沿浦湾红树林大型底栖动物稀有种与常见种对物种多度分布格局及物种多样性的影响
2024-05-13周泽宇水柏年方学河刘永钿胡成业
周泽宇,王 晶,水柏年,方学河,刘永钿,周 鑫,李 艺,胡成业
(浙江海洋大学水产学院,浙江舟山 316022)
稀有种通常是指在群落中种群规模小、栖息地狭窄或是分布范围受限的物种[1],稀有种作为生态系统中种类占大多数的物种[2],在维持物种多样性及物种多度分布格局中扮演重要角色,同时对维持生态系统功能和稳定性也有非常重要的作用[3-4]。ZHANG Zhengqing,et al[5]发现稀有种的多样性与功能多样性呈正相关;艾得协措[6]研究发现稀有种能够增强群落中的生态位分化,稳定物种共存模式,同时增加群落的物种丰富度。然而,稀有种由于自身种群规模小,分布区狭小,以及对于环境及栖息地变化敏感等特性,使其往往成为生态系统中最易丧失的物种。因此,明确稀有种在维持多样性和生态系统功能中的作用已成为生物多样性研究与保护的重要内容[7-8]。
在传统的物种多样性研究中,对稀有种的处理存在剔除与保留2 种方式[9-10]。部分研究认为稀有种对群落分析意义不大,但会对统计结果产生干扰[11-12],故通常按照丰度顺序或随机地剔除;但也有研究认为进行稀有种剔除会使统计信息失真[13-14]。稀有种的准确识别是稀有种处理以及开展相关群落生态学研究的重要前提。RABINOWITZ[1]提出了结合地理范围、栖息地特异性和种群大小等方面进行稀有种识别的方法。然而这种方法受限于数据获取的难度,在实际应用中往往仅将物种丰度、频率、相对重要性指数(IRI)等指标作为稀有种的判断标准,导致结果出现一定程度的偏差[15]。MACIEL[16]在Rabinowitz 观点基础上开发了“rrindex”工具包用于计算每个物种的稀有度指数并排序,有效提高了稀有种识别精度与速度。
物种多度分布格局可反映群落内不同物种个体数量的空间分布[17],最早应用于湖泊底栖动物研究[18],随后广泛用于陆地植物群落生物多样性研究[19-20]。相关研究表明物种多度分布格局可作为评估生态系统受污染及扰动时的生态指标[21]。例如,HILL,et al[22]使用物种多度模型评估热带雨林的生境扰动。NILS[23]通过对巴拿马的小型哺乳类动物、非洲中部的啮齿类动物及瑞典中部高山鸟类等多个生物群落进行研究时发现,在稳定的系统环境当中物种曲线通常符合对数正态分布。
沿浦湾红树林于2016 年进行大规模人工引种,均以秋茄Kandelia candel 作为引种对象,现已建成红树林超80 hm2。沿浦湾大型底栖动物群落相关研究较为丰富,覃胡林等[24]在沿浦湾红树林种植前对沿浦湾大型底栖动物进行调查,研究发现当地大型底栖动物群落结构受到较严重的干扰。张苗苗等[25]比较了沿浦湾红树林种植前后大型底栖动物群落生态位与功能群变化,发现秋茄林种植对大型底栖动物生态位和功能群总体上具有积极的改善作用。张丽源等[26]对沿浦湾大型底栖动物优势种尖锥拟蟹守螺Cerithidea largillierti 生境适宜性进行研究,发现其生境适宜性存在较明显的时空异质性。
