基于Ecopath 模型的中街山列岛海域曼氏无针乌贼生态容量的评估
2024-05-13石雯静张漫瑶
石雯静,张漫瑶,赵 晟
(浙江海洋大学海洋科学与技术学院,浙江舟山 316022)
中街山列岛海域位于浙江省舟山市东北部。该海域受海洋洋流交汇影响以及长江口等河流汇入,使该海域饵料十分丰富。天然优越的地理位置与自然环境条件为许多海洋资源提供了非常优越的产卵场地、育幼场地以及索饵场地。在该海域生存繁殖的海洋生物种类丰富,是我国重要的经济鱼类资源产地,包括大黄鱼Larimichthys crocea、小黄鱼Larimichthys polyactis、带鱼Trichiurusl epturus、曼氏无针乌贼Sepiella maindroni、鲳鱼、鳗鱼等。同时也是一些生物种类的繁殖场所。东极区是曼氏无针乌贼在中街山列岛海域产卵的中心地带,也是厚壳贻贝Mytilus coruscus 和荔枝螺Thais 的主要产地。这一地区具有良好的生态环境和丰富的营养物质,为底栖海藻的生长和繁殖提供了良好的环境,蕴藏着丰富的底栖海藻资源。
由于近年来中街山列岛海域渔业资源的过度捕捞、管理的缺陷以及海域生态环境的恶化,导致该海域渔业资源面临衰竭。曼氏无针乌贼作为该海域面临衰竭的渔业资源之一,亟需人们采取科学合理的渔业资源恢复措施,保护和修复生态环境。海洋渔业资源增殖放流的优势是人为可以控制其增殖放流量,从传统单一的只注重渔业经济增长的方式转向为更加注重增殖放流质量、效益以及多样化的提高,更加注重该海域生态环境质量的提高。增殖放流目前是我国恢复海洋环境中受损生物资源的主要措施。增殖放流活动是比较系统的一项工程,旨在恢复生物资源的数量,确保放流水域的生态系统环境不受破坏,同时也不影响生物物种遗传特征等生物特征。
目前,在国际上已开展了对生物资源的增殖放流的研究。在国外,生物资源增殖放流活动的主要目标是补充资源量、修复生态以及改善生态结构[1]。而在我国,则以提高资源储量为主。渔业资源增殖放流是一种可以使渔业资源得到恢复的有效手段,它在渔业资源的数量、保护和修复水域生态环境、优化渔业资源群落结构、推动渔业资源可持续发展等方面发挥了巨大的作用。有研究表明,生物资源增殖放流会导致野生种群面临生态系统失衡和遗传多样性下降的风险,但这是对资源出现衰退的物种补充资源量最快速、最直接和最有效的方式[2]。
科学合理的渔业增殖放流量对于渔业资源恢复起到非常重要的作用。渔业资源增殖放流量太多或者太少都不能达到渔业资源恢复的目的,因此需要制定科学合理的渔业资源增殖放流策略,科学合理地确定水域增殖放流对象的生态容量,以更好地恢复生物资源量,修复生态环境,改善生态结构,达到良好的增殖放流效果。生态容量即生态承载力。对生态容量进行评估的研究方法有很多种,有些是根据环境因素,或者是食物网中的其中一部分,比如浮游植物、浮游动物,来计算得到的。然而,这种方法并没有将整个生态系统的情况考虑进去。生态系统中不仅仅需要考虑存在捕食与竞争关系,环境因子的影响也不容忽视。考虑到这些情况,目前许多专家与学者使用Ecopath 建模来科学合理的评估生态容量。Ecopath 模型是根据渔业生态环境状况来制定的一种渔业管理模式,它为渔业决策提供了一种有效的方法。Ecopath 模型可以在一定时间尺度上,快速、精确地模拟出一个特殊的生态系统,如生物群落的组成等,将成为新一代水生生物群落研究的关键核心工具。以Ecopath 模型为基础,将水体中的生物体分为多个功能群,并在此基础上,根据各个功能群的生物学特征、食性关系等,构建Ecopath 模型,并对其进行分析,进而对水体中的生态环境进行评估。
1 材料与方法
1.1 研究方法
1.1.1 Ecopath 模型概述
