气候变化事件对头足类繁殖、年龄生长与早期生活史影响的研究进展
2024-05-20陆化杰陈子越陈新军
陆化杰, 陈子越, 陈新军,2,3,4,5
(1. 上海海洋大学海洋科学学院,上海 201306;2. 上海海洋大学,农业农村部大洋渔业开发重点实验室,上海 201306;3. 上海海洋大学,国家远洋渔业工程技术研究中心,上海 201306;4. 上海海洋大学,大洋渔业资源可持续开发教育部重点实验室,上海 201306;5. 上海海洋大学,农业农村部大洋渔业资源环境科学观测实验站,上海 201306)
近年来,全球频发的气候变化事件给人类生存环境带来了巨大影响,产生了严峻的全球气候变化问题,成为国内外共同关注的焦点[1-2]。联合国粮食及农业组织(Food and Agriculture Organization of the United Nations, FAO)渔业和水产养殖部于2009 年呼吁应当在气候变化应对策略中加入渔业管理计划[3]。海洋是储存气候系统中能量的主要载体,气候变化事件的发生会导致海洋环境因子出现变动,从而间接影响海洋渔业资源的渔业生物学特性,许多生物则会根据不同的气候变化事件呈现出不同的响应[2,4-5]。
头足类是重要的渔业资源,资源丰富,开发潜力巨大[6],是生态系统中扮演 “承上启下” 角色的机会主义者,其生长速率快、生命周期短、生活史过程易受海洋环境因子变动的影响[3,7-8]。为能够充分、有效地开发、利用和管理头足类资源,分析气候变化事件对头足类渔业生物学产生的影响至关重要。本文将对气候变化事件影响头足类渔业生物学的研究现状和焦点内容进行归纳,并对影响其生物学特性的机制进行总结,同时提出后续展望,为今后研究气候变化事件下的头足类渔业生物学特性提供科学依据。
1 主要气候变化事件
气候变化事件是指持续几十年或更长周期的气候状态变化,这种变化可以通过变化率或均值变化进行判断[9]。其中,对渔业资源影响较大的气候变化事件有厄尔尼诺-南方涛动(El Niño-Southern Oscillation, ENSO)[2,10]、拉尼娜(La-Niña)[2,10]、印度洋偶极子(Indian Ocean Dipole, IOD)[11]、大洋暖池/冷池(Warm Pool/Cool Pool)[2,10]、北大西洋涛动/北极涛动/南极涛动(North Atlantic Oscillation/Arctic Oscillation/Antarctic Oscillation, NAO/AO/AAO)[3-4,12-13]、太平洋年代际涛动/大西洋多年代际涛动(Pacific Decadal Oscillation, PDO/ Atlantic multidecadal oscillation, AMO)等[2]。
厄尔尼诺和拉尼娜现象是对渔业资源影响最大的气候变化事件之一。近年来,厄尔尼诺事件出现频率增加,通常表现为厄尔尼诺事件刚结束,随 即 出 现 拉 尼 娜 事 件[2,4,14]。厄 尔 尼 诺 事 件 表现 为赤道太平洋中部和东部海域海表温(sea surface temperature, SST)较正常年份偏高,拉尼娜事件则表现为赤道太平洋中、东部海域SST 较正常年份偏低[2,4,14]。厄尔尼诺事件与南方涛动事件联系紧密,即厄尔尼诺事件发生时,赤道太平洋中、东部与印度洋海面存在的大尺度气压升降震荡现象,二者呈此消彼长的反比关系,故称为ENSO 事件,持续时间范围约为3~7 年[2-3]。
