在烤烟移栽后65 d NtHAK、NtNHX2和NtKC1基因相对表达量分别为CK处理的3.77、3.46、2.56倍,在烤烟移栽后90 d NtKC1基因相对表达量为CK处理的2.15倍。其中NtHAK、NtNHX2基因主要在移栽后65 d起作用,NtCPK基因主要在移栽后65、90 d起作用,NtKC1基因主要在移栽后50、90 d起作用。
对烤后烟常规化学成分的研究结果显示,除T2处理上部叶外,与CK处理相比,追施不同钾肥均提高了烤后烟上、中部叶的钾含量,其中追施黄腐酸钾提钾效果最佳,烤后烟上、中部叶的钾含量分别较CK处理提高了10.00%、33.33%,追施不同钾肥均提高了烤后烟中部叶的钾氯比,其中T3处理烤后烟中部叶钾氯比较CK处理提高了107.41%。
参考文献:
[1]夏 颖,姜存仓,陈 防,等. 棉花钾营养与钾肥施用的研究进展[J]. 华中农业大学学报,2010,29(5):658-663.
[2]王 勇,李廷轩,陈光登,等. 不同钾基因型烟草钾吸收和生理生化特性研究[J]. 中国烟草科学,2017,38(5):56-61.
[3]李荣春,李佛琳,徐琼华. 钾对烤烟叶片解剖结构的效应及其品种差异[J]. 中国烟草科学,2001(2):41-43.
[4]谭 军,蒋美红,李卓璘,等. 钾素对烤烟产质量影响及提钾措施的研究进展[J]. 安徽农学通报,2013,19(6):44-47.
[5]王英锋,徐高强,代卓毅,等. 低钾胁迫下不同钙浓度对烟草钾吸收的影响[J]. 中国烟草科学,2021,42(2):15-21.
[6]巫彬芳. 滴灌施用钾肥对烟草生长和品质的影响[D]. 广州:华南农业大学,2017.
[7]席奇亮. 基于水肥一体化技术的烤烟增产提质效应分析[D]. 郑州:河南农业大学,2018.
[8]江文林. 烤烟中上部叶降碱提钾技术及其生理机制[D]. 福州:福建农林大学,2008.
[9]胡珑瀚. 武隆烤烟钾素及烟碱变化规律与提钾降碱技术的研究[D]. 福州:福建农林大学,2017.
[10]Epstein E,Rains D W,Elzam O E. Resolution of dual mechanisms of potassium absorption by barley roots[J]. Proceedings of the National Academy of Sciences of the United States of America,1963,49(5):684-692.
[11]van Kleeff P J M,Gao J,Mol S,et al. The Arabidopsis GORK K+-channel is phosphorylated by calcium-dependent protein kinase 21(CPK21),which in turn is activated by 14-3-3 proteins[J]. Plant Physiology and Biochemistry,2018,125:219-231.
[12]汤 利,施卫明,王校常. 植物钾吸收转运基因的克隆与作物遗传改良[J]. 植物营养与肥料学报,2001,7(4):467-473.
[13]Gambale F,Uozumi N. Properties of shaker-type potassium channels in higher plants[J]. The Journal of Membrane Biology,2006,210(1):1-19.
[14]Lebaudy A,Véry A A,Sentenac H. K+channel activity in plants:genes,regulations and functions[J]. FEBS Letters,2007,581(12):2357-2366.
[15]王 倩,劉好宝. 烟草重要基因篇:2.烟草钾吸收与转运相关基因[J]. 中国烟草科学,2014,35(2):139-142.
[16]Sano T,Becker D,Ivashikina N,et al. Plant cells must pass a K+threshold to re-enter the cell cycle[J].The Plant Journal,2007,50(3):401-413.
[17]Liu H B. Cloning and expression analysis of potassium channel gene NKT3 from Nicotiana tabacum[J]. African Journal of Biotechnology,2012,11(48):10824-10830.
