中国鱚科鱼类分类研究进展
2024-05-18高天翔肖家光
高天翔,李 珂,肖家光
(1.浙江海洋大学水产学院,浙江舟山 316022;2.自然资源部第三海洋研究所,福建厦门 361005)
鱚科(Sillaginidae,Richardson 1846)隶属于硬骨鱼纲Osteichthyes、鲈形目Perciformes、鲈亚目Percoidei,广泛分布于印度洋和西太平洋热带、亚热带、温带海域,常见于浅滩、河口,淡水水域鲜有发现,栖息水深较浅[1-4]。
鱚科鱼类体延长,略呈圆柱状;头前部平坦,口小,吻钝尖,眼位于体两侧;鱼体背侧体色较深,腹侧体色浅。背鳍2 个,第一背鳍具10~13 硬棘,第二背鳍由1 个硬棘及16~27 个软条组成;腹鳍胸位,具有一丝状延长,由1 硬棘及5 条软条组成;臀鳍起始端具2 硬棘(第一硬棘一般较小)及14~26 条软条组成;胸鳍中大;尾鳍截形或浅凹。左右鳃盖膜相连,跨越颊部;鳃耙短钝,幽门盲囊少;鱼鳔有的退化或发育不全,有的结构复杂,具有多变的分支结构;脊椎骨数32~44[1-6]。
鱚科鱼类具钻沙习性,肉质鲜美、营养丰富,经济价值较高,深受人们喜爱,是近海渔业捕捞对象和游钓鱼种。我国沿海均有鱚科鱼类分布,近年也开始了多鳞鱚人工繁育[7-9]。
1 鱚科鱼类分类学研究
鱚科鱼类种类鉴定主要依据可数性状、可量性状相对比例、体色斑纹和鱼鳔等特征。1846 年RICHARDSON 命名鱚科Sillaginidae,1992 年,MCKAY 描述了3 属31 种鱚科鱼类,鱚属记录有29 种:大指鱚亚属Sillaginopodys 1 种、鱚亚属Sillago 4 种和副鱚亚属Parasillago 24 种[1]。
近年来,国内外陆续有一些鱚属新种发布。2010 年,KAGA,et al[10]在阿曼沿海发现一鱚属新种——Sillago caudicula,该新种后来在非洲南部马达加斯加岛沿岸也被发现。2011 年以色列学者GOLANI,et al[11]重新描述了Sillago erythraea,并确定了其种的地位。他还指出Sillago sihama 在红海的分布范围仅限于红海南部水域。然而,有关Sillago sihama 和Sillago erythraea 的关系一直饱受争议,一些学者[5]不认同Sillago erythraea 新种地位。随后TIKOCHINSKI,et al[12]从基因水平再次证明其新种地位,并于2014 年发布其最终命名——Sillago suezensis。2017 年PANHWAR,et al[13]在巴基斯坦近岸发现一鱚属新种Sillago panhwari。通过对印度鱚Sillago indica 鱼鳔的研究,2012 年KAGA,et al[14]指出印度鱚应隶属于鱚亚属,而非副鱚亚属。
国内学者关于鱚科鱼类分类相关的研究很少。1986 年,SHAO Kwang-tsao,et al[15]对台湾地区鱚科鱼类进行了重新整理,并报道了一个新纪录种Sillago chondropus,认为台湾近海存在1 属3 亚属8 种,但台湾鱼类资料库中记载在台湾高雄采集到纤鱚Sillago ciliata,其准确性有待考证[6,16]。通过形态特征和DNA 条形码分析,GAO Tianxiang,et al[17]发现了分布于莱州湾到台湾海峡的鱚属新种—中国鱚Sillago sinica;2013年,韩国学者BAE S E,et al[18]也发现在光阳近海存在中国鱚,并纠正了KWUN H J,et al[19]将该种鉴定为小鳞鱚的错误。