长期碳酸盐碱度胁迫对脊尾白虾生长及卵巢发育的影响
2024-05-20张秀红李吉涛王佳佳王成伟葛倩倩
张秀红, 李吉涛, 王佳佳, 王成伟, 秦 桢,葛倩倩, 刘 萍, 李 健
(1. 上海海洋大学,水产科学国家级实验教学示范中心,上海 201306;2. 中国水产科学研究院黄海水产研究所,海水养殖生物育种与可持续产出全国重点实验室,青岛海洋科技中心海洋渔业科学与食物产出过程功能实验室,山东 青岛 266071)
盐碱水是分布于陆地区域的非海洋性咸水资源,全世界盐碱地面积为95.4 万km2,仅中国就有9.9 万km2盐碱地和4.6 万km2低洼盐碱水域,广泛分布在西北、东北、华北和长江北岸,包括青海、甘肃、河北、山东等19 个省[1-3]。盐碱水具有高碳酸盐碱度、高pH、水质类型复杂以及主要离子比例失调等特点,导致一般适应性弱的水产动物无法在其中正常生存和繁殖[1,4],因此绝大部分盐碱水还处于闲置状态。长久以来,碳酸盐碱度和盐度被认为是影响盐碱水中水生动物生存、生长和繁殖的主要应激源,研究发现,在碳酸盐碱度超过15.7 mmol/L 的条件下,青鳉(Oryzias latipes)的存活率降低,并发生胚胎凝集、胚胎发育畸形、孵化停止等形态学畸形[5]。房文红等[6]研究发现,中国明对虾(Fenneropenaeuschinensis)幼虾的成活率随着碳酸盐碱度的升高而降低;尤琦等[7]研究发现,盐度和碱度对大银鱼(Protosalanx hyalocranius)的毒性为协同作用。Lin 等[8]发现当碳 酸 盐 碱 度>10 mmol/L,且pH>9.0 时,青 蛤(Cyclina sinensis)的存活率、体长增长率、体重增长率等均显著降低。然而有一部分水生动物由于具有较强的耐盐碱能力,可长期生活在盐碱水域中,如瓦氏雅罗鱼(Leuciscus waleckii)能够适应内蒙古达里湖碱度达53.57 mmol/L、pH 9.6 的恶劣水域条件[9];青海湖裸鲤(Gymnocypris przewalskii)能够通过鳃和肾脏的组织学变化进行渗透和离子调节以适应高盐碱环境[10]。大鳞鲃(Barbus capito)能够通过调节代谢降低氨的产生量,以及在体内将氨代谢合成尿素排出体外的方式应对碱度胁迫时的氨中毒[11]。尼罗罗非鱼(Oreochromis niloticus)可通过鳃离子细胞形态和数量调节适应碱度变化[12]。凡纳滨对虾(Litopenaeus vannamei)在高碳酸盐碱度胁迫下,以增加离子调控的方式进行酸碱平衡的调控[13]。
脊尾白虾(Exopalaemon carinicauda)因具有生长速率快、繁殖周期短、适应能力强等优点,现已成为开展甲壳类盐碱适应性生理机制研究的理想材料。柳飞等[14]研究发现,在低碳酸盐碱度下脊尾白虾生长和繁殖均不受影响,高碳酸盐碱度胁迫下脊尾白虾可以通过调节免疫酶的活性,更好地适应高碳酸盐碱度环境。目前脊尾白虾已成功在滨海盐碱水域河北沧州和山东东营盐碱池塘养殖,且能正常生长和繁殖,并取得了明显的经济效益[15]。关于脊尾白虾耐盐碱的研究大多集中在生长存活方面,对于盐碱水对脊尾白虾繁殖的影响研究较少。因此,本研究设置不同碳酸盐碱度长期胁迫脊尾白虾,旨在探明碳酸盐碱度胁迫对脊尾白虾生长及卵巢发育的影响机理,为开展脊尾白虾在盐碱水中的增养殖提供基础数据和参考信息。
1 材料与方法
1.1 实验材料
实验所用的脊尾白虾来自山东省日照海辰水产有限公司,挑选个体健壮、附肢完整、体表洁净、 规 格 一 致[体 长(2.76±0.20) cm、 体 重(0.12±0.08) g] 的雌虾用于胁迫实验。
1.2 实验设计