太平洋年代际涛动(PDO)作为一种长周期长寿命(20~30 年1 次)的年代际气候变化事件,反映了太平洋长期的海洋环境背景[15],对太平洋沿岸国家和地区的降水、气温和气候有着重要的影响[4]。PDO 的变化趋势与厄尔尼诺事件相似,分为冷期(负相)和暖期(正相),与ENSO 事件的关系十分密切。PDO 冷期(暖期)时,热带太平洋东部SST 偏低(偏高),西北太平洋SST 偏高(偏低)[2]。厄尔尼诺事件多发生在PDO 暖期,拉尼娜事件多发生在PDO 冷期,在两类气候事件叠加的情况下,厄尔尼诺(拉尼娜)事件在PDO 暖(冷)期发生的频率更高、强度更强[16](图1)。
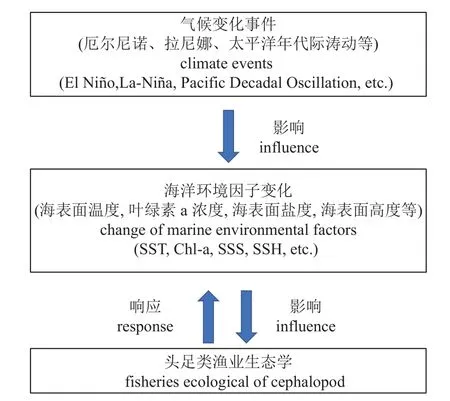
图1 气候变化事件对头足类渔业生态学影响机制Fig. 1 Mechanisms of climate events influencing cephalopod fisheries ecology
2 气候变化事件对头足类个体大小和年龄生长的影响
全球多种气候变化事件(气候和海洋变暖、极端气候事件、极地海冰减少等)往往会推动海洋生态环境的变化,影响海洋生物的生存条件[17]。头足类动物生命周期短,生活史过程可塑性强,对海洋环境变化和气候变化十分敏感,能够迅速地对其做出响应[18-19]。
2.1 气候变化事件对头足类个体大小的影响
头足类受气候变化事件和海洋环境变化的影响在很大程度上取决于海洋环境温度对其个体生长的影响,这也是决定其个体大小的关键因素[18,20-21]。海洋环境温度促使头足类自身新陈代谢速率产生变化,进而导致生长速率变化[18,22]。
研究表明,在较低温度环境下孵化的蛸类生长速率较慢,胴长较大[21];进入到高温环境中生长发育后,其生长速率加快,性成熟后的胴长较小[21],这可能是因为高温环境中的低叶绿素(Chl.a)浓度所致,低水温环境中的生长速率较慢。
气候变化事件引发的海洋环境因子变化与大洋性柔鱼类个体大小变化的关系较为密切。栖息海域温度较低时,茎柔鱼(Dosidicus gigas)的胴长显著增大[23]。1997—1998 年厄尔尼诺事件发生后,加利福尼亚湾海域的茎柔鱼胴长急剧减小,经多个世代后直至2000 年才逐渐恢复正常体型[24-26]。2009—2010 年厄尔尼诺事件发生6 个月以后,加利福尼亚湾海域的茎柔鱼胴长开始显著减小,并维持该体型至2012 年[25,27]。2015 年春季厄尔尼诺事件发生后,该海域茎柔鱼胴长再次骤减,胴体净重较正常年份偏小,并维持小体型直至2018年[25,27-29]。拉尼娜事件发生时,东海太平洋褶柔鱼(Todarodes pacificus)的胴长较正常年份偏小;厄尔尼诺事件发生时,太平洋褶柔鱼的胴长较正常年份偏大[30]。PDO 暖期拉尼娜事件发生时,西北太平洋柔鱼(Ommastrephes bartramii)的胴长较PDO 冷期拉尼娜年份偏大;PDO 冷期厄尔尼诺事件发生时,柔鱼胴长较PDO 暖期厄尔尼诺年份偏大[31]。