2016 年,XIAO Jiaguang,et al[20]在台湾海峡近岸水域发现一鱚属新种—邵氏鱚Sillago shaoi;2021 年,XIAO Jiaguang,et al[21]采集台湾海峡到海南的12 个群体459 尾样品,根据体侧具明显黑色条带特征,命名了新种鱚鱼—黑带鱚Sillago nigrofasciata;2022 年,YU Zhengsen,et al[22]对采自湛江和北海的127尾样品开展了形态特征和COI 条形码序列分析,命名了鱚属鱼类新种—拟多鳞鱚Sillago parasihama。
目前,已知鱚科鱼类约38 种,我国沿海有14 种已命名的鱚科鱼类,见表1[1-4,6,9,16-17,20-25]。
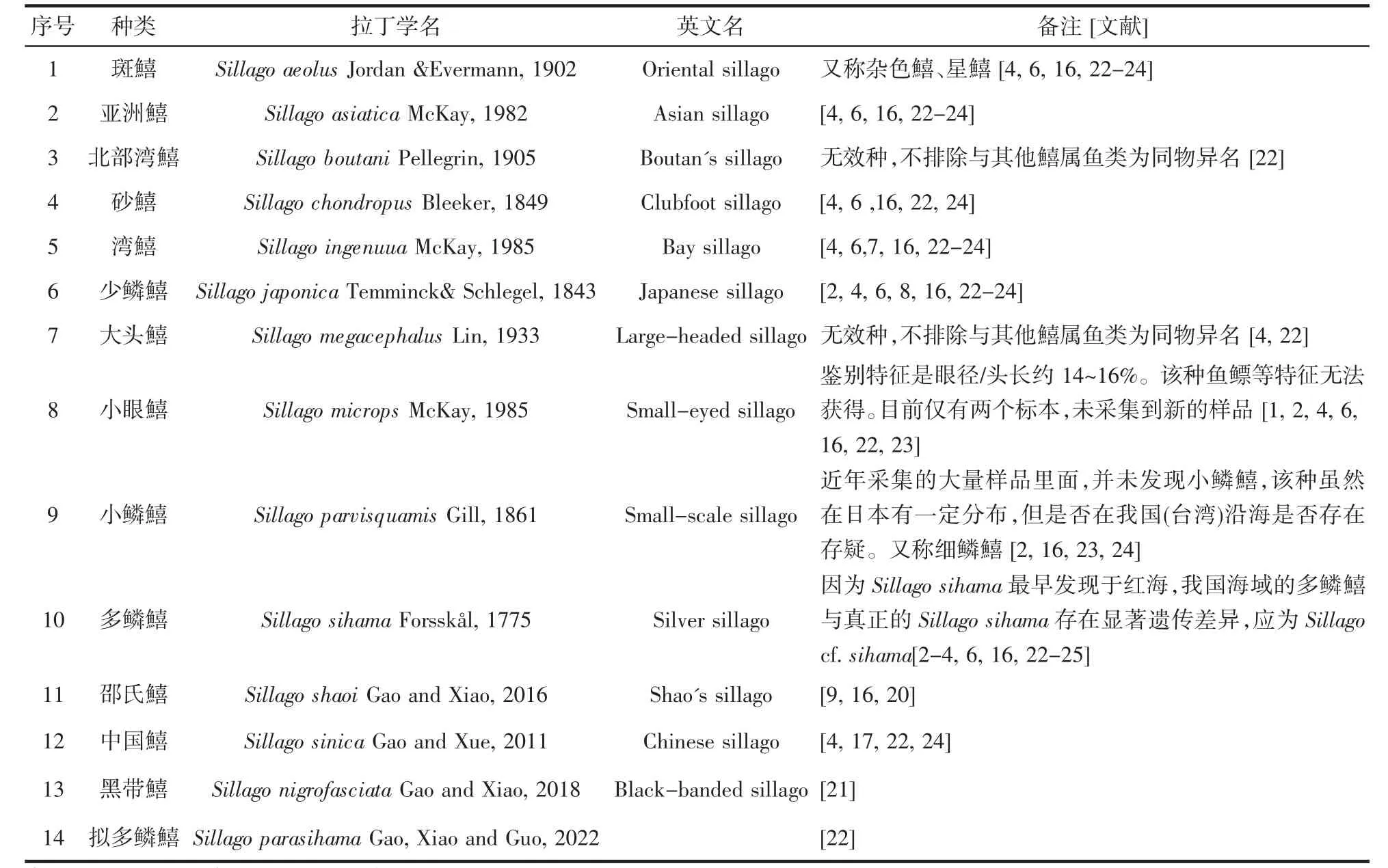
表1 中国鱚科鱼类Tab.1 Sillaginidaespecies in China
2 鱚科鱼类系统发育研究
自1846 年RICHARDSON[26]命名鱚科Sillaginidae 以来,鱼类学家们在鱚鱼的系统发育研究问题上一直存在争议。2004 年,SPRINGER,et al[27]通过鳃弓的背侧肌肉和相关的骨骼研究认为鱚科鱼类与鲈科及溪鳢科鱼类亲缘关系较近,而BETANCUR-R,et al[28]和NEAR,et al[29]基于分子证据的研究结果显示鱚科与美花鲈科Callanthiidae 和松鲷科Lobotidae 关系较近[5]。
1861 年,GILL[30]认为鱚科只有鱚属Sillago Cuvier 和拟鱚属Sillaginopsis Gill;随后,GILL[31]于1862 年又新增似鱚属Sillaginodes。1874 年,BLEEKER[32]的研究也支持GILL 将鱚科分为3 属的观点。但BOULENGER[33]于1901 的研究中仅认可Sillago 一属。鱚科鱼类曾被分为不同的亚科。1933 年,FOWLER将鱚科划分为Sillaginae 亚科(包括似鱚属Sillaginodes 和鱚属Sillago) 和包括拟鱚属Sillaginopsis 的Sillaginopsinae 亚科,Sillago 属分成Sillago 和Sillaginopodys 两个亚属[34],但MCKAY、NELSON 等学者不认可FOWLER 关于亚科的划分[1,3,35]。
1985 年,MCKAY[35]描述了似鱚属Sillaginodes、拟鱚属Sillaginopsis 和鱚属Sillago 3 属25 种鱚科鱼类,其中,似鱚属1 种,拟鱚属1 种,并基于鱼鳔形态特征,将鱚属划分为3 个亚属:大指鱚亚属Sillaginopodys Fowler,1933 (鱼鳔退化);鱚亚属Sillago Cuvier,1817 (鱼鳔具2 个后部延伸);副鱚亚属Parasillago McKay,1985 (鱼鳔具1 个后部延伸和管状结构)。鱚属、拟鱚属、似鱚属曾一度被认为是鱚科鱼类有效分类单位[1,28]。然而,NELSON,et al[3]不支持MCKAY 的亚属观点。2013 年,KAGA[5]在形态特征分析和比较解剖学研究的基础上,将鱚科划分为鱚属、拟鱚属、似鱚属、大指鱚属Sillaginopodys 和新建立的新属Sillaginops 等5 个属,见图1。