实验在200 L 的白色PVC 桶内进行,将同批脊尾白虾雌虾随机分为3 个组,每组3 个重复,每个重复50 尾。以自然海水(盐度25,碳酸盐碱度3 mmol/L)为对照,用1 mol/L 的NaHCO3溶液调整碳酸盐碱度,实验组每天升高1 mmol/L 碳酸盐碱度,直到调整实验用水的碳酸盐碱度分别为5 和8 mmol/L (表1)。胁 迫 期 间 按 体 重 的3%~5%进行饲料投喂,每天投喂2 次(08:00 和18:00),投饵1 h 后进行换水,用虹吸管清除残饵和粪便,每次换水不超过1/3。采用酸碱滴定法测定碳酸盐碱度,以溴甲酚绿-甲基红作为指示剂,整个实验周期为60 d。待实验结束后,统计各组脊尾白虾数量以及卵巢发育情况,然后从实验组和对照组取其肌肉、鳃、肝胰腺和卵巢组织,一部分用组织固定液保存,其余各组织迅速装入1.5 mL 的离心管中放入液氮保存。实验过程中操作人员严格遵守实验动物伦理规范。

表1 各实验组水体盐度和碳酸盐碱度Tab. 1 Salinity and carbonate alkalinity of water in each treatment group
1.3 检测指标与测定方法
实验期间每7 天记录一次脊尾白虾体长和体重,分别从每桶中随机抽取10 尾脊尾白虾,用滤纸吸干虾体表面水分,电子天平称其重量(精确至0.01 g),刻度尺测其体长(精确至0.01 cm)。
待生长实验结束后,统计各组脊尾白虾卵巢发育至不同时期的数量。
式中,L0、Lt分别为实验开始及结束时脊尾白虾的体长(cm);W0、Wt分别为实验开始及结束时脊尾白虾的体重(g);N0、Nt分别为实验初始及结束时脊尾白虾的数量(尾)。
1.4 组织切片制作
用固定好的鳃、肝胰腺和卵巢和组织制备H.E 染色石蜡组织切片。具体步骤:将组织用4%多聚甲醛固定液固定,24 h 后转移至70%的乙醇中保存。制作切片时,将组织进行梯度乙醇(80%乙醇1h,95%乙醇1 h,100%乙醇1 h)脱水。经二甲苯透明[纯乙醇∶二甲苯(体积比1∶1) 1 h,二甲苯1 h] ,并用石蜡渗透[二甲苯∶石蜡(体积比1∶1) 62 °C 1 h,石蜡62 °C 2 h] 、包埋、切片。切片厚度为6 μm,H.E 染色,然后用中性树脂封片,光学显微镜下观察切片组织学变化。
1.5 酶活性测定
碳酸酐酶和Na+/K+-ATP 酶活性采用苏州科铭生物技术有限公司的试剂盒进行测定。准确称取0.1 g 肝胰腺、鳃、肌肉组织,按重量体积比1∶10 加入提取液,进行冰浴匀浆,8 000×g4 °C离心10 min,收集上清液置于冰上,然后按试剂盒说明书进行测定,使用酶标仪进行检测读数。
1.6 数据分析
所得实验数据用平均值±标准差(mean±SD)表示,采用SPSS Statistics 22.0 分析软件进单因素方差分析(One-Way ANOVA),采用Duncan 氏比较组间差异显著性,以P<0.05 为显著水平。
2 结果
2.1 碳酸盐碱度对脊尾白虾生长性能及卵巢发育的影响
生长前期碳酸盐碱度对脊尾白虾体长、体重均无显著影响(P>0.05),42 d 后,5 mmol/L 组体长与对照组差异不显著(P>0.05),但体重与对照组差异显著(P<0.05) (表2,表3)。8 mmol/L 组体长、体重均与对照组差异显著(P<0.05)。5 mmol/L 组GG、WG 和SR 与对照组差异不显著(P>0.05),随水体碳酸盐碱度的上升,即碳酸盐碱度为8 mmol/L 时,脊尾白虾的GG、WG 和SR 均显著低于对照组(P<0.05) (图1)。

图1 碳酸盐碱度胁迫对脊尾白虾生长性能的影响1. 3 mmol/L,2. 5 mmol/L,3. 8 mmol/L。(a)体长增长率;(b)体重增长率;(c)成活率;不同字母表示差异显著(P<0.05),下同。Fig. 1 Effects of carbonate alkalinity stress on growth performance of E. carinicauda 1. 3 mmol/L, 2. 5 mmol/L, 3. 8 mmol/L. (a) growth gain; (b) weight gain; (c) survival rate; different letters mean the difference is significant (P<0.05),the same below.