EwE(Ecopath with Ecosim)是美国夏威夷海洋学研究所的Polovina 教授于1983 年首次使用的一种基于线性方程的生物能量流动模型,它能够准确地描述生物体在不同营养级之间的物质交换和分配,研究能量在各个营养层次的流动情况。并能够准确地反映生物体内的物质循环。Ecopath 模型属于静态模型,也被称为生态系统稳定营养模型[3]。Ecopath 软件是当前生态环境效果评估的一个主要工具。目前,Ecopath 模型已被越来越多的专家和学者应用于海洋、湖泊、水库、河口、人工鱼礁区域等水体生态系统的评价中。可以用以下公式表示:
式中:Q 代表消耗量;P 代表生产量;R 代表呼吸量;U 代表未消化的食物量。
Ecopath 模型的定义指出,一个生态系统是一个由一系列相关的功能群组成的,并且功能群都包含了整个生态系统的能量流。功能组的数量范围一般在12~50 之间。在这些功能群中,包含了碎屑,浮游生物,以及具有同样生态特征的鱼类种群。Ecopath 模型构建的前提条件是,在规定的时间内(一般是1 a),生态系统处于一种平衡状态。以热力学原理及能量守恒定律为基础,模型对生态系统中的总输入量与输出量是相等的,且每一个生物种群的系统能量输入和系统能量输出保持平衡,也可以表达为生产量-捕食死亡-其他自然死亡-产出量=0。该模型由一套联立线性方程式来界定,每个方程式都是该生态系统的一种功能群。用以下数学公式表示:
式中:Pi代表功能组i 生物的生产量;Yi代表总捕捞量;Bi代表生物量;Ei代表净迁移;BAi代表生物量积累;Ri代表呼吸量;Ui代表未消化的食物量;Qi代表消耗量;MOi代表其他死亡率;M2i代表捕食死亡率;EEi是功能组i 的生态营养效率,代表生产量在系统中利用的比例。在Ecopath 模型中,Bi、(P/ B)i、(P/ B)j和EEi中4 个参数中,模型需要3 个已知的数据,可以由模型计算得出一个未知的参数。
1.1.2 功能组划分
Ecopath 模型主要原理是把生态系统界定成由一系列相关的具有生态系统关联的生态系统功能群,这些生态系统功能群简称为功能组。本研究拟在已有的中街山列岛海洋生态系统的基础上,结合模型操作规则,选择食物组成、年龄组成、取食方式、个体大小、捕获物类型等基本相同的物种,对其进行归并,从而实现对食物网络的简化。将具有相似功能地位的生物划分为一个功能组。根据已有调查数据,笔者以中街山列岛海域的生物物种、生物学特性、生境特征及食物来源等为基础,将中街山列岛海域生态系统分为17 个功能组,这17 个功能组基本覆盖中街山列岛海域的整个能流过程。功能组包括浮游植物、碎屑、浮游动物、底栖甲壳类、虾类、小型底栖动物类、蟹类、头足类、曼氏无针乌贼、小黄鱼、鳀科类、小型底层鱼类、其他中上层鱼类、大黄鱼、大型底层鱼类、带鱼、其他食鱼性鱼类功能组。由于该文主要针对曼氏无针乌贼的增殖放流情况进行研究,所以将曼氏无针乌贼单独设为一个功能组。大黄鱼、小黄鱼、带鱼等在该海域鱼类也具有代表性,因此也单独设为一个功能组。具体生态系统模型功能组划分见表1。

表1 中街山列岛海域Ecopath 模型各功能组及主要生物种类Tab.1 The functional groups and main biological species of the Ecopath model in the adjacent waters of the Zhongjieshan Islands
1.2 数据来源及参数指标
根据东海区营养动力学有关的研究划分该海域生态系统功能组。其中消耗量与生物量的比值(Q/B)和生产量与生物量的比值(P/B)主要来源于国际水生资源管理中心网站和历史资料数据[4-7]。由于Q/B、P/B 部分数据难以获得,本模型参考与中街山列岛海域具有相似生态系统特征的模型数据,如东海海域Ecopath模型相关系统参数,东海海域关于生态营养动力学有关研究文献[6],其中碎屑的生物量数据参照李睿等[8]、生态系统功能组生物P/B 值主要以林群等[9]为参考计算、Q/B 值主要参照欧阳力剑等[10]计算方式得出。其中生态系统渔业资源数据主要来源于环境现场数据与历史数据的收集与分析以及已发表的文献的数据。通过对渔业资源和生态环境的历史文献数据分析整理得出。食性分析矩阵的来源主要是对取样的鱼的胃含物和有关的文献进行的分析[11]。生态营养转换效率(EE)一般较难直接测定,一般EE 值由模型自动计算得出。
