不同年份下陆地棉和海岛棉杂交F?代配合力及其稳定性分析
2024-05-22赵玉玲邵永杰郭金成曹新川何良荣
赵玉玲 邵永杰 郭金成 曹新川 何良荣

赵玉玲,邵永杰,郭金成,等. 不同年份下陆地棉和海岛棉杂交F1代配合力及其稳定性分析[J]. 江苏农业科学,2024,52(7):56-64.
doi:10.15889/j.issn.1002-1302.2024.07.008
(塔里木大学农学院,新疆阿拉尔 843300)
摘要:对陆海杂交棉F1、F2代在不同年份(2019年、2020年)下各性状配合力比较和遗传稳定性分析,为棉花陆海杂交亲本选择和遗传改良提供理论依据。以6个陆地棉作母本,3个海岛棉作父本,按不完全双列杂交配置18个杂交组合分别于2019年和2020年种植,测定农艺性状、产量和品质性状,对不同基因型生长性状的差异及变异规律进行分析,同时进行配合力和遗传研究,并进行稳定性分析和综合评价,筛选高产、优质、稳定性好的优良基因型。各性状受年份及环境间的影响在2年间难以保持一致;在基因型与环境、年份互作效应分析中,陆海杂交F1代各性状中,基因型的影响最大,其次为基因型×年份和区组,年份对其影响最小;各性状在亲本一般配合力和组合特殊配合力以及配合力与年份互作中,一般配合力方差较特殊配合力方差大,则说明大部分性状主要受加性效应控制,而部分受非加性效应控制或两者共同控制;根据在不同年份下特殊配合力稳定性分析可知,F1组合中惠民6号×新海21和创517×新海21可作稳产组合,J206-5×新海27、J206-5×新海21和创518×新海21可作高产组合。本试验研究得出创518和中641可作为优良亲本,惠民6号×新海27、创518×新海46和创518×新海21可作为高产稳产组合,这为后期优良种质资源的筛选提供了参考。
关键词:陆地棉;海岛棉;配合力;稳定性
中图分类号:S5620.32 文献标志码:A 文章编号:1002-1302(2024)07-0056-09
棉花是我国甚至全世界最重要的经济作物。陆地棉是4个栽培种世界上种植最多的,产量占到棉花总产量的95%以上,其产量高,适应性广,但纤维品质较差,且遗传基础狭窄;海岛棉也称长绒棉,在几个栽培种中品质表现最优,产量占到世界棉花总产量的8%,其具有纤维品质优异、生育期较长、产量稍低、适应范围窄的特点[1]。因此,结合各品种棉花的优质特性,开展优质棉育种工作刻不容缓[2]。
配合力是选择杂种亲本的一项重要依据,可分为一般配合力(GCA)和特殊配合力(SCA)。在不同年份的气候环境因素影响下,使得作物性状产生变异,从而导致GCA和SCA出现年际间的差异,影响性状的稳定性[3]。有研究表明,配合力与基因型、环境互作在水稻、玉米、小麦等方面已有研究[4-6]。目前棉花配合力的分析研究颇多[7-9],但是關于棉花陆海杂交种经济性状GCA和SCA在年份间是否稳定还有待进一步研究。至于棉花陆海杂交种经济性状GCA和SCA与地点互作变异的程度,及其效应值间的相对大小在年份间是否存在显著差异未见相关报道[10]。
