陆地棉主要株型性状关联分析及优异等位基因挖掘
2024-04-17王娟王新田琴马晓梅周小凤李保成董承光
王娟 王新 田琴 马晓梅 周小凤 李保成 董承光
(新疆农垦科学院棉花研究所,石河子 832000)
新疆是我国最重要的优质商品棉生产基地[1],2022 年棉花种植面积为2 496.9 千hm2,总产量为539.1 万t,分别占全国的83.2%和90.2%(国家统计局,2022)。新疆棉花机械化程度高,尤其新疆生产建设兵团棉花机采率达94.2%。株型是影响棉花机械化和产量的关键因素,棉花株型因子主要包括株高、始果节高、始果节位、果枝数、果枝类型等性状[2]。合理的株型有利于提高种植密度、提升光合效率及降低机械采摘过程中的产量损失等。
近年来,棉花株型育种已成为新疆棉花育种中主要育种目标性状。棉花株型性状均属于复杂的数量性状,受基因型和环境的共同控制[3-5],利用传统的育种方法改良株型的难度较大,利用分子标记技术结合连锁分析和关联分析方法,可以发掘数量性状基因位点,是解析植物数量性状基因的重要方法[6]。采用构建分离群体结合分子标记方法,研究学者定位了多个株型性状的QTL[7-17],随着分子生物学技术和统计分析技术的不断发展,通过自然群体结合关联分析方法,多个株型相关性状优异等位变异进一步被发掘定位[18-23],为棉花株型性状的分子育种研究奠定了基础。
随着新疆棉花机械化采收的不断普及,对适宜机采棉花株型性状提出更高的要求,迫切需要科研人员加大对棉花株型分子育种的研究,以加速品种选育速度,提高选育效率。尽管前人在株型性状基因挖掘方面做了大量工作,但目前仍缺少主效QTL应用于棉花株型育种中。
本研究以我国不同棉区的403 份陆地棉品种资源为主要研究对象,在新疆机采棉栽培模式下,通过对4 个主要株型性状因子的多年多点性状调查鉴定,并结合SSR 标记的全基因组扫描关联分析研究,进一步发掘以上性状的关联位点及优异等位基因,为新疆棉花株型性状分子育种提供材料和标记来源。
1 材料与方法
1.1 材料
选取403 份陆地棉种质资源为试验材料,来自新疆农垦科学院棉花研究所多年来收集保存的国内外种质资源,其中来自于我国西北内陆棉区的有199 份,占总材料份数的49.4%;来自于黄河流域棉区的有87 份,占总材料份数的21.6%;来自于长江流域棉区的有63 份,占总材料份数的15.6%;来自于北部特早熟棉区的有29 份,占总材料份数的7.2%;来自于国外的有25 份,占总材料份数的6.2%(具体材料名称及来源详见附表1)。
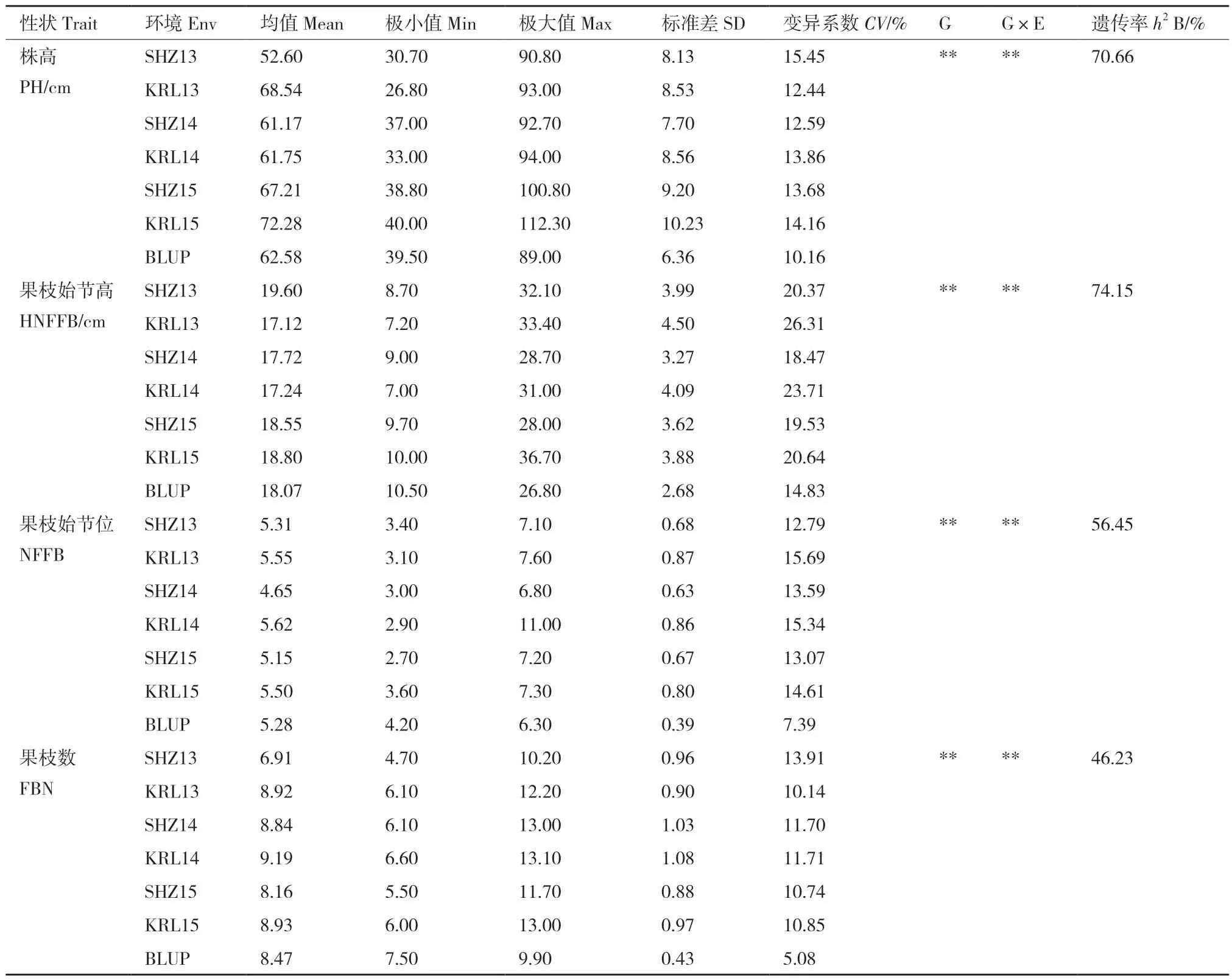
表1 不同环境下4 个主要农艺性状的统计分析Table 1 Descriptive statistics for main agronomic traits in different environments
1.2 方法