图1 MCKAY(1992)与KAGA(2013)鱚科分类结果比较(数字分别表示鱚科各属的鱼种数)Fig.1 Comparison of the Sillaginidae classification between McKay (1992) and Kaga (2013) (numbers indicate the number of fish species respectively)
目前,FISHBASE[25]采用了KAGA 鱚科5 属观点,但CHENG Jie,et al[36]基于多基因联合分析的分子系统发育研究结果与其并不一致。
3 我国鱚科鱼类分类研究概况
据文献报道,我国已命名的鱚科鱼类有14 种,除砂鱚是否为大指鱚属鱼种尚存在争议外,其它皆为鱚属鱼种(表1)。SHAO Kwang-tsao,et al[15]对台湾地区鱚科鱼类进行了重新整理,确定台湾地区1 属8 种,其中小眼鱚为台湾特有种类。国内主要文献记载大陆沿岸广泛分布有斑鱚、多鳞鱚、少鳞鱚、湾鱚[7,37-39],此外还有亚洲鱚、大头鱚、北部湾鱚等及新发现的中国鱚、邵氏鱚、黑带鱚[1,17,20-21,40,41]。国内鱚科鱼类现有研究资料较为陈旧,相关文献中信息不全甚至有误。台湾鱚鱼研究文献自邵广昭后,至今鲜有相关报道。大陆资料匮乏且存有诸多错误,例如成庆泰等主编的《中国鱼类系统检索》[7],中国科学院动物研究所等主编的《南海鱼类志》[42],朱元鼎等主编的《东海鱼类志》[38]238-240,《福建鱼类志》编写组的《福建鱼类志》[43]都只记载了斑鱚、多鳞鱚、少鳞鱚,其中有些描述尚存在争议,如“多鳞鱚在我国沿海皆有分布”;一些新近出版的鱼类志(鱼类图鉴)等的相关资料依然遵循以往出版的鱼类志,未作出相应更新及错误订正[1,4,15,17,20-21,24,39,42-47],见图2。

图2 我国鱚科鱼类研究主要历程Fig.2 Development history of Sillaginidae research in China
GenBank(https://www.ncbi.nlm.nih.gov/)数据库中鱚科鱼类序列也存有较多问题,主要分为2 大类,一个是学名的误用,国内有研究Du Jianguo,et al 将斑鱚Sillago aeolus 误用为Sillago maculata,而后者只在澳大利亚东海岸近海有分布;另外就是鱼种鉴定错误,王晶等将少鳞鱚Sillago japonica 与多鳞鱚Sillago sihama,朱元鼎等许多学者多年来也将亚洲鱚Sillago asiatica、中国鱚Sillago sinica 和多鳞鱚相混淆[6,36,38,48-50]。
由于鱚科鱼类形态和体色上的相近,有些鱼种之间差异体现在鱼鳔等内部解剖,使得该科鱼类的分类一直存在诸多问题。MCKAY[1]最初依据外部性状指标和鱼鳔特征将鱚科鱼类各鱼种分类,但近年来随着分子方法的成熟,越来越多的新种及隐存种被发掘。它们大都与多鳞鱚外形甚至鱼鳔特征都极相似,不借助分子手段仅利用传统形态方法很难将之准确区分开来[6,36]。近几年新发现的Sillago caudicula,Sillago suezensis,Sillago sinica,Sillago shaoi 在之前的研究中都一直被误认为是多鳞鱚[11,13,17,21]。由此可见,分布范围最为广泛的多鳞鱚,极有可能是由诸多外形上极相似的不同种组成的复合种群。