在中街山列岛海域Ecopath 模型中,经过模型相关参数调试后的输入和输出参数见表2。从表2 可以看出中街山列岛海域生态系统中各功能组的营养级范围在1~4.648 之间。营养级最高的为食鱼性鱼类,为4.648。其次为带鱼及大型底层鱼类,营养级分别为4.407 和4.317。碎屑和浮游植物是营养级最低的,均为1。功能组中浮游动物营养级是2。底栖生物营养级为2.4 左右。主要经济生物资源类的营养级在2.5~4.5之间。中街山列岛海域中生态系统各功能组生态营养效率(EE)值均小于1,在0.013~0.991 之间,生态系统均处于平衡状态。可通过模型进行估算缺失的某些参数,如P/B 参数缺失的功能组。据表可以看出该海域中曼氏无针乌贼的营养级为3.562,生态营养效率(EE)值为0.991,在该生态系统内属于中等水平。

表2 中街山列岛海域Ecopath 模型输入参数及估算参数Tab.2 Basic input parameters and estimated parameters of Ecopath model in the adjacent waters of Zhongjieshan Islands
2 结果与分析
2.1 生态系统总体特征
表3 为中街山列岛海域Ecopath 模型建立后,输出的中街山列岛海域生态系统特征总参数。通过参数特征对中街山列岛海域生态系统的能量流动过程、发展状况和总体特征进行了分析。
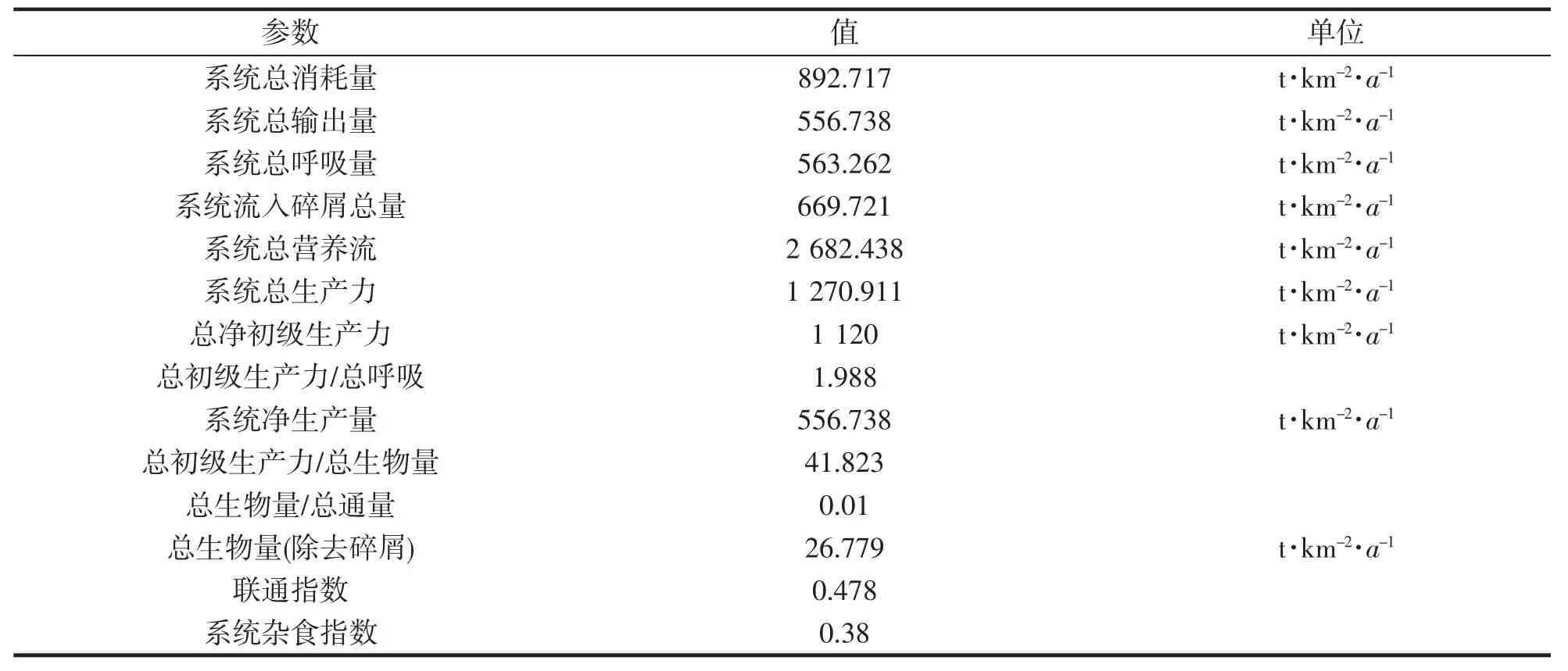
表3 中街山列岛海域生态系统特征参数Tab.3 Ecosystem characteristic parameters in the adjacent waters of Zhongjieshan Islands
当前系统状态下,系统总消耗量为892.717 t·km-2·a-1,总输出量为556.738 t·km-2·a-1,总呼吸量为563.262 t·km-2·a-1,流向碎屑总量为669.721 t·km-2·a-1,系统总营养流2 682.438 t·km-2·a-1,系统总生产力1 270.911 t·km-2·a-1,总净初级生产力1 120 t·km-2·a-1,总初级生产力/总呼吸为1.988,净生产量556.738 t·km-2·a-1,总初级生产力/总生物量为41.823,总生物量/总通量为0.01,总生物量(不计碎屑量)为26.779 t·km-2·a-1。
总初级生产力/呼吸总量之比(TPP/TR)是衡量一个地区生态系统成熟性的一个关键指标,其数值逼近1 表示该地区生态系统的成熟性越高。当其值超过1 时,表明该生态系统处于未成熟状态。中街山列岛海域TPP/TR 指数为1.988。表示中街山列岛海域生态系统成熟度低。总生物量/总通量(TB/TT)也是衡量一个区域生态系统的成熟度的重要指标,其数值较大,说明该区域的物种多样性较高。结果表明,中街山列岛海域的TB/TT 很小,仅为0.01,说明中街山列岛海域的生态系统物种多样性相对于其他海区来说很小,且不够稳定。
系统连通性(CI)和系统杂食性(SOI)是反映生态系统中食物网络关系的2 个重要指标,随着生态系统向成熟化方向发展,其功能群间的关系也会变得更加复杂。系统联结指数(CI)是指一个食物网络的真实联结数量与其潜在联结数量的比值。系统杂食性指标(SOI)反映了生态系统中各功能群对食物的分配状况。系统杂食性指标(SOI)在0~1 范围内,0 表明只对某个营养级别的食物进行摄取,并具有很强的针对性,1的数值代表了不同的营养水平。中街山列岛海域CI 和SOI 分别为0.478 和0.38,这2 个指标都远低于1,说明明中街山列岛海域各功能群落间的掠食性关系比较简单。各功能群落间的关联性不强,其食物链的组成比较单一。