这表明PDO 冷期时,海洋环境因子对柔鱼个体的生长起限制作用,胴长较小,而PDO 暖期时,海洋环境因子对柔鱼生长起积极作用,胴体生长迅速,胴长较大[31]。拉尼娜年份致使南海中沙海域鸢乌贼(Sthenoteuthis oualaniensis)的胴长较正常年份大,而厄尔尼诺年份胴长较正常年份小[32-34];西沙海域鸢乌贼则为拉尼娜年份胴长较小,厄尔尼诺年份较大[35](表1)。

表1 不同温度下头足类胴体生长情况Tab. 1 Growth condition of body of cephalopods at different temperatures
枪乌贼类的个体大小变化与大洋性柔鱼类颇为相似。海洋环境温度较低时,得益于较高的生产力和充足的食物,莱氏拟乌贼[36]、夜光尾枪乌贼[37]和加州乳光枪乌贼[38]均具有较大体型。枪乌贼(Loligo vulgaris)冬春生群的主要生长期为温度较高的夏秋季,夏秋生群主要的生长期为温度较低的冬春季。季节性上升流的存在使该海域夏秋季的生产力高于冬春季,但经历了食物充沛的夏秋生长季的冬春生群个体依旧小于夏秋群[39-40]。
温度和食物丰度决定了头足类的个体生长和胴体大小[41-44]。在气候事件影响的背景下,海水温度较高的年份,意味着该海域的Chl.a浓度和初级生产力较低,饵料较为缺乏,生活在该海域的头足类胴长相应较小;海水温度较低的年份,海域的Chl.a浓度和初级生产力较高,饵料较为充足,头足类胴体生长情况较好[18,21,38,43]。不同种类头足类的个体生长模式存在一定的差异,但是较高的海洋环境温度致使头足类胴体生长情况较差(低温使胴体生长情况较好)这一规律基本不变,这也表明了头足类在应对气候变化和海洋环境变化中具备高度灵活的生活史策略[18]。
2.2 气候变化事件对头足类日龄与生长的影响
气候变化事件导致的海洋环境温度变化改变了头足类的生长速率,其个体生长策略意味着自身能够对环境或生态系统变化做出快速反应[18]。
海洋环境温度变化对头足类硬组织外形生长和轮纹沉积存在显著影响。海洋环境温度较低时,会导致耳石生长率减缓,日龄轮纹间距变窄,出现较深的标记轮[45]。厄尔尼诺年份,西北印度洋海域鸢乌贼的耳石相对尺寸随胴长增加而减小[46]。随着海洋酸化现象日益严重,海水中pH 值的降低会影响皮氏枪乌贼(Doryteuthis pealeii)的耳石微结构生长,并出现异常结构[47]。同时,SST 较适宜的年份,海洋环境中的Chl.a浓度较高,柔鱼的角质颚、耳石均较低SST 年份偏大[48-49]。PDO冷期,柔鱼角质颚上颚大小随胴体生长而增加;PDO 暖期,角质颚下颚大小在PDO 冷期/暖期发生时均随胴体生长显著增加[50]。气候变暖所引发的海洋酸化现象令海水中pH 值降低,也会导致乌贼的内壳钙化程度降低[51]。