截至目前,我国明确记录的鱚科鱼类有9 种(图版)[6,17,20-21]。样品采集地见表2。

表2 中国鱚科鱼类样品地理分布Tab.2 Geographical distribution of sampling sites of Sillaginidae in China
图版Ⅰ斑鱚Sillago aeolus 背鳍X~XI,I,18~20;臀鳍II,17~19;侧线鳞65~74,侧线上鳞5~7;第一鳃弓鳃耙数2~3+4~8;脊椎骨:腹椎13~14,血道弓门4~7,尾椎13~16,共33~35。体侧侧线附近及背方具两行黄褐色斑块,自头后至尾柄。鱼鳔具一个前部延伸和一个后部延伸;两侧各具3~4 侧部延伸,结构渐变简单。
图版Ⅱ亚洲鱚Sillago asiatica 背鳍XI~XII,I,20~22;臀鳍II,20~23;侧线鳞64~74,侧线上鳞4~5;鳃耙数2~5+6~10;脊椎骨:腹椎13~15,血道弓门4~8,尾椎12~17,共33~35。新鲜状态下体侧中央有一条窄的银白色条带。鱼鳔具一个前部延伸;左右各具一前外侧延伸,皆沿鳔体伸向后部,一般约达到鳔体的1/4~1/5 处;具一个后部延伸。
图版Ⅲ湾鱚Sillago ingenuua 背鳍XI~XII,I,20~22;臀鳍II,20~23;侧线鳞64~74,侧线上鳞4~5;鳃耙数2~5+6~10;脊椎骨:腹椎13~15,血道弓门4~8,尾椎12~17,共33~35。新鲜状态下体侧中央有一条窄的银白色条带。鱼鳔具一个前部延伸;左右各具一前外侧延伸,皆沿鳔体伸向后部,一般约达到鳔体的1/4~1/5 处;具一个后部延伸。
图版Ⅳ少鳞鱚Sillago japonica 背鳍X~XII,I,20~23;臀鳍II,22~24;侧线鳞65~73,侧线上鳞3~4;第一鳃弓鳃耙数3~5+7~10;脊椎骨:腹椎13~14,血道弓门8~11,尾椎11~13,共34~36。鳞片易脱落,样品新鲜时体侧中部具一模糊锯齿状银色条带。鱼鳔具一个前部延伸;左右各具一前外侧延伸,皆伸向前部且一般比前部延伸略短;具一个后部延伸。
图版Ⅴ多鳞鱚Sillago sihama 背鳍X~XI,I,20~22;臀鳍II,20~24;侧线鳞65~77,侧线上鳞4~6;鳃耙数2~4+5~8;脊椎骨:腹椎14~15,血道弓门2~7,尾椎13~16,共34~35。新鲜时臀鳍近透明,无色素黑点分布。鱼鳔具两个前部延伸;两个后部延伸,两后部延伸基部连接紧密;鳔体两侧各具一前外侧延伸;具一小管,小管基部远离两后部延伸基部。
图版Ⅵ邵氏鱚Sillago shaoi 背鳍XI,I,20~22;臀鳍II,21~22;侧线鳞70~73,侧线上鳞5~6;鳃耙数3~4+5~6;脊椎骨:腹椎13~14,血道弓门4~6,尾椎14~17,共34~35。鱼鳔具两个前部延伸,两个后部延伸间距较大且两后部延伸起始端分离;具一小管与肛门相连,小管基部位于两个后部延伸起始端之间。
图版Ⅶ中国鱚Sillago sinica 背鳍X~XII,I,20~22;臀鳍II,20~23;侧线鳞72~80,侧线上鳞5~6;鳃耙数2~3+5~7;脊椎骨:腹椎12~14,血道弓门2~6,尾椎18~22,共37~39。第二背鳍沿鳍条有3~4 行规则排列的黑色小斑点。鱼鳔具前外侧延伸后亚延伸沿鳔体向后延伸,达鳔体长的1/3~2/3,且延伸上具向外侧的小刺状结构,数目不定。