2.2 生态功能组营养级与食物网结构分析
图1 是通过本研究调查得出的中街山列岛海域生态系统食物网结构简图。通过中街山列岛海域Ecopath 模型把该海域复杂交织的生态系统进行简化梳理,用圆圈以及线条的方式构成一个相对简单的食物网络体系图。从图1 可以清楚地了解到,在这一海域的生态系统中,各生态系统的营养流的相互关系,以及各生态系统间的食物竞争与捕食效应。在图中,圆圈及大小代表着系统中的各个功能组及生物量的大小,线条表示的是各个功能组之间系统能量的传递方式,线条的粗细表示的是能量流动的大小。

图1 中街山列岛海域生态系统能量通道示意图Fig.1 Schematic diagram of the energy channel of the ecosystem in the adjacent waters of the Zhongjieshan Islands
由图可知,中街山列岛海域主要集中在1~5 营养级之间进行能量流动,它们主要沿着2 个方向,即从碎屑开始的碎屑食物链和从浮游植物开始的牧食食物链。整个生态系统的能量来源主要为碎屑类和藻类,且从低级到高级的顺序是逐渐减少的。由图可以看出营养级最低的为碎屑和浮游植物,营养级最高为食鱼性鱼类,由之前的研究结论可知,曼氏无针乌贼的营养级为3.562,在调查海域中属于中等营养级。中街山列岛海域生态系统存在5 个营养级,第一营养级是碎屑和浮游植物;第二营养级包括浮游动物、小型底栖动物类等;第三营养级包括小黄鱼、小型底层鱼类等;第四营养级大多为杂食性鱼类;第五营养级大部分为一些食鱼性鱼类。
2.3 生态位重叠分析
功能群的生态位重合性分析是指在研究对象所涉及的生态系统中,一些生物在其所在的生态系统中发挥着类似的作用,而且它们的生态位相对相近,因此它们之间的竞争也就更加显著。比如,它们需要的食物种类,它们的生存环境,它们的捕食习性等。在所研究的生态系统中,当2 种功能组生态位相同或相似的条件越多,其生态位重叠现象就会越显著,其生态重叠度数值也会越高。在自然环境中,环境资源是有限的,生物不可能在环境中单纯地获取自己所需的食物与能量。所以,在生态系统中,生物之间必然会存在竞争关系,通过种内、种间竞争的方式,来获得自己所需的食物和栖息环境,进而进行生长发育并繁殖后代。在经过了竞争之后,一定会有一方被淘汰,从而做出了让步,比如迁移到其他环境,选择更换其他生物为食物来源,或是选择其他时候来进行捕食活动等,从而与竞争对手在生态位上进行了交叉交替,从而停止了竞争[12-13]。
通过中街山列岛海域Ecopath 模型对生态系统食物网结构进行分析,进而分析生态系统功能组之间的饵料重叠指标以及捕食者重叠指标两个指数,以此反映生态系统功能组之间的生态位重叠的状况。其中,饵料重叠指数代表的是2 个功能组之间是否存在着类似的食物来源,当饵料重叠指数取值为0 时,表明2 个功能组之间没有相同的摄食情况,当饵料重叠指数的取值为1 时,表示2 个生态系统功能组对同一摄食对象竞争巨大。捕食者重叠指数反映了2 个功能性群体中捕食者和被捕食压力的相似程度,其数值为0 表明没有天敌,数值为1 表明两个功能性群体中存在相同天敌。分析结果见图2,生态位重叠度指数最高的是头足类与蟹类,重叠度在0.8 以上。食鱼性鱼类与鳀科类重叠指数最低,在0.1 左右。
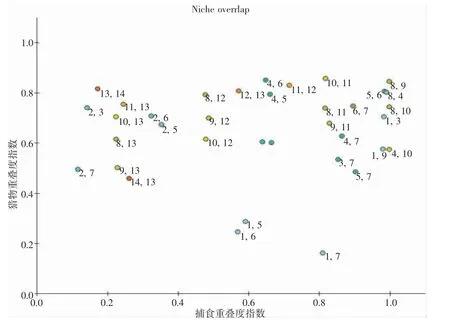
图2 中街山列岛海域生态系统的生态位重叠分析Fig.2 Analysis of niche overlapping of ecosystems in the adjacent waters of Zhongjieshan Islands