1.2.1 性状调查及数据处理 试验于2013 年、2014年、2015 年连续3 年分别在新疆农垦科学院棉花研究所北疆育种试验站(石河子)与南疆育种试验站(库尔勒)2 个不同环境种植上述403 份材料,各试验点田间试验设计采用完全随机区组设计,3 次重复,双行区,平均行距42.5 cm,平均株距10 cm,采取人工点播种植,膜下滴灌栽培,从出苗到吐絮全程化控6 次,平均每亩用缩节胺20 g,7 月10 日采用人工打顶方式打顶,其他田间管理措施以当地常规方法进行。分别在棉花成铃后期在各重复中每小区选中间连续10 株调查株高(plant height, PH)、果枝始节高(height of first fruit branch node, HNFFB)、果枝始节位(the first fruit branch node, NFFB)、果枝数(fruit branch number, FBN)4 个主要株型性状,各环境分别定义为:SHZ13、KRL13、SHZ14、KRL14、SHZ15和KRL15。采用R语言的Ime4软件包,计算获得各环境各性状表型的最佳线性无偏预测值(best linear unbiased prediction, BLUP),用于后续关联分析。
1.2.2 基因型鉴定及关联分析 参考Zhao 等[24]发表的棉花四倍体分子遗传图谱,在每条染色体上平均每隔10 cM 左右选取一个SSR 分子标记,并进一步结合前人已发表的在陆地棉中多态性较好的SSR 标记结果[25-27],选取均匀分布在26 条染色体的560 对引物对群体进行PCR 扩增分析。SSR 标记引物序列来自CMD(cotton marker database)数据库(http://www.cottonmarker.org),标记名称中的大写字母,如BNL、JESPR、HAU、NAU 等表示引物的来源,引物名称后的小写字母“a”“b”“c”“d”等代表标记的多态性位点,且位点顺序依次按分子量由大到小排列。群体结构分析、连锁不平衡分析及关联分析详见文献[28]。
1.2.3 优异等位变异挖掘 优异等位变异参考钱能[29]的方法计算,ai =(Σxij/ni)-(ΣNk/nk)。其中,ai 代表第i 个等位变异的表型效应值,xij 为携带第i 个等位变异的第j 材料性状表型测定值,ni 为具有第i 等位变异 的材料数,Nk 为所有材料的表型测定值, nk 为材料数。若ai 为正, 则认为该等位变异为增效等位变异,反之为减效等位变异。最终获得与表型性状显著关联的位点等位变异、表型效应及典型品种。
2 结果
2.1 表型性状分析
403 份陆地棉种质材料的4 个主要农艺性状3年两点6 个环境的表型值及BLUP 值统计结果如表1 所示。结果显示,4 个性状6 环境平均变异系数分别为,株高13.70%、果枝始节高21.51%、果枝始节位14.18%、果枝数11.51%;联合方差分析结果表明,4 个性状的基因型、基因型与环境互作变异方差均达极显著水平;4 个性状的广义遗传率变化范围介于46.24%-74.15%,其中最高的为果枝始节高,最低的为果枝数。4 个性状的BLUP 结果显示,均符合正态分布规律(图1)。