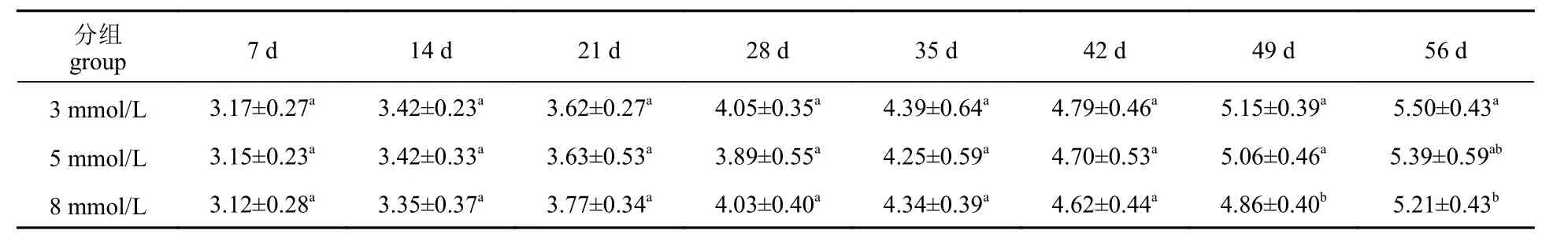
表2 碳酸盐碱度胁迫对脊尾白虾体长的影响Tab. 2 Effect of carbonate alkalinity stress on length of E. carinicauda
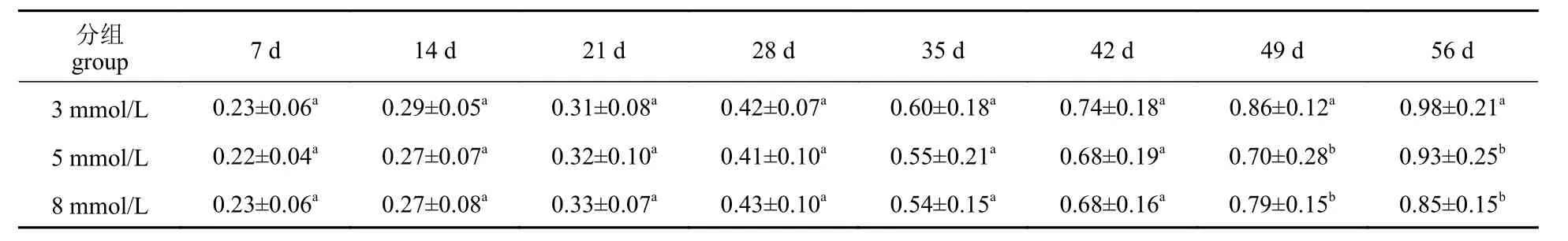
表3 碳酸盐碱度胁迫对脊尾白虾体重的影响Tab. 3 Effect of carbonate alkalinity stress on weight of E. carinicauda
三组脊尾白虾卵巢均能发育,发育至Ⅱ期的百分比分别为20.51%、10.52%和6.25%,其中5 mmol/L 组与对照组间无显著差异(P>0.05),8 mmol/L 组显著低于对照组(P<0.05) (图2-a)。对照组和5 mmol/L 组均有脊尾白虾卵巢发育至Ⅲ期,但两组差异不显著(P>0.05),8 mmol/L 组未见有卵巢发育至Ⅲ期(图2-b)。
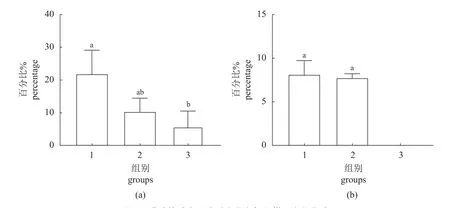
图2 碳酸盐碱度胁迫对脊尾白虾卵巢发育的影响(a)各组卵巢发育至Ⅱ期的脊尾白虾数量百分比;(b)各组卵巢发育至Ⅲ期的脊尾白虾数量百分比。Fig. 2 Effects of carbonate alkalinity stress on ovary development of E. carinicauda(a) percentage of E. carinicauda at stage Ⅱ of ovary development in each group; (b) percentage of white E. carinicauda at stage Ⅲ of ovary development in each group.