2.4 营养结构和能量流动
为使食物网关系简单,更便于对各营养级的能量流动及分配进行分析,将整合营养级这一概念引入Ecopath 模型中。通过营养级聚合原理,主要是将不同功能组的营养流合并为几个以整数表示的营养级[14]。Ecopath 模型是将各个功能组根据它们在食物源中所占有的营养水平的比例,划分到不同的整合营养级中。不同功能组的能流在生态系统中的分配可分为相对流量和绝对流量表示。
通过模拟计算,得出中街山列岛海洋生态系统的综合营养等级为I-XII。但是由于营养级VII-XII 的营养流动相对较小,在本研究中可以忽略不计,因此可以将中街山列岛海域设为VI 个营养级组成的生态系统。生物量和生态系统总流量在第I 个营养级是最高的,且随生态系统营养级的升高而减小。
表4 为中街山列岛海域生态系统各整合营养级的能量流动,通过营养级整合,将中街山列岛海域生态系统整合为VI 个营养级层次,生态系统总流量随着生态系统营养级的升高而逐渐降低。生态系统营养级I 以及II 的系统能量流动较高,但生态系统营养级VI 的系统能量流动较低,能量流动符合能量金字塔规律。能量流动主要分布集中在前IV 个营养级,中街山列岛海域生态系统总能流为2 682 t·km-2·a-1。第I 营养级的生态系统总流量为1 790 t·km-2·a-1,占比生态系统总能流的66.7%;其中被捕食量为803.1 t·km-2·a-1,占总被摄食量的89.9%。第I 营养级是系统能量的主要来源,主要由浮游植物和有机碎屑组成。第II 营养级总能流为803.1 t·km-2·a-1,占系统总能流的29.9%,其中被捕食、流入碎屑和用于呼吸的能流量分别为76.82 t·km-2·a-1、206.7 t·km-2·a-1、519.6 t·km-2·a-1,分别占总被捕食,总流入碎屑,总呼吸量的8.6%,30.8%,92.24%。第Ⅲ营养级总能流为76.82 t·km-2·a-1,占系统总能流的2.8%。第IV 到第VI 营养级的能流较小,分别占系统总能流的0.38%,0.07%,0.01%。说明能量流动大部分还是在较低的营养级,中街山列岛海域第I 营养级的初级生产者和碎屑功能组是生态系统能量的主要供给来源。
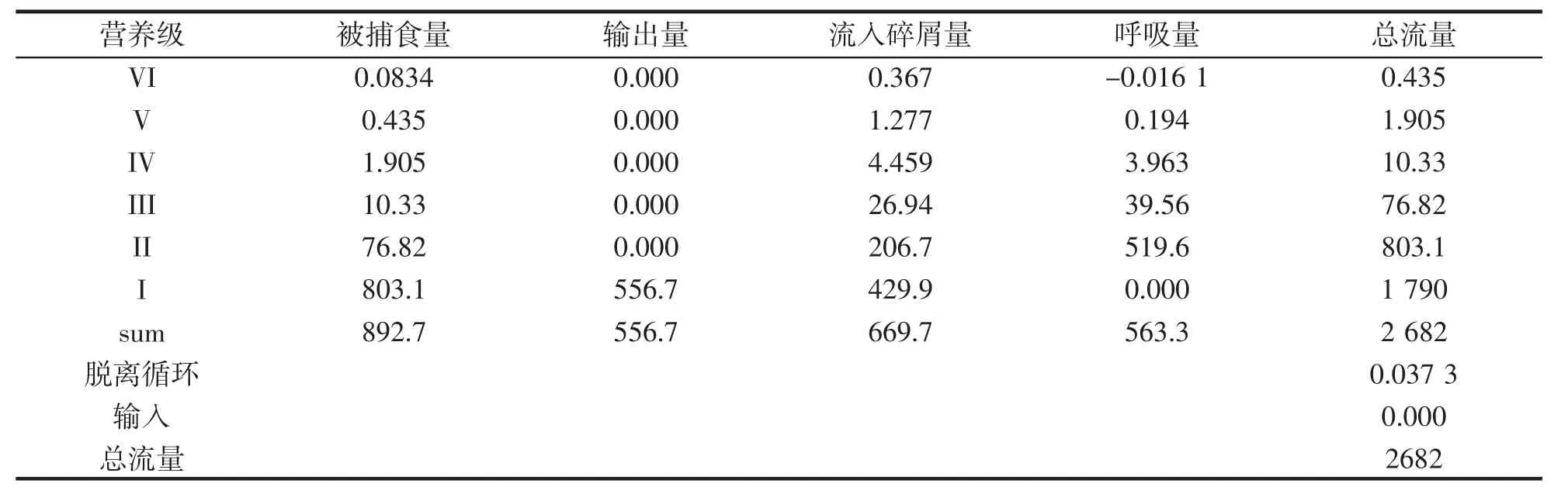
表4 中街山列岛海域生态系统各整合营养级的总能流Tab.4 The total energy flow of the integrated trophic levels of the ecosystems in the adjacent seas of the Zhongjieshan Islands t·km-·2a-1
由表5、6 可以看出,在生态系统总能流中,来自初级生产者流量为1 885 t·km-2·a-1,来自碎屑流量为797.0 t·km-2·a-1,各占70.28%和29.72%,这表明,营养级I 的大部分能量都是以碎屑的形式沉淀下来,而没有被利用,因此,它的能量利用的不充分。能量还有剩余。而且,牧食食物链的能量流比碎屑食物链稍高一些。各功能组生物量在高营养级生物中所占比例很小,主要是因为高营养级生物要以低营养级生物为主要食物来源,故所占比例不高。
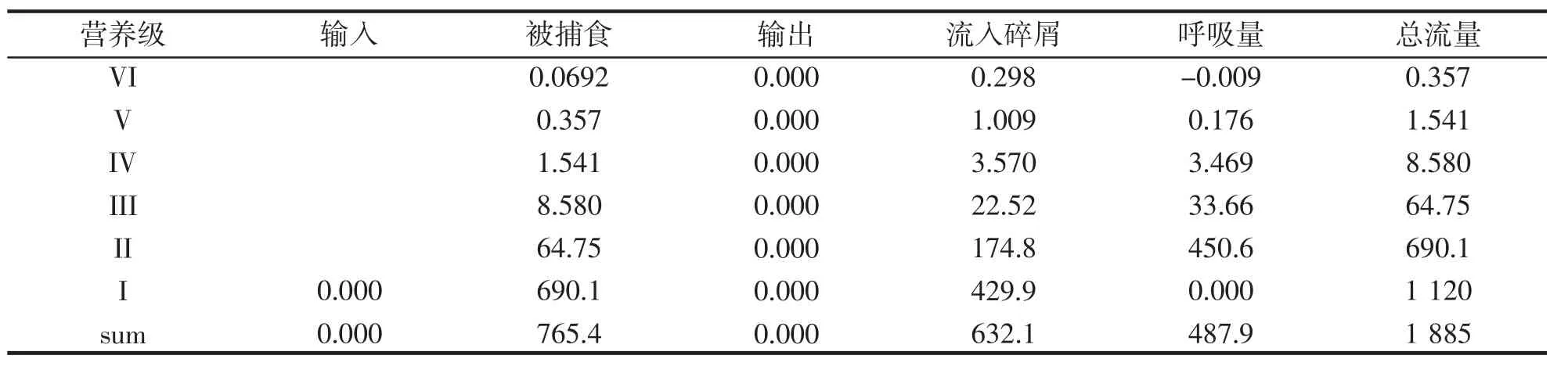
表5 从初级生产者流入不同营养级的能流Tab.5 Energy flow from primary producers to different trophic levels t·km-·2a-1

表6 从碎屑流入不同营养级的能流Tab.6 Energy flow from debris into different trophic levels t·km-·2a-1
2.5 增殖容量估算