本试验采用NCⅡ设计,以3个海岛棉为父本、6个陆地棉为母本于2018年配置18个F1杂交组合,在2019年种植F1代、2020年种植F1、F2代,根据数量遗传学分析模型和方法,对陆海杂交F1代的主要性状作配合力分析,对各亲本不同性状的GCA效应和组合的SCA效应进行分析,比较分析各性状的基因效应,对亲本的遗传潜力和育种价值以及组合的优势利用进行评价。
]1 材料与方法
1.1 试验材料
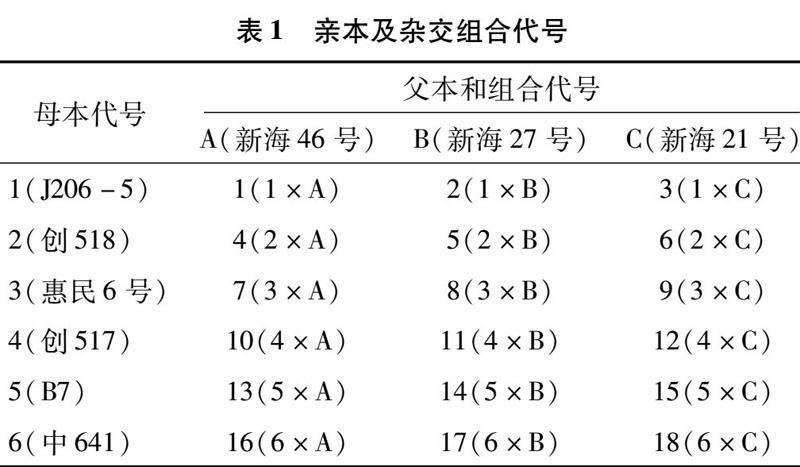
选用9个品种(系)作为亲本,以陆地棉J206-5(1)、创518(2)、惠民6号(3)、创517(4)、B7(5)、中641(6)为母本,以海岛棉新海46号(A)、新海27号(B)、新海21号(C)为父本,采用NCⅡ遗传设计配置18个杂交组合(表1)。
1.2 试验方法
1.2.1 试验设计
田间试验于2019—2020年在第一师阿拉尔市十二团试验基地进行。2019年种植9个亲本及18个F1代杂交组合,收获亲本自交系及F2代,2020年在阿拉尔市十二团种植亲本、F1和F2代,以中棉49作为对照品种。
试验采用随机区组设计。亲本和F1代行长2 m,1个小区2行(1垄);F2代行长2.5 m,1个小区6行(1膜),重复2次,超宽膜模式种植,行距(10+66+10+66+10) cm,株距0.107 m。分别于2019年4月17日和2020年4月11日人工点播,田间铺设滴灌带,试验地连作棉花,肥力条件适中,田间按常规进行管理。
1.2.2 性状调查 于8月中下旬,在每小区中选取具有代表性的10株棉株,测定株高、节数、始果节节位及高度、果枝数、叶枝数、单株铃数、果枝性状[长度和果节数(本研究测定的是单个果枝的果节数)]等农艺性状。于10月中下旬,在棉花枯霜期1周内每小区收获棉株中部棉铃(亲本和F1代收获15个,F2代收获30个),进行室内考种轧花,测定单铃重、衣分等,最终将皮棉样品编号后送至新疆生产建设兵团第一师农业科学研究所棉花质量检测中心,测定纤维品质性状,包括上半部平均长度、整齐度、马克隆值、比强度、伸长率、反射率等。
1.3 不完全双列杂交(NCⅡ)的统计模型
p×q为不完全双列杂交的数学模型[11-14]。其中,母本材料p个,父本材料q个,p和q进行杂交组配,在大田种植r(r=pq)个杂交F1代,每个F1杂交组合小区取n个观察值,则对于任意一个组合内的第k个观察值yijk,其线性模型[15]为:
yijk=μ+gi+gj+sij+eijk。(1)
式中:μ代表总体平均数;gi、gj为母本配合力效应;sij为组合特殊配合力效应;eijk为组合内的个体效应。
p个母本和q个父本皆为固定的研究材料,因此本试验采用固定模型,对于固定模型,约束条件为:
∑[DDi[DD)]gi=0且∑[DDj[DD)]gj=0且∑sij=0。(2)
1.4 数据统计与分析
1.4.1 一般配合力与特殊配合力 因一般配合力是指某一个亲本在杂交后代中的平均表现,其效应[13-14]可按下式计算:
i=xi-x;(3)
j=xj-x。(4)