2.2 碳酸盐碱度对脊尾白虾组织结构的影响
碳酸盐碱度为3 mmol/L 时,脊尾白虾肝胰腺组织中细胞结构正常且分布均匀,肝小管基膜完整,管腔呈星型,肝细胞结构正常且分布均匀(图版Ⅰ-1)。碳酸盐碱度为5 mmol/L 时,肝小管结构相对正常,管腔呈星型,B 细胞和R 细胞分布均匀,部分B 细胞体积增大,部分转运泡内出现颗粒物质(图版Ⅰ-2)。碳酸盐碱度为8 mmol/L 时,管腔结构严重变形,细胞排列紊乱,R 细胞数量相对较少,B 细胞内部运转泡体积显著降低,转运泡内的颗粒物质明显增加,由于转运泡体积的增大而导致肝小管的形态结构发生变化(图版Ⅰ-3)。
碳酸盐碱度为3 mmol/L 时,脊尾白虾卵巢发育至Ⅰ期,卵原细胞呈卵形,处于增殖状态,细胞核圆形,多数细胞能观察到1 个核仁,少数可观察到2 个核仁,核仁染色最深,卵原细胞排列紧密,周围有单层紧密排列的滤泡细胞(图版Ⅱ-1)。随着碳酸盐碱度的增加,卵巢的发育速率有所减缓,当碳酸盐碱度增加到8 mmol/L 时,卵巢腔内细胞多为卵原细胞,但卵原细胞排列疏松,周围滤泡细胞排列疏松且数量较少(图版Ⅱ-c)。
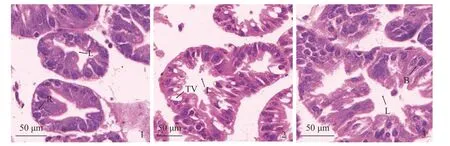
图版 Ⅰ 碳酸盐碱度胁迫对脊尾白虾肝胰腺组织结构的影响1.碳酸盐碱度3 mmol/L (对照组);2.碳酸盐碱度5 mmol/L;3.碳酸盐碱度8 mmol/L,下同。B.分泌细胞,L.管腔,R.储存细胞,TV.转运泡。Plate Ⅰ Effects of carbonate alkalinity stress on the structure of hepatopancreas of E. carinicauda 1. carbonate alkalinity 3 mmol/L (control group); 2. carbonate alkalinity 5 mmol/L; 3. carbonate alkalinity 8 mmol/L, the same below. B. secretory cells,L. lumen, R. storage cells, TV. transport vesicles.