经模型数据运算后得出,曼氏无针乌贼的营养级为3.562,在营养级中处于中上水平,是浮游生物及底栖生物的捕食者。随着曼氏无针乌贼生物量的提高,对其它食物类如底栖动物的取食压力也将随之增加。从模型模拟和中街山列岛海域的食物链结构来看,这一区域的食物链发育程度不高,很少有高营养等级的生物体。由于底拖网、桁杆拖网等捕鱼工具的应用,对底层鱼类或底栖生物等水生生物进行了过量的捕捞,从而对海底环境和动物的栖息地造成了某种程度的破坏,一些破坏甚至是不可逆的,这就造成了高营养级生物向低营养级生物的转化,寿命长的生物向寿命短的生物转化[15]。为此,应进行适宜的增殖放流,以确保生物量的稳定。多数研究表明,曼氏无针乌贼在全年的增殖放流中都表现出波动性,这种波动性可能是由于其增殖放流策略和捕捞策略所致。因此,有必要对曼氏无针乌贼进行有效的增殖与放流,以确保其生物资源的可持续发展。在利用Ecopath 模型来估计算中街山列岛海域曼氏无针乌贼的增殖能力时,当输入所需要的原始资料时,其输出的EE 并不都是1,这说明该模式无法达到完美均衡,所以需要对模型的有关数据进行调整,本研究中主要调试了食物矩阵中变量较小的资源数据,例如底栖生物类资源数据。
在对中街山列岛海域曼氏无针乌贼的生态承载能力进行评价时,需要将Ecopath 模型下的曼氏无针乌贼的种群数量最大化,并将模型调节到各种群的生态效率(EE)大于1,使曼氏无针乌贼种群的种群数量达到最大,从而保持生态系统的稳定。最大增殖容量是在已有的生物量基础上,减去原有的生物量得到的生物量值。曼氏无针乌贼历史最高年产量浙江省曾达到6×104t。利用海域面积进行估算后,考虑到中街山列岛海域是曼氏无针乌贼的适宜生境地。通过调查研究,将该模型中中街山列岛海域曼氏无针乌贼原始生物量设为0.01 t·km-2。模型中,曼氏无针乌贼生物量从原始数据基础上以0.03 t·km-2为补偿,当不断增加曼氏无针乌贼的生物量至0.16 t·km-2时,模型中各生物群组EE 值小于1,此时生态系统处于平衡状态。继续增大至0.17 t·km-2时,模型中小型底层鱼类的EE 值为1.001,其他功能组的EE 值均小于1,此时生态系统处于不平衡状态。因此由模型可以得出此时曼氏无针乌贼的生态容量约为0.16 t·km-2。具体调试过程见表7。

表7 曼氏无针乌贼增殖容量在模型中的变动情况Tab.7 The variation of the releasing capacity of S.maindroni in the model
中街山列岛海洋特别保护区面积为202.9 km2,除去岛陆面积后大约是193 km2。经计算,中街山列岛海洋特别保护区的曼氏无针乌贼生态容量为30.8 t。在之前的研究中,表明曼氏无针乌贼幼体的存活率在受饵料的影响下的养殖存活率在33.3%~67.5%之间[16],在本研究中假定其存活率同样如此。除此之外,考虑到中街山列岛海域作为曼氏无针乌贼的适宜生境地,结合曼氏无针乌贼的生活习性、洄游、产卵特性、近海生长特性等渔业生物学特征以及浙江省曼氏无针乌贼资源演变过程和演变原因等因素后。据研究,以现在的捕捞状态计算,每放流10 000 粒受精卵可形成捕捞产量137.3 kg[17]。按这个标准计算,要形成30.8 t的产量,可放流2 240 000 粒受精卵。根据曼氏无针乌贼受精卵约10 000 粒·kg-1可推算,可放流曼氏无针乌贼受精卵约224 kg。从增殖放流的观点出发,该模型得出的增殖容量只是一个理论放流量,最终实际放流情况则需要根据其渔业状况和生物学特性,并结合当地实地情况,例如经济、社会效益等。
3 讨论
3.1 曼氏无针乌贼增殖放流前后海域生态系统特征对比
为了更加科学合理地增殖放流曼氏无针乌贼,需要对比曼氏无针乌贼增殖放流前后生态系统特征,通过生态系统特征参数是否发生明显变化来判断增殖放流是否科学合理。该模型以系统的成熟度、稳定性、尺度等生物学参数为指标,对系统的整体特性进行了描述。表8 显示了中街山列岛海域生态系统目前的整体特性和曼氏无针乌贼生物量达到其生态承载能力后的整体特性。