拟使用MACIEL 方法[16]识别沿浦湾红树林大型底栖动物稀有种,同时依次模拟稀有种丧失、常见种丧失和2 种群落均保留3 种情景,探究稀有种在维持群落物种多样性中的作用;同时利用统计模型、生态位模型和中性理论模型等物种多度分布模型,探究稀有种和常见种对群落物种多度分布格局的影响及其潜在的生态过程,以期对红树林生态系统生物多样性保护与修复提供理论依据。
1 材料与方法
1.1 大型底栖动物采集
于2020 年7 月(夏季)、10 月(秋季)、1 月(冬季)和2021 年4 月(春季)在浙江省沿浦湾红树林区(27°13′10′′N,120°27′30′′E)布设15 个采样点(图1),每个采样点随机设置3 个30 cm×30 cm 的样方框,收集样方框内30 cm 深的沉积物并用0.5 mm 孔径的网筛进行淘洗,将淘洗所得大型底栖动物装入样品瓶加入无水乙醇溶液固定,最后将装有大型底栖动物的样品瓶带回实验室进行分析鉴定。物种名录登记均参照《中国海洋生物名录》和WORMS 网站(http://www.marinespecies.org/)进行校对和复核[27]。大型底栖动物的采集处理和保存分析遵循《海洋调查规范》(GB/T 12763.6-2007)[28]执行。
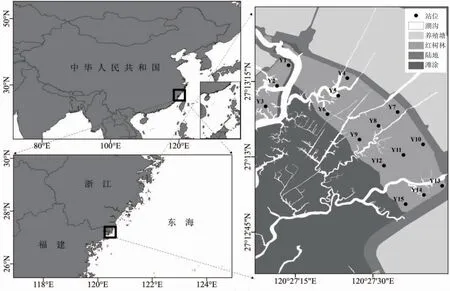
图1 沿浦湾红树林大型底栖动物采样站位分布图Fig.1 Map of macrobenthos sampling sites in the mangrove of Yanpu Bay
1.2 稀有种的识别
基于地理范围、种群规模和栖息地特异性识别红林大型底栖动物稀有种。为避免因季节性变化对大型底栖动物群落稀有种的准确识别造成干扰,以全年大型底栖动物群落为研究对象进行稀有种识别。稀有度指数计算公式分别如下:
式中:rr 为稀有度指数,越接近1 表示物种越稀有,越接近0 表示物种越常见。
将群落中稀有度指数高于平均值的物种定义为稀有种[16]。gri 为地理范围指数,psi 为种群规模指数,hsi 为生境特异性指数。其中,latrange为物种i 纬度分布的范围,而longrange为物种i 经度分布范围。Pmax为物种i 出现的采样点数。hmax为物种i 的最大丰度。
1.3 物种多样性分析
Shannon-Wiener 指数(H′)、Pielou 均匀度指数(J′)、Margalef 丰富度指数(d)和Simpson 指数(E),计算公式分别如下:
式中:S 为总种类数;N 为群落中所有物种的总丰度;Pi为样品第i 种个体丰度与总丰度的比值;Ni为第i个种的丰度。
1.4 物种多度分布模型拟合