式中:i表示P1(亲本1)中第i个亲本的GCA效应;j表示P2(亲本2)中第j个亲本的GCA效应;xi表示P1中i个亲本的一般配合力的平均值; xj表示P2中j个亲本的一般配合力的平均值;x表示P1和P2一般配合力的平均值。
特殊配合力(SCA)是指某些特定的组合与其双亲平均表现基础上的预期结果的偏差,因此其效应值可按下式估算:
ij=xij-x-i-j。[JY](5)
利用DPS软件分析供试材料的一般配合力(GCA)和特殊配合力(SCA),然后根据数据进行分析,最终得出相关结论。
1.4.2 遗传力分析 采用随机模型,计算一般配合力方差(Vg)与特殊配合力方差(Vs)[16],公式如下:
Vg=σ2f+σ2m;Vs=σ2fm。(6)
式中:σ2f表示母本方差;σ2m表示父本方差;σ2fm表示母本与父本互作方差。
广义遗传力(H2):
H2=σ2Gσ2G+σ2GEn+σ2εnr×100%。(7)
式中:σ2G表示组合方差;σ2GE表示组合与年份互作方差;σ2ε表示误差项;n表示年份;r表示重复。
2 结果与分析
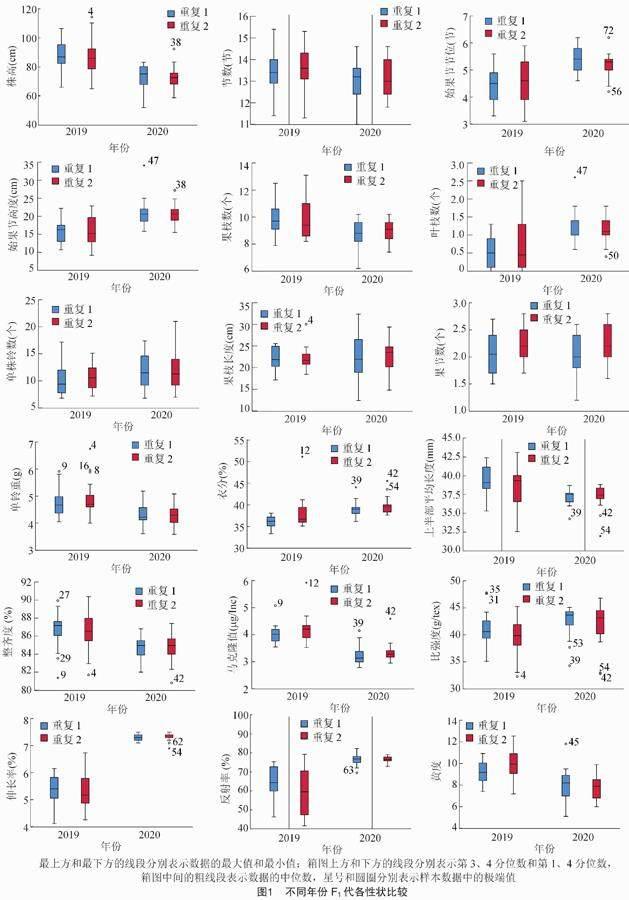
2.1 不同年份F1代各性状比较
对不同年份F1代各组合农艺性状进行比较可知,在不同的年份,F1代18个组合的各性状生长情况并不是完全一致。株高、 果枝数和节数的均值表
现为2019年明显高于2020年,叶枝数、始果节节位和始果节高度的均值表现为2020年高于2019年,而果节数和果枝长度的均值于2019年、2020年没有表现出明显的变化。 由此可以看出,品种组合的各个性状在不同年份下难以保持一致的稳定,F1代各组合的性状也在一定程度上受年份间的影响,导致性状之间出现差异。
2.2 不同年份下基因型联合方差分析
对基因型各性状在不同年份下的联合方差分析结果见表2,各性状区组间衣分达极显著水平,果节数和马克隆值达显著水平;年份间株高、始果节节位、果枝数、叶枝数、单株铃数、衣分、整齐度和伸长率达极显著差异,节数、始果节高度、单铃重、上半部平均长度、马克隆值、比强度和反射率达显著差异,其余极少部分性状表现不显著;说明各个性状受不同环境的影响较大;基因型间单铃重、衣分、上半部平均长度、整齐度、马克隆值、比强度和伸长率均达显著或极显著水平,说明组合间的差异显著;基因与年份互作中始果节节位、叶枝数、单株铃数、单铃重和整齐度达显著或极显著水平,说明各性状受不同年份下的环境影响存在差异,对基因型也产生了影响。
2.3 不同年份下配合力联合方差分析
由表3可知,父本GCA在始果节节位、始果节高度、果枝数、单株铃数、衣分、上半部平均长度和马克隆值存在显著或极显著差异;母本GCA除株高、节数、始果节节位、果枝数、叶枝数、果枝长度、果节数、单株铃数和反射率外,始果节高度、单铃重、衣分、上半部平均长度、整齐度、马克隆值、比强度、伸长率和黄度等9个性状达显著或极显著差异;组合SCA始果节高度、单株铃数、马克隆值、伸长率和反射率存在显著差异;年份×父本GCA互作的节数、果枝长度、单铃重、上半部平均长度、比强度、伸长率和反射率存在显著或极显著差异;年份×母本GCA在果枝数、单铃重、马克隆值、比强度和反射率上存在显著或极显著差异;年份×组合SCA的始果节节位、叶枝数、单株铃数、上半部平均长度、马克隆值和伸长率存在显著或极显著差异。综上,不同性状在不同年份下存在遗传差异。