通常在食物获取不受限制的情况下,低温环境中头足类的生长率往往比高温环境的种类低,生命周期较长;而高温水域的头足类生长率较高,生 命 周 期 较 短[18,36,43,52-54]。挪 威 沿 岸 褶 柔 鱼(T.sagittatus)[55-56]和秘鲁沿岸茎柔鱼[23]群体的生长率较暖水域群体慢,平均日龄较暖水域群体大;菱鳍乌贼(Thysanoteuthis rhombus)[57]和安哥拉褶柔鱼(T.angolensis)[58]在稍冷水域中有较慢的生长率;南半球的七星柔鱼(Martialia hyadesi)[59]和阿根廷滑柔鱼(Illex argentinus)[60]的春季孵化群体较冬季孵化群体的生长速率快。不同海域头足类种群的生长策略随水温高低变动,这说明海洋温度上升对头足类生长率提升的影响较大。亚热带海域的莱氏拟乌贼生长速率较热带海域慢[36],夜光尾枪乌贼[37]和枪乌贼[61]等在较冷海域均拥有较慢的胴体生长速率。温度较低的海水促使莱氏拟乌贼[36]、夜光尾枪乌贼[37]、茎柔鱼[23]、褶柔鱼[55-56]、太平洋褶柔鱼[62]和科氏滑柔鱼(I.coindetii)[63]的性腺发育成熟延迟,生命周期也得到一定延长。生活在极地和深海海域的头足类会限制其自身蛋白质的合成,可能会导致其胚胎发育和生长过程发生延迟[64]。但也有研究表明,拉尼娜事件发生时,经历了较低温度的加州乳光枪乌贼个体生长率较厄尔尼诺年间快,这可能是由于拉尼娜年间水温较低,海域较高的生产力能够提供充沛的食物所致[38]。夜光尾枪乌贼适宜栖息在亚热带和温带海域,进入热带海域后生长速率反而较亚热带和温带海域下降,但其总体生长速率的变化规律还是随纬度增加而降低,生命周期得到延长。这种情况可能是由于头足类的代谢速率被限制在一定的温度范围内,栖息海域温度过高可能会使代谢速率高于食物消化速率,胴体生长反而受到抑制[37,53-54]。常年生活在12 °C 海水中的球形深海多足蛸(Bathypolypus sponsalis)的日体重生长率(growth rate, GR)为2%[64-65],7 °C 的 深 海 多 足 蛸(B.arcticus) GR 为0.4%[64],0 °C 的沙氏近爱尔斗蛸(Pareledone charcoti) GR 为0.1%[64,66],而 生 活 于 热 带 的 周 氏 蛸(Octopus joubini) GR 为5%[64],远远大于冷水域蛸类(表2)。

表2 不同温度下头足类胴体生长率Tab. 2 Growth rate of body of cephalopods at different temperatures
Forsythe[22]通过模型预测认为,即使头足类在孵化后前3 个月中所经历的海洋环境温度变化存在很小的差异,也会对整个生活史后期的生长特性造成很大的影响。特定年份中,孵化时间较晚但同时经历较高环境温度的幼体生长速率更快[22]。孵化于低温且生产力较高海域的头足类,随着水温的升高,胴体生长速率逐渐加快[21]。生活史早期阶段经历一定时间范围高温环境可能会加快头足类的胴体生长速率并达到更大体型[22,53,67]。但是,并不是较高的海洋环境温度一定能促使头足类的生长速率都得到加快,因为过高的温度同样会导致海洋环境中的生产力下降和胴体生长能力下降,从而降低其胴体生长速率[18,68]。这说明了在气候事件发生的背景下,影响头足类成熟阶段的个体大小主要因素为气候事件发生所带来的海洋温度变化,以及遗传和食物供给等多因素的共同作用[64]。
3 气候变化事件对头足类繁殖特性的影响