综合群落复杂性及生物和环境交互作用的影响,为提高模型的精度与准确度,选择统计模型(对数级数模型、泊松对数正态模型)、生态位模型(断棍模型、几何级数模型)和中性理论模型(复合群落零和多项式模型、Volkov 模型)拟合大型底栖动物群落物种多度分布格局。
1.4.1 统计模型
(1)对数级数模型。对数级数分布是一种离散分布模型,不考虑个体数为0 的物种,计算公式如下:
式中:α 为群落的多样性指数,α>0 且与样本大小无关;当α 较大时,表明该群落中稀有种较常见种多;当α 较小时,表明该群落中常见种较稀有种多。X 为常数0<X≤1,与样本大小有关[29]。
(2)泊松对数正态模型。该模型运用泊松对数正态分布对观察数据进行拟合,则样本中1 个种包含r 个个体的概率为:
式中:r=0,1,2,…,λ>0。此概率取决于2 个参数σ、m,σ 与样本大小无关,m 为中位多度,是样本大小的函数[30]。
1.4.2 生态位模型
(1)断棍模型。假设S 个物种具有相近的分类地位和竞争力,且同一时间内在群落中出现[31]。J 表示群落中物种的个体总数,第i 个物种的多度表示为:
(2)几何级数模型。该模型基于生态位假说,假设总资源量按照优势度依次被各物种所占据,直到剩余资源量无法维持一个物种生存所需为止[32],则第i 个种的理论多度对应的比例Ai为:
式中:Ai为第i 个物种的物种多度值;Q 为资源量总量;p 为优势度最高的物种所占有得资源量的比重。
1.4.3 中性理论模型
(1)复合群落零和多项式模型。假设群落内物种总数固定不变,当一个物种的数量减少,会使其他物种同等数量增加。在群落中,任意一个个体消亡后,替代新物种的出现概率为V,而留存个体的后代被取代的概率为1-V,则该模型中任意取样点内多度为n 的物种数量S 为[33]:
式中:y 为某个物种的多度,θ=2Ju。Γ(n)是Gamma 分布。
(2)Volkov 模型。该模型是复合群落零和多项式模型的进一步拓展,增加物种迁移系数m,并假设其为固定值。因此,模型中多度为n 的物种数S 可表示为:
式中:J 为群落样本大小;Γ(z)为z 的函数;r 为迁移到局域群落的个体数[34]。
1.5 模型拟合效果检验
1.5.1 Kolmogorov-Smirnov 检验
使用Kolmogorov-Smirnov(K-S)检验对模型的拟合效果进行检验。将站位随机划分为2 部分,计算累积分布函数距离(统计量D)并判断是否存在显著差异。为避免出现数据连结的问题,使用基于bootstrap 模拟方法的K-S 检验[35],循环次数为1 000 次,当p<0.05 时表明该模型被拒绝[36]。
1.5.2 AIC 准则
赤池信息准则(Akaike information criterion,AIC)常用于比较模型拟合效果。一般,AIC 值越小,模型拟合效果越好,其公式为:
式中:L 为所使用模型极大似然函数的估计值;k 为所采用的模型中出现的参数数量。ΔAIC>2 时,模型间有显著差异[37]。
2 结果与分析
2.1 稀有种识别
本次调查共采集到大型底栖动物30 种,隶属于6 纲10 目17 科24 属。沿浦湾红树林大型底栖动物的地理范围指数(gri)为0.999 847~1,生境特异性指数(hsi)为0.066 667~1,种群规模指数(psi)为0.007 353~1,稀有度指数(rr)范围在0.357 955~1。其中,rr 平均值为0.65,有16 种物种的rr 值超过平均值,被定义为稀有种,占总物种数的53.33%,剩余物种为常见种(表1)。
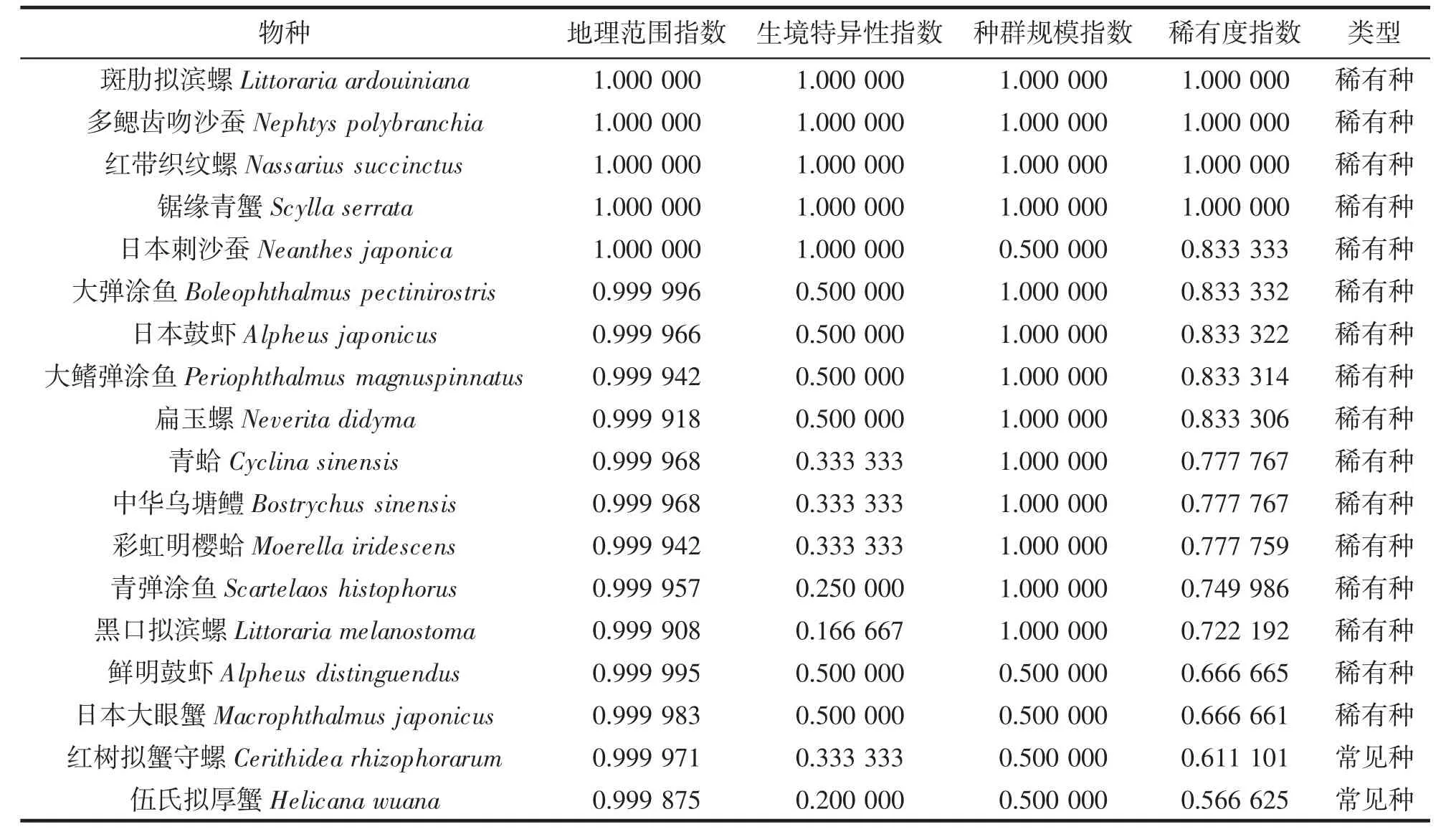
表1 沿浦湾红树林大型底栖动物稀有度指数Tab.1 Rare index of macrobenthicin the mangroves of Yanpu Bay
在16 种稀有种中,腹足纲、软甲纲与硬骨鱼纲种类数均占稀有种种类数的25%,多毛纲与双壳纲种类占稀有种种类的12.5%。在14 种常见种中,腹足纲、软甲纲种类占常见种种类的35.72%,多毛纲、双壳纲、硬骨鱼纲与革囊星虫纲种类均占常见种种类的7.14%。
2.2 稀有种与常见种对物种多样性的影响
通过模拟稀有种、常见种丧失和稀有种、常见种均保留的等3 种情景分析稀有种与常见种对物种多样性的影响。在模拟稀有种丧失后发现,Margalef 丰富度指数呈极显著下降趋势(P<0.01),而Shannon-Wiener指数、Pielou 均匀度和Simpson 指数变化趋势并不显著(P>0.05);在模拟常见种丧失后发现,Shannon-Wiener 指数呈极显著下降趋势(P<0.01),Pielou 均匀度指数呈极显著上升趋势(P<0.01),Margalef 丰富度和Simpson 指数的变化均不显著(图2)。
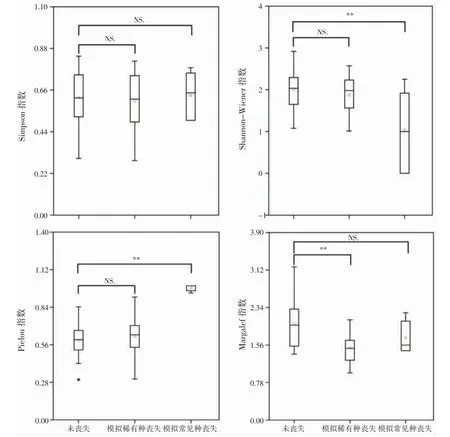
图2 沿浦湾红树林大型底栖动物稀有种与常见种对物种多样性指数影响Fig.2 Effects of rare and common species of macrobenthos on species diversity index in the mangroves of Yanpu Bay
2.3 稀有种、常见种与全部物种的物种多度模型拟合
物种多度模型拟合结果见表3,在情景1 中,稀有种与常见种均保留。K-S 检验结果显示几何级数模型与断棍模型P 值均小于0.05,表明这2 个模型拒绝假设,其余模型均通过。AIC 结果表明,泊松对数正态模型为最优模型,表明红树林大型底栖动物群落处于非干扰状态。
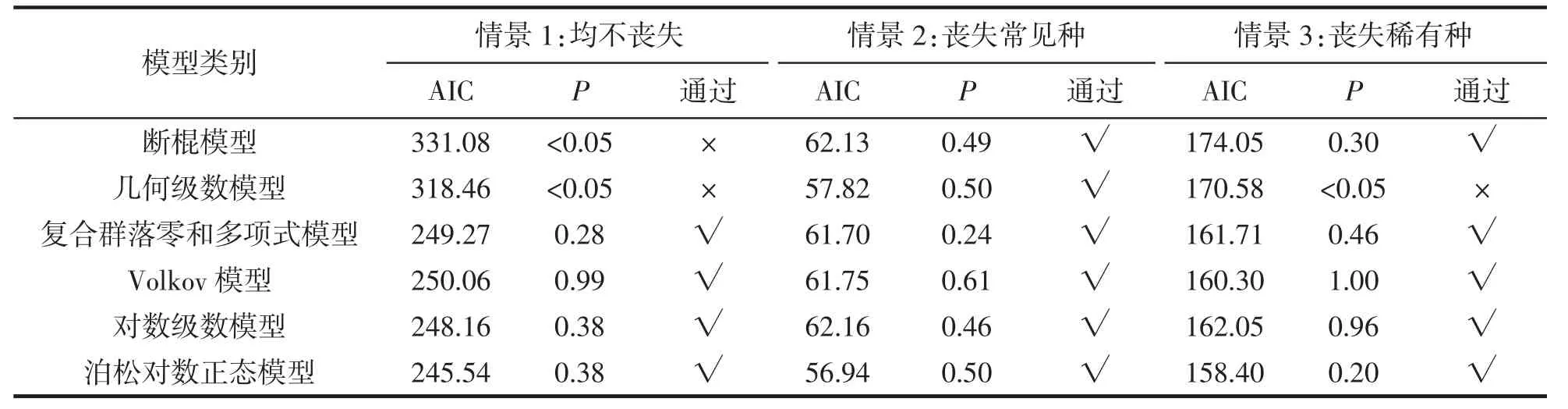