2.4 不同年份下遗传效应分析
由表4可知,陆海杂交F1代不同性状的遗传力差异较大,节数、始果节节位、叶枝数、单株铃数、单铃重、衣分、马克隆值、伸长率和黄度的遗传力均大于60%,其中衣分、马克隆值和伸长率的遗传力较高,达到99.99%、96.30%和95.23%;其他性状遗传力均低于60%,反射率、果节数和株高的遗传力相对于其他性状较低,为1.50%、8.37%和8.38%。一般配合力(GCA)主要是由亲本品种中基因的加性效应决定的,而特殊配合力(SCA)则主要由亲本品种间基因的非加性效应(包括显性效应和上位性效应)所控制。株高、始果节高度、果枝长度、果节数、比强度和反射率主要受加性效应控制,表现为这些性状的一般配合力方差大于其特殊配合力方差,其余性状则受非加性效应的控制。
2.5 不同年份下亲本产量与品质性状GCA分析
9個陆海亲本在2019年、2020年GCA相对效应值综合分析如图2所示。在株型性状方面,针对株高,亲本1、A和B表现稳定,节数中亲本1、2和6表现稳定,始果节节位中亲本2、6和C表现稳定,始果节高度中亲本2、A和C表现稳定,果枝数中亲本1和B表现稳定,果节数中亲本A表现稳定。在产量性状方面,单株铃数中亲本B表现稳定,单铃重中亲本1、2和3表现稳定,衣分中1、2和C表现稳定。在品质性状方面,上半部平均长度中亲本5、6和B表现稳定, 整齐度中亲本2、6和A表现稳定, 马克隆值相对效应值为负时表现为正向效应,马克隆值中亲本3、4、5、6、A和B表现稳定,比强度中亲本4、5、6和A表现稳定,伸长率中亲本2、4和A表现稳定,黄度中亲本2和C表现稳定。
亲本2除在果枝数、马克隆值、比强度、反射率中2年表现为负向效应,其余大多表现稳定;亲本1在株型性状方面表现较好,亲本1和2在产量性状方面表现较好,亲本2、6、A和B在品质性状方面表现较强优势。
综合可得,亲本2在株型和品质选择上可做优良亲本,亲本2、6和A可在品质选择上作为优良亲本,为选育优良种质资源提供选择,其余亲本在特定的性状中表现出其独有的优势,根据后期育种目标对其进行选择。
2.6 不同年份F1代产量与品质性状稳定性分析
通过作出GGE双标图对F1代18个杂交组合进行稳产性分析,组合在带箭头的直线(环境平均轴)的右边时,其表现大于总体平均值,组合与平均环境轴之间的垂线越长,表示组合越不稳定,反之,垂线越短,组合越稳定。
由图3可知,株高中9个组合高于平均值,较稳定的组合有7和8;节数中8个组合高于平均值,较稳定的组合有13、11和18; 始果节节位中8个组合高于平均值,较稳定的组合有7、12、17和8;始果节高度中9个组合高于平均值,较稳定的组合有4、7和12;果枝数中9个组合高于平均值,较稳定的组合有1和9;叶枝数中11个组合高于平均值,较稳定的组合有15和16;果枝长度中有7个组合高于平均值,较稳定的组合有13和11;果节数中有9个组合高于平均值,较稳定的组合有1、11、2和14。