气候变化事件导致的海洋环境因子季节性变化对头足类的繁殖情况变动存在着深远的影响,其变化会导致头足类生殖能量分配和生殖投入的时间和速率存在差异[54,69-71]。其中,环境中温度和食物量等因子对头足类不同生活史阶段的繁殖力和性腺发育的影响较为明显,可推动性腺提前或推迟成熟,影响成体交配活动,对头足类的繁殖策略产生重大影响[22,72]。
3.1 气候变化事件对头足类性成熟度的影响
研究表明,挪威沿岸的褶柔鱼群体较西非沿岸群体性成熟推迟[56]。同生活在温带水域的柔鱼秋生群性成熟速率较冬生群快[73]。较高的SST 同样会使得太平洋褶柔鱼[62]和科氏滑柔鱼[63]的生命周期缩短,性成熟体型变小,性成熟提前。1997—1998 年厄尔尼诺期间,茎柔鱼的性成熟胴长较小,性成熟阶段提前[63]。经历1999 年拉尼娜的低水温以后,茎柔鱼性成熟阶段推迟;2009—2010 年厄尔尼诺期间,日龄为6 个月的茎柔鱼以较小的胴长提前达到性成熟阶段[25,74]。2015 厄尔尼诺年间,性腺发育成熟的茎柔鱼仍为小个体表现型,直至2018 年才恢复成正常性成熟体型[27-28]。
较高的海水温度同样会导致夜光尾枪乌贼[37,54]、枪乌贼[75-76]、莱氏拟乌贼[77]和澳大利亚拟乌贼(Sepioteuthis australis)[78]的生命周期缩短,性成熟体型变小,性成熟提前。温带水域的夜光尾枪乌贼,秋季群体的性腺发育成熟阶段的速率最快,冬季群体的性腺发育阶段成熟最迟[37]。这说明气候事件引发的海洋温度逐渐升高,将会促使头足类的性成熟提前,性成熟体型变小,世代更替速率加快[54](表3)。
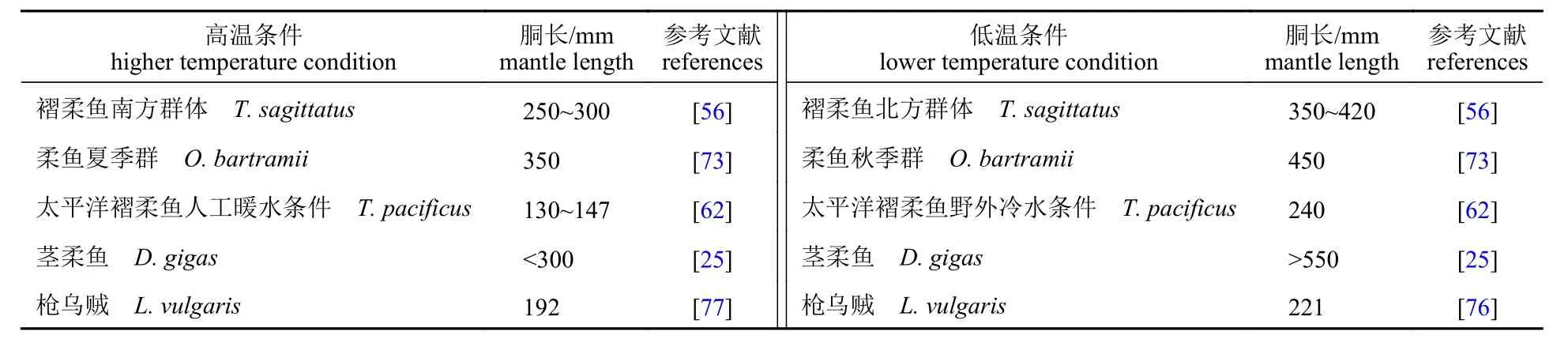
表3 不同温度下头足类性成熟度Tab. 3 Sexual maturity of cephalopods at different temperatures
3.2 气候变化事件对头足类繁殖力的影响
水温变化对头足类的繁殖力变化存在显著影响,气候异常年份的头足类个体较气候正常年份存在差异,个体大小变化导致其繁殖力也存在显著差异。饲养水温为24 °C 时,参与产卵活动的雌性玛雅蛸(O.maya)个体较多,当水温升至31 °C时,仅13%的个体参与产卵活动,且怀卵量明显减少[79]。厄尔尼诺年间茎柔鱼的繁殖力显著下降,冷水域大型茎柔鱼的繁殖力是暖水域小型个体的8~9 倍[23,28,80]。通 常 认 为,低 水 温 及 食 物 充 足 条件下头足类具备更高的繁殖力,高水温及食物匮乏时的繁殖力较低[25,81],间接说明除温度以外,食物丰富度对头足类繁殖力的影响也不容小觑[25]。
3.3 气候变化事件对头足类生殖投入的影响
性腺发育指数(gonad-somatic index, GSI)结果表明,暖水域枪乌贼的相对生殖投入水平较冷水域的高[75];温暖月份孵化的雄性澳大利亚拟乌贼相对生殖投入水平较高[78]。一般情况下,暖水域头足类的生殖投入水平较高,生殖系统的重量占体重的比例也更高,但暖水群个体较小且性腺重量更轻,胴体的大小限制了性腺组织的发育[36,54,69]。因此,即使暖水群体的相对生殖投入比冷水群多,但冷水群体的绝对生殖投入水平更高(即生殖系统的总重量)[36]。