[18]曲平治,刘贯山,刘好宝,等. 普通烟草K+通道基因NKT4的克隆、序列和表达分析[J]. 植物遗传资源学报,2009,10(3):354-359.
[19]司丛丛,刘贯山,刘好宝,等. 烟草钾离子通道基因NKT5的克隆和序列分析[J]. 中国烟草科学,2010,31(4):8-13.
[20]黄路平,刘 仑,鲁黎明,等. 烟草NtKAT3基因克隆、序列和表达分析[J]. 浙江农业学报,2017,29(7):1057-1063.
[21]卓 维,陈 倩,杨尚谕,等. 烟草K+通道NtSKOR基因的克隆及表达分析[J]. 华北农学报,2018,33(5):99-105.
[22]靳义荣,宋毓峰,白 岩,等. 林烟草钾离子通道基因NKT6的克隆与表达定位分析[J]. 作物学报,2013,39(9):1602-1611.
[23]Hamamoto S,Marui J,Matsuoka K,et al. Characterization of a tobacco TPK-type K+channel as a novel tonoplast K+channel using yeast tonoplasts[J]. The Journal of Biological Chemistry,2008,283 (4):1911-1920.
[24]闫亚飞,王根洪,程廷才,等. 烟草钾离子通道基因NtTPK2的克隆及序列分析[J/OL]. 中国科技论文在线,(2012-03-07)[2023-05-08]. http://www.paper.edu.cn/releasepaper/content/201203-247.
[25]Sato Y,Nanatani K,Hamamoto S,et al. Defining membrane spanning domains and crucial membrane-localized acidic amino acid residues for K+transport of a Kup/HAK/KT-type Escherichia coli potassium transporter[J]. Journal of Biochemistry,2014,155(5):315-323.
[26]Mser P,Thomine S,Schroeder J I,et al. Phylogenetic relationships within cation transporter families of Arabidopsis[J]. Plant Physiology,2001,126(4):1646-1667.
[27]Yang Z F,Gao Q S,Sun C S,et al. Molecular evolution and functional divergence of HAK potassium transporter gene family in rice (Oryza sativa L.)[J]. Journal of Genetics and Genomics,2009,36(3):161-172.
[28]Zhang Z B,Zhang J W,Chen Y J,et al. Genome-wide analysis and identification of HAK potassium transporter gene family in maize (Zea mays L.)[J]. Molecular Biology Reports,2012,39(8):8465-8473.
[29]Guo Z K,Yang Q,Wan X Q,et al. Functional characterization of a potassium transporter gene NrHAK1 in Nicotiana rustica[J]. Journal of Zhejiang University Science B,2008,9(12):944-952.
[30]鲁黎明,杨铁钊. 烟草钾转运体基因NtHAK1的克隆及表达模式分析[J]. 核农学报,2011,25(3):469-476.
[31]谭 颖,秦利君,赵 丹,等. 共转化法获得HAK1基因高表达烟草提高植株钾吸收能力[J]. 植物生理学报,2013,49(7):689-699.
[32]鲁黎明. 烟草钾转运体基因TPK1的电子克隆及生物信息学分析[J]. 中国农业科学,2011,44(1):28-35.
[33]宋毓峰. 林烟草钾转运体基因NsHAK11的克隆与功能分析[D]. 北京:中國农业科学院,2014.
[34]陆玉建,李 震,张弘扬,等. AtHKT1启动子转化本生烟草的初步研究[J]. 华北农学报,2018,33(增刊1):10-16.
[35]马扬扬,陈思原,吴志高,等. 水肥一体化技术对烤烟生育期钾氯变化的影响[J]. 西北农林科技大学学报(自然科学版),2023,51(10):1-11.
[36]王瑞新. 烟草化学[M]. 北京:中国农业出版社,2003.
[37]张 美. 烟草钙依赖型蛋白激酶生化及表达分析[D]. 武汉:武汉大学,2005.
[38]武恒燕. 富钾基因型烤烟的筛选及NHX基因的生物信息学变异分析[D]. 郑州:河南农业大学,2018.