单株铃数中有7个组合高于平均值,较稳定的组合有13;单铃重中有8个组合高于平均值,较稳定的组合有12、8和4;衣分中有7个组合高于平均值,较稳定的组合有1和18。上半部平均长度中有8个组合高于平均值,较稳定的组合有1和5;整齐度中有10个组合高于平均值,较稳定的组合有14和18;马克隆值中有10个组合低于平均值,较稳定的组合有7、10和18;比强度中有10个组合高于平均值,较稳定的组合有15和17;伸长率中有10个组合高于平均值,较稳定的组合有12;反射率中有9个组合高于平均值,较稳定的组合有7;黄度中有7个组合低于平均值,较稳定的组合有14、9和10。
[BT1][STHZ]3 结论与讨论
作物品种的各性状受环境、基因型及两者互作的影响,在不同年份下进行分析时,环境因素对性状差异的影响是首先要考虑的因素,同时也要考虑基因型与环境互作效应[17]。本试验采用2年数据,年际间气候差异较大,国内外学者的相关研究结果[18-20]显示,环境变化是引起性状产生差异的最主要原因,其次是基因与环境互作效应,基因型效应所占比例较低[21]。
[BT2]3.1 基因型与环境互作效应分析
配合力是亲本在后代性状表现中所起作用的度量,是选择亲本的重要依据,也是产生强优势杂交种的遗传基础[22]。研究表明,各性状在年份间、基因型间都存在显著差异;而赖仲铭等研究结果显示,玉米各性状的一般配合力和绝大多数的特殊配合力方差分量在年份间无显著差异,这可能是由作物不同,遗传特性也不同导致的[23]。沈高中等在研究玉米性状结果中表明,加性效应与环境互作是影响玉米产量等性状稳定性的主要因素[24]。
本试验结果表明,区组间3个性状(果节数、衣分和马克隆值)存在差异,但大多数性状表现无差异;年份间8个性状达极显著差异,7个性状达显著差异,其余3个性状(果枝长度、果节数、黄度)差异不显著;基因型中产量和品质9个性状达显著或极显著差异;基因型×年份的叶枝数、单株铃数达极显著水平,始果节节位、单铃重、整齐度、伸长率4个性状达显著水平,说明在陆海杂交F1代中各性状受基因型的影响最大,其次为基因型×年份和年份,区组对其影响最小。
各性状在亲本GCA和组合SCA以及配合力与年份互作中均存在差异性,环境的变化必然会引起基因型的改变,随之也会对配合力的表现产生影响。在不同性状遗传效应中,衣分的遗传力最高,达99.99%,其次为马克隆值(96.30%)、伸长率(95.23%),反射率的遗传力最低(1.50%),GCA主要由基因的加性效应所决定,SCA的遗传基础则是基因的非加性效应,8个性状的GCA方差大于SCA方差,则说明这几个性状主要受加性效应控制。
3.2 配合力在不同年份下稳定性分析
GGE双标图是研究基因型与环境互作及不同环境下作物品种高产稳产的方法[25],其可以将数据以图像的形式展现出来,更直观地反映品种特定的代表性[26-28],通过GGE双标图对参试品种进行综合分析,筛选出丰产性、稳产性好的品种加以利用。罗俊等应用双标图分析甘蔗品种在不同试点的产量和品质性状[29];王启杰等利用双标图对我国油葵主产区12个试点的11个品种进行丰产性和稳产性的综合评价[30];肖继兵等通过双标图分析了30个高粱品种的相关农艺性状和产量[31]。以上均为前人利用GGE双标图对作物品种进行综合分析,筛选得到稳产高产品种。