小个体头足类的绝对生殖投入较少,需要的食物量也较少,在食物相对匮乏时能摄取数量更多、体型更小的食物维持生长和繁殖[43,82]。雄性头足类的生殖投入水平比雌性小,有更多的精力快速响应海洋环境变化[38,82]。春夏季孵化的雌性澳大利亚拟乌贼的绝对生殖投入水平较秋冬季个体低[78]。春夏季的低绝对生殖投入水平使其排出的卵子体积小且数量多,而秋冬季雌性排出的卵子体积大但数量少[78]。这说明在全球变暖的背景下,随着海水温度的升高,头足类动物的相对生殖投入将会增加,绝对生殖投入相对减少,低温状态下则相反[78]。
4 气候变化事件对头足类早期生活史特性的影响
4.1 气候变化事件对头足类卵期的影响
全球变暖的趋势下,升高的海水温度对头足类的卵粒受精和卵黄发育均有负面影响[79],卵内的卵黄体积与孵化时的外界海水温度呈负相关[83],胚胎在较低温度下拥有更多卵黄[79,84]。海洋环境温度能够直接影响卵黄转换和胚胎发育的持续时间,进而影响孵化个体的大小[54,85],低温下孵化的枪乌贼胚胎体重和胴长明显较高温下孵化的胚胎高8%和7%[85]。低水温通常会导致胚胎在孵化前的发育时间更长,意味着能够孵化出体型相对较大的仔鱼,具备更好的游泳和捕获猎物的能力[54,85],提升了幼体的存活率[86-87]。升高的海水温度加快了卵内胚胎的代谢率,卵黄内的营养也在发育过程中消耗得更快、更早,提早孵化的仔鱼发育程度较差,存在较大的概率出现个体发育异常和畸形现象[88]。
4.2 气候变化事件对头足类幼体大小和生长的影响
头足类幼体生长易受到气候变化事件和海洋环境影响[38],幼体大小和幼体生长速率与海洋环境温度变化密切相关[22,84]。虽然较高的水温会导致孵化个体较小,但对头足类的新陈代谢和幼体生长速率也存在促进作用[89-90]。较晚孵化但历经一段较高环境温度的幼体,其幼体胴体生长优于较早孵化的个体[22]。厄尔尼诺年间,乳光枪乌贼幼体的个体较拉尼娜年间更大,且该气候变化事件对幼体生长的影响主要集中在仔鱼期后期[86,91]。水温的升高将会缩短乳光枪乌贼(D.opalescens)[92-93]、枪乌贼[88]和巴塔哥尼亚枪乌贼(D.gahi)[94]的孵化期,仔鱼生长速率加快;温暖月份出生的枪乌贼[40]和巴塔哥尼亚枪乌贼[95]幼体生长速率较冬生群快,仔、稚鱼的体型更大。生活史早期经历高水温的巴塔哥尼亚枪乌贼,较冷水中的个体拥有更大的体型[67,93]。
胚胎的发育高度依赖海洋环境温度,但发育过程中会存在适宜温度的上限和下限[89],过高或过低的水温均不利于头足类胚胎的发育[86]。水温范围为13~17 °C 时,枪乌贼胚胎成活率为92%~96%,水温升至19 °C 时,成活率急剧下降至47%[96]。逐渐上升的海水温度同样使夜光尾枪乌贼的胚胎期缩短,孵化幼体的个体更小[54,97]。厄尔尼诺事件的发生使柔鱼冬春生西部群体的孵化成活率下降[98],水温低于16 °C 时,柔鱼胚胎器官发育出现异常,水温为22~24 °C 时,胚胎发育恢复正常[99]。