正常脊尾白虾鳃组织为叶状鳃,鳃丝排列整齐、结构完整清晰,呼吸上皮细胞及支柱细胞排列整齐,细胞结构以及次级层片形态结构正常(图版Ⅲ-1)。当碳酸盐碱度为5 mmol/L时,脊尾白虾鳃丝排列紊乱,出现轻微鳃丝肿大现象,角质层略有变形,角质层下间隙扩张变长,上皮细胞与支柱细胞排列紊乱(图版Ⅲ-2)。当碳酸盐碱度为8 mmol/L 时,脊尾白虾鳃丝肿大,排列不规则,出现血细胞肿胀现象,鳃丝上皮细胞破坏,空泡化严重,毛细血管网结构形态改变,角质层下间隙变形(图版Ⅲ-3)。
2.3 碳酸盐碱度对脊尾白虾酶活性的影响
脊尾白虾体内碳酸酐酶活性依次是鳃>肝胰腺>肌肉(图3)。肌肉组织中,5 mmol/L 组的碳酸酐酶活性与对照组差异显著(P<0.05) (图3-a)。碳酸酐酶在肝胰腺组织中差异不显著(P>0.05) (图3-b)。鳃组织中的碳酸酐酶活性随水体碳酸盐碱度的升高整体呈上升趋势,5 mmol/L 和8 mmol/L 组均显著高于3 mmol/L 组(P<0.05) (图3-c)。

图3 碳酸盐碱度胁迫对脊尾白虾碳酸酐酶活性的影响(a)肌肉,(b)肝胰腺,(c)鳃,图4 同。Fig. 3 Effects of carbonate alkalinity stress on carbonic anhydrase activity of E. carinicauda(a) muscle, (b) hepatopancreas, (c) gills, the same as Fig.4.
脊尾白虾体内Na+/K+-ATP 酶活性依次是鳃>肝胰腺>肌肉(图4)。Na+/K+-ATP 酶在肌肉组织中差异不显著(P>0.05) (图4-a)。肝胰腺中,8 mmol/L组的Na+/K+-ATP 酶活性显著低于其他组(P<0.05)(图4-b)。鳃组织中的Na+/K+-ATP 酶活性随水体碳酸盐碱度的升高整体呈上升趋势,其中5 mmol/L和8 mmol/L 组的Na+/K+-ATP 酶活性显著高于3 mmol/L 组(P<0.05) (图4-c),且5 mmol/L组 和8 mmol/L 组之间差异显著(P<0.05)。
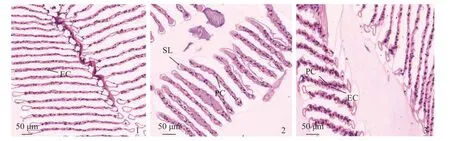
图版 Ⅲ 碳酸盐碱度胁迫对脊尾白虾鳃组织结构的影响EC. 上皮细胞,PC. 支柱细胞,SL. 次级层片。Plate Ⅲ Effects of carbonate alkalinity stress on gill structure of E. carinicauda EC. epithelial cells, PC. pillar cell, SL. secondary slice.

图4 碳酸盐碱度对脊尾白虾Na+/K+-ATP 酶活性的影响Fig. 4 Effects of carbonate alkalinity on Na+/K+-ATPase activity of E. carinicauda
3 讨论
3.1 碳酸盐碱度对脊尾白虾生长性能的影响