9个亲本在2年GCA相对效应值中可知,个别性状在部分性状中表现较为优良,2号亲本(创518)除在果枝数上均表现负向效应,其余性状中均为2020年表现优于2019年,2019年单株铃数呈负向效应,其余性状呈现正向效应,6号亲本(中641)在品质方面表现出强优势,综合可知,创518和中641可作为优良亲本進行组配杂交组合,后期根据选育目标进行选择优良的杂种后代。利用GGE双标图对F1代产量与品质性状稳定性分析,结果显示,组合8(惠民6号×新海27)、组合4(创518×新海46)和组合6(创518×新海21)可作为高产稳产组合,组合9(惠民6×新海21)和组合12(创517×新海21)可作稳产组合,组合2(J206-5×新海46)和组合3(J206-5×新海27)可作高产组合。
参考文献:
[1]王巧玲,李 哲. 棉花海陆杂交F1代纤维品质杂种优势及配合力分析[J]. 河南科技学院学报(自然科学版),2013,41(3):12-18.
[2]朱建东. 中、早熟陆地棉与海岛棉杂交种主要性状遗传效应比较分析[D]. 新乡:河南科技学院,2012:32-39.
[3]卢 超,高明博,焦小钟,等. 几个小麦亲本主要农艺性状的配合力评价及遗传力分析[J]. 麦类作物学报,2010,30(6):1023-1028.
[4]何梅洁,陈 佳,李娥贤,等. 青稞产量性状的基因型与环境互作分析[J]. 湖北农业科学,2022,61(7):10-12,19.
[5]王才林,汤玉庚. 杂交粳稻主要经济性状的配合力与年份的互作效应[J]. 江苏农业学报,1989,5(2):1-6.
[6]田笑明. 冬小麦亲本配合力在不同世代和年份间的稳定性研究[J]. 作物学报,1995,21(5):587-591.
[7]朱协飞. 棉花复合杂交育成亲本主要产量性状的杂种优势及配合力分析[J]. 种子,2017,36(10):69-72.
[8]刘 丽,余 渝,王旭文,等. 新疆转Bt基因棉主要性状的配合力和杂种优势分析[J]. 安徽农业科学,2021,49(8):44-48.
[9]崔秀珍,李 哲,常俊香,等. 棉花海陆种间杂交品质性状杂种优势及配合力分析[J]. 湖北农业科学,2008,47(6):647-649.[ZK)]
[10][ZK(#]郭金成,曹新川,宋 佳,等. 不同年份下陆地棉杂交F1产量、品质性状的配合力及稳定性分析[J]. 山東农业科学,2023,55(2):36-43.
[11]杨欢欢,方 晴,聂中欣,等. 番茄优良自交系部分品质性状杂种优势及配合力分析[J]. 东北农业大学学报,2023,54(2):37-48.
[12]孔繁玲. 植物数量遗传学[M]. 北京:中国农业大学出版社,2006.
[13]朱 军. 遗传模型分析方法[M]. 北京:中国农业出版社,1996:61-63.