表8 中街山列岛海域生态系统整体特征参数变化情况对比表Tab.8 Changes in the overall characteristic parameters of the ecosystems in the adjacent seas of the Zhongjieshan Islands
对比当前生态系统特征状态(V1)与曼氏无针乌贼达到增殖生态容量(V2)时的生态系统总体参数,对比可以看出大部分参数并未发生太大的改变。总消耗量、系统总营养流、系统总生产力略有升高。总消耗量由902.717 t·km-2·a-1增加到893.917 t·km-2·a-1,总呼吸量由563.262 t·km-2·a-1变化到555.822 t·km-2·a-1,系统总营养流由2 682.438 t·km-2·a-1增加到2 691.171 t·km-2·a-1。系统总生产力由1 270.911 t·km-2·a-1增加到1 279.311 t·km-2·a-1。
系统连通度(CI)与杂食性(SOI)可高效地展示生态系统的复杂性特征。总初级生产量/总呼吸量(TPP/TR)是表征生态系统稳定性与成熟度的指标参数,当曼氏无针乌贼达到生态容量时的三者参数变化较小,表明当曼氏无针乌贼达到生态容量时,中街山列岛海域生态系统的稳定性受到的影响较小。
3.2 研究总结
主要以中街山列岛海域为研究区域,利用EwE6.5 版模型软件,通过搜集和整理有关资料,建立中街山列岛海域的Ecopath 模型。在此基础上,探讨了对中街山列岛海域生态系统结构与功能的影响,并对其能流进行了估算。对中街山列岛海域能量流过程、发展状态及系统整体特性进行了研究。该研究可为中街山列岛海域曼氏无针乌贼的增殖能力评估奠定理论基础。研究的主要结果如下:
(1)在查阅国内外有关资料和海洋资源调研的基础上,将中街山列岛海域的食物链进行了简化,建立了17 个生态学功能组,包括碎屑、初级生产者、浮游动物和高营养性鱼类在内的生态学群落,并对该生态学群落的能流特征进行了分析。系统中总流量和总生物量主要集中在前VI 个营养层,各个营养级的生物量、总流量呈金字塔型。中街山列岛海域生态系统各功能组有效营养级范围处于1~4.648 之间。食鱼性鱼类、带鱼、大型底层鱼类等功能组的营养级较高。浮游植物、碎屑、浮游动物等功能组的营养级较低。各功能组的生态营养效率在0.013~0.991 之间。虾类、底层及中上层鱼类等功能组的生态营养效率较高,碎屑,浮游植物、浮游动物等生态营养效率较低。其中,以初级生产者和碎屑为主的I 级生物量最高,而以食鱼性鱼类为主要成分的V 级生物量最低。总体来看,大部分高营养级的鱼类,其生物量相对较少,但其生态转换效率相对较高。在这一过程中,一些功能群具有很高的生态营养效率(EE),这说明它们受到了捕食与捕捞的双重胁迫,只有很小一部分被重新利用。生态系统中各功能组之间的混合营养效应显示,浮游植物和碎屑在生态系统中起到了主要的作用,浮游植物和碎屑作为初级生产者在生态系统中对其他生物功能组都产生一定的正效应。由于生物捕食之间的关系,浮游动物对浮游植物是捕食作用,产生了负效应。由于生物竞争关系,生态系统中底栖生物之间产生负效应。食鱼性鱼类作为高营养级生物,对多数鱼类具有捕食关系,因此对多数生态系统功能组都产生负效应。
(2)中街山列岛海域的能流主要是通过两个途径来实现的。一种是牧食食物链,其起始点是初级生产者,而一种是碎屑食物链,其起始点是有机碎屑。这两条食物链都对中街山列岛海域生态系统的物质平衡和能量传递都起着重要的作用。中街山列岛海域生态系统总能流为2 682 t·km-2·a-1。在生态系统的总能流中,初级生产者和有机碎屑的流量分别占到了系统总能流的70.28%和29.72%,这表明了在生态系统中,牧食食物链的能量流要比碎屑食物链的能量流要高。