增重率、生长率和成活率是衡量水产动物生长和养殖效益的重要指标。研究发现,碱度胁迫会影响尼罗罗非鱼的生长[16]。本研究表明,在碳酸盐碱度胁迫下,脊尾白虾能够生存,但其成活率随着碳酸盐碱度的升高而降低,且在8 mmol/L碳酸盐碱度条件下,其生长速率低于正常组,而5 mmol/L 碳酸盐碱度组与正常组无显著差异,推测高碳酸盐碱度会影响脊尾白虾的生长速率。另一方面,高碳酸盐碱度和高pH 是制约盐碱地水产养殖的主要因子之一[17],且二者对养殖生物生存与生长的影响具有协同作用[18]。研究发现当水环境pH 超过9.0 且碳酸盐碱度为3 mmol/L 时,凡纳滨对虾存活率受到显著影响,随着碳酸盐碱度的升高,存活率降低,且耐受力随着pH 的升高逐渐降低[5]。房文红等[6]研究发现,中国明对虾幼虾的成活率随着碱度的升高而降低,且碳酸盐碱度和pH 对幼虾的致毒效应表现出协同作用。雷衍之等[19]也认为高碱度对水生动物产生的致毒效应受到水体的pH 和盐度等因素的影响。于天基等[20]研究发现,当pH 为5.2~9.9 时,脊尾白虾96 h 几乎没有死亡,这说明脊尾白虾对低pH 具有很高的耐受性,然而随着碳酸盐碱度的升高,水体中的OH-增多,导致pH 升高,因此在高pH条件下脊尾白虾对碳酸盐碱度的耐受性会降低。
鳃作为甲壳动物进行气体交换和离子调节的重要器官,在发挥生理功能时与外界水环境直接接触。研究显示,长期的外界环境因子的变化会引起鳃的结构变化,水体环境对水产动物刺激较大时,会导致鳃的结构出现器质性损伤[21-22],影响甲壳动物的呼吸功能和渗透调节作用。水产动物在对碱度胁迫的适应过程中,鳃的组织结构也发生了不同程度的适应性改变,但当超过机体自身耐受范围,则会发生器质性病变,严重时导致死亡[23]。武鹏飞[24]在研究碳酸盐碱度对达里湖高原鳅(Triplophysa dalai)的鳃组织结构的影响中发现,高碱度会使达里湖高原鳅的鳃丝萎缩、鳃小片弯曲缩短,上皮细胞出现不同程度的水肿,甚至脱落现象。本研究发现,碳酸盐碱度会损伤脊尾白虾的鳃组织结构,脊尾白虾的鳃为叶状鳃,由鳃轴和鳃叶两部分组成[25];在5 mmol/L 碳酸盐碱度胁迫下鳃组织出现轻微肿大、角质层间隙扩张现象,推测脊尾白虾经过一段时间调节后能基本适应此碱度。而当脊尾白虾在8 mmol/L 碳酸盐碱度胁迫下,由于水体中高浓度的OH-、HCO3-、CO32-等复杂离子组成,会直接作用于鳃组织表面,推测各种离子浓度可能超过了脊尾白虾的渗透调节功能,破坏了鳃丝上皮细胞的离子交换体系,使体内酸碱缓冲体系和细胞膜通透性遭到破坏,从而造成脊尾白虾的鳃组织器质性损伤。
肝胰腺是甲壳动物的重要器官,发挥着类似于脊椎动物肝、胰脏和肠道等的功能,其组织结构的变化在一定程度上能反映出机体对环境的适应性改变[26-27]。研究发现,碳酸盐碱度也会导致脊尾白虾肝胰腺组织损伤。研究表明,环境变化可能会引起甲壳动物肝胰腺中细胞组成比例发生适应性变化,特别是B 细胞和R 细胞[28]。三疣梭子蟹(Portunus trituberculatus)在低盐胁迫72 h 后出现了R 细胞减少,B 细胞内转运泡数量增多、体积增大,肝细胞中空泡增多的现象[29]。中华绒螯蟹(Eriocheir sinensis)在氨氮胁迫后也出现了B细胞内空泡增多的现象[28]。这些与本研究的结果具有一定的相似性。B 细胞不仅有分泌功能,还具有消化吸收营养物质的作用。甲壳动物在环境胁迫时,会降低自身活动水平并且将储存的营养物质更多地用于维持机体的基本生理代谢[30]。本研究中,碳酸盐碱度胁迫使脊尾白虾可能需要更多的营养物质来维持基本生理代谢,而B 细胞及其内部转运泡增多、体积增大,促进了代谢吸收,有助于为其提供更多的能量。