表3 沿浦湾红树林大型底栖动物保留稀有种和常见种、稀有种丧失与常见种丧失物种多度分布模型拟合优度Tab.3 The goodness of fit of species abundance distribution model of rare and common species,rare species loss and common species loss of macrobenthos in the mangroves of Yanpu Bay
在情景2 中,模拟常见种丧失。K-S 检验结果显示6 个模型P 值均大于0.05,表明这6 个模型均通过假设。AIC 结果表明,泊松对数正态模型与几何级数模型均为最优拟合模型(AIC 差值小于2),表明稀有种群落受生态位过程主导。
在情景3 中,模拟稀有种丧失。K-S 检验结果显示几何级数模型P 值小于0.05,表明几何级数模型拒绝假设,其余模型均通过。AIC 结果表明,泊松对数正态模型与Volkov 模型均为最优拟合模型(AIC 差值小于2),表明常见种群落受中性过程主导。
3 讨论
3.1 基于稀有度指数对稀有种与常见种进行识别
在以往稀有种研究中多采用多度、频率、相对重要性指数等作为稀有种的识别指标[38]。而在不同的生物群落其指标也有所差异,如在植物群落中主要是树高、盖度与多度[39-40];而对于移动能力较强游泳动物,仅采用多度或频度难以客观反映群落中物种的优势度和稀有度,常使用包含丰度、生物量及频度的相对重要性指数(IRI)进行识别[41]。然而在空间尺度较大或者物种丰度较多的情况下,使用多度、频率以及相对重要性指数识别稀有种仍存在不足,难以兼顾物种地理分布范围、栖息地特异性与种群大小,进而影响稀有种的准确识别。为此,Maciel 结合物种的地理分布范围、栖息地特异性和种群大小开发出“rrindex”工具包,可有效提高稀有种识别的准确性。
基于“rrindex”工具包识别沿浦湾红树林大型底栖动物稀有种及常见种发现,常见种以蟹类和螺类为主,而稀有种主要为多毛类和弹涂鱼类。红树植物凋落物及其碎屑可为蟹类、螺类等植食性动物提供丰富的食物来源。例如,相手蟹科动物是红树凋落物的主要摄食者,其对凋落物的消耗速率是微生物分解速率的75 倍[42-43]。王瑁等[44]研究发现红树蚬Geloina coaxans 主要取食沉积物表层凋落的红树植物碎屑,树皮和叶片的食物贡献率为26%~50%。此外,红树植物的生长不仅提高生境异质性[45],而且减弱高温对底栖动物的影响[46],为林内的蟹类和螺类提供适宜的栖息环境,故上述物种多为红树林常见种。沙蚕与弹涂鱼经济价值较高,是当地重要采捕对象,高强度的采捕使其种群规模和分布空间不断减小,稀有化加剧,故多为稀有种。然而,稀有种具有分布区狭窄、种群规模小以及受扰动敏感的特征,当受到扰动时对群落的影响相对较小,因此大型底栖动物群落在物种多度分布模型拟合结果中呈现稳定性较好的现象。
3.2 稀有种和常见种丧失对物种多样性的影响