图版Ⅷ黑带鱚Sillago nigrofasciata 背鳍X~XII,I,20~22;臀鳍II,20~22;侧线鳞67~75,侧线上鳞4~6;鳃耙数2~4+5~8;脊椎骨:腹椎14~15,血道弓门3~7,尾椎13~18,共34~35。新鲜状态下,体侧中下部经常会具一黑色条带;鱼鳔两后部延伸间缝隙较大,且两后部延伸基部连接紧密;具一小管与肛门相连,小管基部接近两后部延伸基部。
图版Ⅸ拟多鳞鱚Sillago parasihama 背鳍XI~XII,I,19~21;臀鳍II,19~21;侧线鳞65~70,侧线上鳞4~5;鳃耙数2~3+5~7;脊椎骨:腹椎14,血道弓门4~7,尾椎13~16,共34。鱼鳔前外侧延伸后亚延伸沿鳔体向后延伸,长度因个体而异,达鳔体长的1/5~1/2,且延伸上具向外侧的小刺状结构,数目不定[6,17,20-21]。
4 展望
由于缺乏系统的分类学研究,鱚科鱼类的生物地理一直未能得到有效的认识。MCKAY 对31 种鱚科鱼类的地理分布进行过详细记述,但未提及各物种成因、进化路径及分化时间,仅预测鱚科可能起源于始新世的古特提斯海道(tethys seaway),分布于印度—西太平洋沿岸的各个种类在第三纪中新世逐渐分化[1,51]。虽然MCKAY、KAGA 等学者先后报道鱚科鱼类新种、新属,近年也有多个新种发现,但也只停留于分布与物种描述层面[11,14,17,20-21,52],由于鱚科鱼类分布广泛,其科内物种厘定及系统发育关系尚存在众多争议,中国也存在鱚属未明种类及许多同物异名和学名误用问题[6]。
传统的鱚科鱼类分类学研究仍受到诸多限制。近年来迅速发展的分子系统学研究为鱚科鱼类的分类学研究提供了一条客观有效的途径,尤其是DNA 条形码的应用为其新种及隐存种的发掘提供了充实的证据[20-21,36]。由系统发育学和基因组学交叉形成的系统发育基因组学(Phylogenomics)具有信息量大、结果可靠的优点,能有效消除横向基因转移和类群间基因进化速率差异等因素对系统发育树的影响[53],被逐渐用于动物系统发育与进化研究,这门崭新交叉学科的出现也成了重建生命之树的一条迷人途径[54-55]。
随着测序技术日渐成熟,全基因组测序越来越普遍,也使得生物学在分子层面的研究更加深入,为探讨物种间和种内群体间的遗传多样性、自然选择及物种形成等方面都带来了便捷[56]。自2001 年首先对斑马鱼的基因组测序起,到目前为止已有上百个鱼种的全基因组测序完成。国内开展鱼类全基因组学研究虽然起步较晚,但呈现快速增长的趋势[57]。2014 年,CHEN Songlin,et al[58]发表了我国第一种鱼类的基因组测序和全基因组序列图谱,使半滑舌鳎成为世界上第一个测定了全基因组序列的鲽形目鱼类;同年,我国学者相继发表了鲤、弹涂鱼、大黄鱼全基因组结果[59-61]。2018 年,第1 个鱚科鱼类—中国鱚全基因组序列被破译,相关成果发表于GigaScience[62];广东海洋大学于2020 年报道了养殖多鳞鱚的全基因组序列[63]。高质量基因组的发表极大地推进了鱼类系统发育基因组学研究的进程[64],近年发表于国际著名学术刊物的多篇文章[65-68],也为研究全球鱼类生物地理格局特征及其多样性形成机制、建立可靠的鱚科系统发育关系奠定了良好基础。
致谢:本文是15 a 来薛泰强等多名博士、硕士研究生关于鱚科鱼类分类研究成果的总结,研究也得到了澳大利亚昆士兰博物馆McKay 先生、台湾大学沈世杰先生、台湾中央研究院生物多样性研究中心邵广昭教授等诸多专家的指导和帮助。