[14]黄远樟,刘来福. 作物数量遗传学基础:六、配合力:不完全双列杂交[J]. 遗传,1980,2(2):43-46.
[15]刘来福. 作物数量遗传学基础:三、遗传力与选择效果[J]. 遗传,1979,1(5):44-48.
[16]Hanna A D,Kaczmarek Z,Kielczewska H. Genetic analysis of line x tester progenies compared in orthogonally supplemented efficiency balanced incomplete block designs[J]. Biometrical Journal,1989,31(6):753-757.
[17]朴红梅,穆 楠,王延兵,等. 基于AMMI模型的黄淮海夏玉米基因型与环境互作效应对产量性状的影响分析[J]. 北方农业学报,2021,49(4):1-12.
[18]滕振宁,张玉烛,方宝华,等. 用AMMI双标图分析早稻稻米镉含量的基因型与环境互作效应[J]. 生态环境学报,2016,25(4):692-697.
[19]Nowosad K,Bocianowski J,Kianersi F,et al. Analysis of linkage on interaction of main aspects (genotype by environment interaction,stability and genetic parameters) of 1 000 kernels in maize (Zea mays L.)[J]. Agriculture,2023,13(10):1-10.
[20]吴雯雯,欧杨虹. 应用AMMI模型对玉米杂交组合多点试验的稳定性分析[J]. 山东农业科学,2016,48(4):24-27,33.
[21]叶美金,卜俊周,魏建伟,等. 河北省夏玉米品种基因型与环境互作综合分析[J]. 玉米科学,2020,28(1):72-78.
[22]陈亮亮,张 梦,郭立平,等. 陆地棉杂交组合F1、F2苗期优势表现及亲本配合力分析[J]. 新疆农业科学,2023,60(2):261-271.
[23]赖仲铭,杨克诚,郑有良,等. 年份对玉米经济性状配合力及其与地点互作影响的初步研究[J]. 四川农业大学学报,1988,6(2):81-86.
[24]沈高中,赖仲铭. 玉米自交系主要性状的配合力与环境互作的研究[J]. 作物学报,1987,13(1):69-76.
[25]魏鹏程,陈 杜,罗英舰,等. 基于AMMI和GGE双标图的贵州不同生态区春玉米丰产性稳产性和试点辨别力评价[J]. 玉米科学,2023,31(1):22-31.
[26]史跃伟,郎胜勇,王志红,等. 烤烟新品种(系)重要经济性状丰产性和稳定性评价研究[J]. 华北农学报,2013,28(增刊1):238-242.
[27]聂迎彬,穆培源,桑 伟,等. AMMI模型和GGE双标图法在新疆冬小麦区域试验产量分析上的应用[J]. 新疆农业科学,2012,49(9):1569-1575.
[28]闫 雷,邹 莹,沈艳芬,等. 利用不同分析模型评价湖北省马铃薯品种比较试验[J]. 江苏农业科学,2022,50(23):96-101. [HJ2mm]
[29]罗 俊,张 华,邓祖湖,等. 应用GGE双标图分析甘蔗品种(系)的产量和品质性状[J]. 作物学报,2013,39(1):142-152.
[30]王启杰,姜 艳,王 鹏,等. 基于GGE双标图对向日葵品种丰产性和稳定性的综合评价[J]. 干旱区资源与环境,2023,37(1):127-135.
[31]肖继兵,刘 志,孔凡信,等. 基于GGE双标图的高粱品种农艺性状和稳产性分析[J]. 作物杂志,2023(2):36-45.
基金项目:华中农业大学-塔里木大学科研联合基金(编号:HNLH202007);新疆生产建设兵团农业科技创新工程专项(编号:NCG202224)。
作者简介:赵玉玲(2000—),女,甘肃张掖人,从事棉花遗传育种研究。E-mail:1974907991@qq.com。
通信作者:何良荣,博士,教授,从事棉花种质资源与遗传育种研究。E-mail:hlrzky@163.com。