在局域群落中常见种具有丰度高且分布广泛的特点,在丰度模型中起到主导作用[47]。常见种的丧失使得群落物种丰度急剧下降,故Shannon-Wiener 多样性指数出现极显著下降(P<0.01);而稀有种虽种类众多,但规模小,故稀有种的丧失对群落物种丰度变化影响较小,不会导致Shannon-Wiener 多样性指数显著降低(P>0.05)。Shannon-Wiener 多样性指数可反映群落受干扰程度,扰动越大其值越低[48]。稀有种是群落中易受扰动影响的物种,轻微的扰动即可使其在局域群落中丧失[49];常见种多为广生态位种,对扰动适应能力较强[50],当常见种丧失后,表明群落已受到较大干扰,Shannon-Wiener 多样性指数将显著降低,这与该研究结论一致。
扩散限制作用与生境异质性可能是物种呈现聚集分布的主要原因[51]。该研究中,多度模型拟合结果显示中性过程在常见种群落构成中占据主导地位,表明扩散限制作用对常见种分布的调控作用较强;红树林秋茄的种植提高生境异质性,促使常见种的斑块分布。常见种丧失将会进一步降低物种空间聚集分布,因此Pielou 均匀度指数将呈极显著上升趋势。Margalef 丰富度指数是反映群落物种丰富程度的关键指标[52]。稀有种种类众多,在Margalef 丰富度指数中占有较大比重且有强烈影响[9,53]。稀有种丧失后,造成群落物种种类数急剧下降,因此Margalef 丰富度指数将呈极显著下降趋势。
3.3 稀有种和常见种丧失对物种多度分布格局的影响
笔者的研究中常见种有尖锥拟蟹守螺Cerithidea largillierti、长足长方蟹Metaplax longipes、微黄镰玉螺Lunatia gilva 等,这些物种多为广生态位种,对资源利用能力较强[50],因此常见种在种间生态位分化作用较弱,而扩散限制对群落物种组成的影响增强[54],从而中性过程在常见种群落构成中占据主导地位。而稀有种恰与常见种模式相反,生态位宽度较窄[25],资源利用率低,使得种间的生态位分化作用较强[55],故生态位过程在其物种多度分布格局中起主导作用。陈圣宾等[56]认为几乎没有完全受扩散限制或生态位限制单独限制作用的群落,大部分群落处于从完全受扩散限制到完全受生态位限制的谱带上。稀有种与常见种的多度分布格局受不同生态过程的影响也间接说明生态位过程与中性过程并非相互矛盾,只不过是作用强度不同。因此,生态位过程和中性过程相互补充、相互作用,共同调控群落物种多度分布格局。
沿浦湾大型底栖动物物种多样性主要受稀有种物种数量与常见种分布特点影响,群落分布格局的形成受到稀有种间生态位分化作用与常见种间扩散限制作用共同影响,两者相互作用共同维持大型底栖动物群落物种共存。但鉴于单使用多度模型无法明确其生态学机制背后具体成因,后续研究建议引入环境因子、功能多样性、β 多样性等研究角度,以验证沿浦湾红树林大型底栖动物群落多样性维持机制。