秋生群柔鱼的幼体比冬生群生长更快[70],并且随着厄尔尼诺导致的温跃层加深,柔鱼仔鱼将会在寻找食物的过程中投入更多的能量[97]。栖息于较高水温的阿根廷滑柔鱼幼体,体型较低水温中栖息的个体更大[60]。相比于冬生群的仔鱼,出生于高水温海域的夏生群仔鱼通常更能够适应气候变暖[88]。但是,低温月份意味着较高的生产力和丰富的食物,这时往往会有较多的成体同时繁殖,孵化出更多的幼体以匹配有利的环境条件[93]。通常情况下,头足类胚胎生长速率随海水温度的升高而显著增加[96],卵内胚胎在较高的温度下发育速率较快,孵化出的仔鱼体型也更小[18]。在未来全球变暖趋势下,头足类早期生活史特性最显著的改变便是幼体提早孵化和体型偏小,出现畸形个体的概率增加[88,96]。
4.3 气候变化事件对头足类早期生活史的其他影响
全球变暖和海洋酸化加剧导致的高CO2浓度和低pH 环境使头足类的胚胎期延长,生长发育变得迟缓[86,100]。CO2浓度升高的情况下,皮氏枪乌贼的胚胎孵化期延长,仔鱼的耗氧率显著降低[47],胚胎发育出现了较高比例的异常和畸形现象[96]。13~14 °C 时,胚胎成活率为92%~94%;当温度高于这个区间时,胚胎成活率明显下降至71%[92]。与此同时,海水中较低的pH 值(高CO2浓度)也会使得幼体具有更高的耐高温极限[96]。
气候事件背景下,头足类的几个关键生活史阶段均受到海洋环境温度变化带来的不同程度的影响[99-101],温度控制着胚胎发育期和成熟期的长短,对延长生命周期和早期生活史变化的影响较为明显[64]。头足类动物整个生活史中对环境变化最敏感的阶段是仔鱼期(paralarval stage)[102-103],该阶段死亡率较高[99,104],个体生长和发育与海水温度变动密切相关[99,105-106]。伴随着日益加剧的全球海洋变暖,卵内非生物压力条件将会对头足类胚胎的存活率和生长产生有害的影响[88]。全球海洋变暖还会增加头足类仔鱼的食物需求量,但高CO2浓度可能会导致生物体代谢率处于较低的水平,导致蛋白质合成减少,进而影响个体生长[96]。
较低的海洋环境温度通常意味着深层海水上升,营养和氧气均较为充沛,初级生产力较高[92]。高营养水平和低温会导致头足类性成熟推迟,而低营养水平和高温会导致性成熟提前[41,43]。因此,高水温且食物匮乏条件下新孵化的仔鱼存活时间十分有限,高代谢率也促使仔鱼需要更多的食物,但在死亡之前能找到食物的所能分配的时间也更少[88]。相比之下,低水温条件下孵化的仔鱼捕食成功率较高,也更加容易获取食物,能够摄食的时间也更加充沛[92]。除温度外,盐度同样也是影响头足类仔鱼成活率的重要影响因素之一[62]。头足类动物在各生活史阶段经历的海洋环境温度和营养状况决定了其生长和性成熟状态,而这二者又控制着个体所能够达到的最大体型,进而影响下一代的孵化个体[43]。
5 总结与展望
5.1 气候变化事件对头足类个体大小和年龄生长的影响
头足类分布范围广,其生物学特性易受到气候变化事件引发海洋环境因子变动的影响[3,7-8]。本文通过归纳和总结当前国内外相关领域的研究成果,分析了作为当前热点的气候变化事件对头足类渔业生物学特性可能带来的影响。其中,气候变化事件引发的海洋环境温度变化是影响头足类个体大小和年龄生长的关键因素[18,20-21]。一般情况下,较高的水温意味着低初级生产力,饵料较匮乏,生活于其中的头足类个体较小,生长速率较快,生命周期较短;较低水温下则反之[18,36,43,52-53]。同时,头足类的代谢速率被限制在一定温度范围之内,持续过高的水温并不会一直提高头足类的胴体生长率,反而会出现代谢速率过高的现象,导致胴体生长情况欠佳[53-54]。综上所述,不同气候变化事件背景下,影响头足类个体大小和年龄生长特性的重要因素是海洋环境温度和栖息海域饵料丰度。