(3)中街山列岛海域生态系统总初级生产力/总呼吸(TPP/TR)值为1.988,系统杂食指数(SOI)值为0.38,连接指数(CI)值为0.478,说明中街山列岛海域生态系统功能组之间联系不紧密,食物网结构较为简单。总生物量/总通量(TB/TT)值为0.01。在当前系统状态下,总消耗量为892.717 t·km-2·a-1,总输出量为556.738 t·km-2·a-1,总呼吸量为563.262 t·km-2·a-1,流向碎屑总量为669.721 t·km-2·a-1,总生物量(不计碎屑)为t·km-2·a-1。对比曼氏无针乌贼达到增殖生态容量时的生态系统总体参数后,发现大部分参数都未发生明显的变化,生态系统基本稳定,总消耗量略有升高,由892.717 t·km-2·a-1增加到893.917 t·km-2·a-1,系统连接指数基本保持不变,系统杂食性指数有所升高。其他系统参数大都基本不变,变化基本不大,表明曼氏无针乌贼的生物量达到增殖生态容量时,对该海域其他功能组的生物以及系统平衡和稳定性并未产生较大影响。
(4)结合构建的中街山列岛海域Ecopath 模型分析得出曼氏无针乌贼在该海域的营养级为3.562,属于中等营养级。生态营养效率为0.991。利用模型估算得出中街山列岛海域曼氏无针乌贼的生态容量约为0.16 t·km-2。在考虑了生物存活率、生活习性等各种因素后,经估算得出曼氏无针乌贼在中街山列岛海洋特别保护区的生态容量为30.8 t。以现在的捕捞状态计算,每放流10 000 粒受精卵可形成捕捞产量137.3 kg。按这个标准计算,要形成30.8 t 的产量,可放流2 240 000 粒受精卵。根据曼氏无针乌贼受精卵约10 000 粒·kg-1可推算,可在中街山列岛海洋特别保护区放流曼氏无针乌贼受精卵约224 kg。
3.3 展望
(1)Ecopath 作为一种静态的模型,具有比较理想的状态,它只能从物质的能量均衡角度对增殖能力进行估算,无法对生物体的生长进行评估。在接下来的研究中,可以使用Ecosim 对渔业政策和环境变化等参数进行动态模拟,对在一定时间内对生物量以及捕捞量的影响进行评价,并与Ecospace 进行组合,进行空间性的深入分析。
(2)在建立Ecopath 模型时,针对一些难以获取的数据,例如,胃含物分析数据、生物量数据、P/B、Q/B等,在研究中参考邻近相似海域模型的文献进行数据的整合与参考。但由于数据精准性不足等各种因素会导致模型结果的偏差。在今后的研究过程中,还需要对该地区的数据进行进一步的获取和研究,将会对多个作业类型的调查数据进行更为精确的收集,并将气候、环境等多种影响因素进行全面的考虑,从而实现对该地区生态系统能量流动特征的精确分析,从而使Ecopath 模型的精度和可靠性得到进一步的提升。在此基础上,通过对曼氏无针乌贼放流后的死亡率和成活率进行分析,并结合曼氏无针乌贼本身的特点进行生态承载力评估,从而为曼氏无针乌贼的增殖放流提供更多的理论基础和技术支持。
(3)中街山列岛海域的生物类资源正处在衰退之中,因此,要想使中街山列岛海域的生物资源能够得到良好的发展,就必须对该海域的能量流动进行深入的探讨和研究。比如,可以通过对营养级层次间能流的数据进行分析,来分析生态系统的能流状态。在此基础上,深入分析其复合效应矩阵,从而判断各功能组的关联性特征。除此之外,该研究中还可以考虑将曼氏无针乌贼的栖息地环境,食性等因素进行更加科学的分析,结合生物遗传多样性和遗传结构研究。以提高曼氏无针乌贼增殖放流效果评估的准确性。
(4)随着我国沿海渔业资源的不断减少,如何科学合理地进行捕捞和放流是当前亟待解决的问题。但是,在进行增殖放流研究中,除考虑渔业资源外,还应考虑渔民的经济收入、社会经济、宗教信仰等因素。由于国内在该领域的研究起步比较晚,所以在该领域的应用和发展上仍有许多不足之处。在未来的研究中,应加强对衰退渔业资源的精准分析,以更好更加全面地应对渔业资源压力等问题。