此外,碳酸盐碱度还会影响脊尾白虾体内酶的活性,碳酸酐酶是生物体内广泛存在的含Zn金属酶,主要参与CO2的水解过程(CO2+H2O⇆HCO3-+H+),广泛参与细胞内外CO2、HCO3-、H+浓度的调节,与水产动物渗透压调控和酸碱平衡密切相关[31-32]。潘爱军等[33]发现,盐度从5 突变到25 后,凡纳滨对虾鳃和触角腺碳酸酐酶活性显著升高。本研究中,脊尾白虾鳃组织中的碳酸酐酶活性随水体碳酸盐碱度的升高整体呈上升趋势,其中5 mmol/L 组的碳酸酐酶活性与3 mmol/L 组差异不显著,8 mmol/L 组与5 mmol/L和3 mmol/L 组差异均显著,推测在碳酸盐碱度的胁迫下,脊尾白虾通过增强碳酸酐酶的活性来调节体内酸碱平衡与渗透压。渗透调节是广盐类甲壳类动物调节渗透和离子浓度的重要机制,Na+/K+-ATP 酶是渗透压、细胞体积内稳态和维持电化学梯度的关键酶[34],它的主要功能是将胞内的Na+排出,同时向胞内运输K+,在维持细胞膜内外的电化学梯度中起着核心作用[35]。甲壳类动物对离子的吸收主要归因于Na+/K+-ATP 酶的活性[36]。本研究中,随水体碳酸盐碱度的升高,脊尾白虾鳃组织中的Na+/K+-ATP 酶活性整体呈上升趋势,其中5 mmol/L 组和8 mmol/L 组的Na+/K+-ATP 酶活性显著高于3 mmol/L 组,推测在碳酸盐碱度胁迫下,脊尾白虾可以通过增加Na+/K+-ATP 酶活性来调节体内渗透压的稳定。
3.2 碳酸盐碱度对脊尾白虾卵巢发育的影响
水生动物的正常生长繁殖需要一定的碳酸盐碱度来保持水体缓冲能力,在一定范围内调高碱度会提高养殖系统的生产力[37]。有研究表明,在碱度为2.05~4.58 mmol/L 的条件下,蒙古裸腹溞(Moina mongolica)生长发育和生殖的各项指标最佳;碱度为6.43~8.98 mmol/L 时,各项指标都有递降的趋势[38]。徐伟等[11]在大鳞鲃对盐碱的耐受性实验中显示,在盐度3.2、碱度14.32 mmol/L 以下的水体中,胚胎72 h 成活率不受影响;在盐度5.1、碱度14.32 mmol/L 以下的水体,仔鱼96 h 成活率不受影响。本研究结果显示,三组碳酸盐碱度胁迫下脊尾白虾卵巢均能发育,其中5 mmol/L与对照组间无显著差异,8 mmol/L 组显著低于对照组,推测在8 mmol/L 的碳酸盐碱度环境中,脊尾白虾可以生长繁殖,但是其卵巢的发育速率受到了影响。甲壳类动物卵子的发生与滤泡细胞关系密切,滤泡细胞伴随卵母细胞生长的整个过程,随卵母细胞发育程度的不同而相应变化,国内外学者普遍认为滤泡细胞对卵母细胞外源性卵黄物质的积累起着重要作用[39]。通过脊尾白虾卵巢组织学切片发现,5 mmol/L 和对照组卵巢发育几乎同步,卵巢中主要是卵原细胞和单层滤泡细胞,当碳酸盐碱度增加到8 mmol/L 时,卵巢腔内卵原细胞排列疏松,且周围滤泡细胞数量较少,推测在高碳酸盐碱度的影响下,卵母细胞外源性卵黄物质的积累速率减缓,从而影响卵巢的发育速率。综上,脊尾白虾可以在低于8 mmol/L的碳酸盐碱度中生长发育,但其生长速率以及卵巢发育速率仍会受到高碳酸盐碱度的影响。
4 结论
本研究通过不同碳酸盐碱度胁迫脊尾白虾,初步探讨脊尾白虾在长期碳酸盐碱度胁迫下的生长及卵巢发育的情况,研究结果发现,低碳酸盐碱度对脊尾白虾体长增长率、体重增长率以及成活率无显著影响,但高碳酸盐碱度会造成脊尾白虾鳃和肝胰腺组织损伤,导致其体长增长率、体重增长率以及成活率受到影响。通过对脊尾白虾卵巢组织切片观察发现,当碳酸盐碱度增加到8 mmol/L 时,卵巢腔内卵原细胞排列疏松且周围滤泡细胞数量较少,推测在高碳酸盐碱度的影响下,卵母细胞外源性卵黄物质的积累速率减缓,从而影响卵巢的发育速率。
(作者声明本文无实际或潜在的利益冲突)