5.2 气候变化事件对头足类繁殖特性的影响
海洋环境因子的剧烈变化会影响头足类性腺发育、繁殖力和生殖能量分配及投入,改变繁殖策略[22,54,69]。全球范围内,逐步升高的海水温度将会促使头足类的性腺发育加快,性成熟提前,性成熟体型变小,繁殖力下降,世代更替速率加快,水温较低时则反之[36,54]。生殖投入方面,暖水条件下的头足类群体生殖系统占体重的比例大于冷水域群体,其相对生殖投入较大;但是冷水条件中的头足类个体较大,性腺的绝对重量大于暖水域群体,绝对生殖投入较大[36,54,69]。为应对气候事件的发生,头足类的繁殖策略根据海洋环境的变化作出响应,这也充分说明了解气候变化事件背景下头足类的繁殖特性变化是研究其渔业生态学特性和生活史的关键点之一,但是目前大多数相关的研究还停留在基础生物学的表象,结合海洋环境因子深入探究影响程度的研究较少。
5.3 气候变化事件对头足类早期生活史特性的影响
头足类的胚胎发育、孵化大小和幼体生长速率易受到海洋环境温度变化的影响[22,38,85],过低或过高的海水温度均会影响胚胎发育和孵化的成功率[96,98-99]。当今全球变暖的趋势下,头足类胚胎孵化期出现逐渐缩短的现象,孵化的个体体型较小,幼体生长速率加快[96]。与此同时,较高的水温使卵内卵黄消耗速率加快,胚胎的成活率降低,出现畸形个体的概率增加[79,83-87]。全球海洋酸化的出现,加剧了胚胎发育速率变缓程度,影响了幼体生长发育[96]。除气候变化事件改变海洋环境温度以外,饵料丰度变化也是改变头足类繁殖策略和早期生活史特性的主要影响因子[43,83,93,102-103]。
5.4 展望
气候变化事件现已成为全球学界关注的焦点。头足类动物作为海洋环境中举足轻重的一环,同时也是人类重要的渔业资源,因此研究气候变化事件对头足类动物的影响逐渐被重视。通常情况下,气候变化事件影响并不直接作用于头足类动物本身,而是通过引起海洋环境变动来引发海洋初级生产力变动,进而影响头足类的渔业生物学特性。与此同时,各方研究涉及的头足类种类存在一定偏向性,研究对象多为常见经济种类;研究过程中往往也只考虑到海洋环境温度的变化,并未涉及其他的环境因子,涉及气候变化事件影响头足类的生物学和生态学研究也较为稀缺。大洋性柔鱼类和枪乌贼类由于特殊的栖息环境和生活习性,幼体的采集难度较大,当前主要通过采集渔获物样本进行研究,相比近海的蛸类和乌贼类不易开展人工饲养实验。今后研究中应扩大采样海域,改进采样方式,结合浮游生物采样,有针对性地对潜在或已知产卵场海域进行调查,尽可能地获取早期生活史阶段样本,以充分了解早期生活史特性。同时,应注重对海洋环境数据的收集(大多为温度这一环境因子,盐度、季风、海水酸度、混合层深度等环境因子应用较少),同时补充和结合多年间海洋环境数据,推动气候变化事件影响头足类动物的研究进展,从而指导头足类渔业生产活动。
(作者声明本文无实际或潜在的利益冲突)