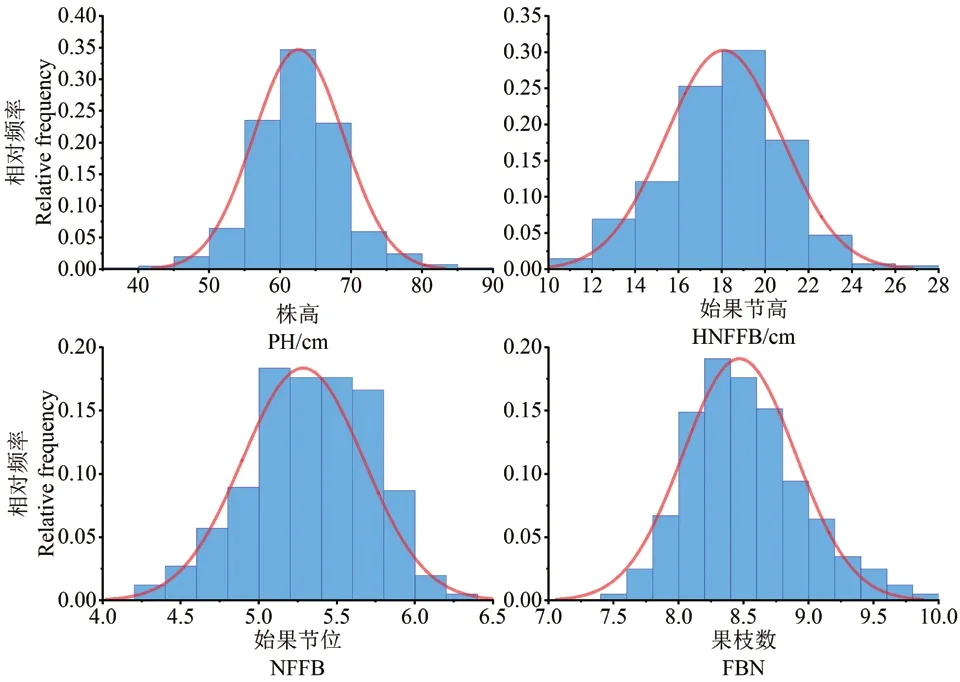
图1 4 个主要农艺性状的BLUP 值频率分布图Fig.1 Frequency distributions of the BLUP of four main agronomic traits
2.2 基因型分析
对403 份种质材料先按照不同棉区的生态来源信息选取遗传差异较大的48 份材料,结合选取的560 对SSR 引物进行小群体的标记鉴定。最终获得多态性明显、易于确认的SSR 标记201 对。201 对标记分别分布在棉花基因组的26 条染色体上(A亚组分布91 对,D 亚组分布110 对),其中标记最多的是D05 染色体(18 对),最少的是A02 染色体(3 对)。进一步对201 对SSR 标记进行了大群体分子标记鉴定。结果(表2)表明,201 对SSR 标记位点在403 份材料中共检测到394 个等位变异,每个标记的平均等位变异为1.96 个,变幅为1-4;每个标记的平均基因型为4 个,变幅为2-14;平均等位基因频率为0.563,变幅为0.501-0.923;平均遗传多样性指数为0.556,变幅为0.142-0.699;平均多态信息含量为0.483,变幅为0.132-0.677。

表2 201 对多态性SSR 标记在403 份材料中的引物多态性信息Table 2 Polymorphism information of 201 SSR loci in 403 experiment materials
2.3 关联分析
利用TASSEL 5.0 软件的混合线性模型MLM 程序,将上述394 个位点的等位变异分别与各个性状的6 个环境的表型值及BLUP 值进行关联分析。为最大程度地避免虚假关联的出现,研究结果只考虑能同时在BLUP 结果(P < 0.01)和2 个以上环境中检测到的显著(P < 0.05)关联的位点。结果表明,4 个主要农艺性状共检测到38 个位点(表3),与株高(PH)关联的位点共16 个,不同环境中表型变异解释率0.99%-5.00%,除BLUP 外,能同时在2个环境中检测到的位点有7 个、3 个环境中检测到的有2 个、4 个环境中检测到的有1 个、5 个环境中检测到的有5 个,BNL3089a 能同时在6 个环境中全部检测到;与果枝始节高(HNFFB)关联的位点共6 个,不同环境中表型变异解释率0.98%-3.65%,除BLUP 外,能同时在2 个环境中检测到的位点有2 个、3 个环境中检测到的有3 个、4 个环境中检测到的有1 个;与果枝始节位(NFFB)关联的位点共11 个,不同环境中表型变异解释率0.97%-3.93%,除BLUP 外,能同时在2 个环境中检测到的位点有5个、3 个环境中检测到的有3 个、4 个环境中检测到的有2 个、5 个环境中检测到的有1 个;与果枝数(FBN)关联的位点共5 个,不同环境中表型变异解释率1.00%-2.63%,除BLUP 外,能同时在2 个环境中检测到的位点有2 个、3 个环境中检测到的有2个、4 个环境中检测到的有1 个。
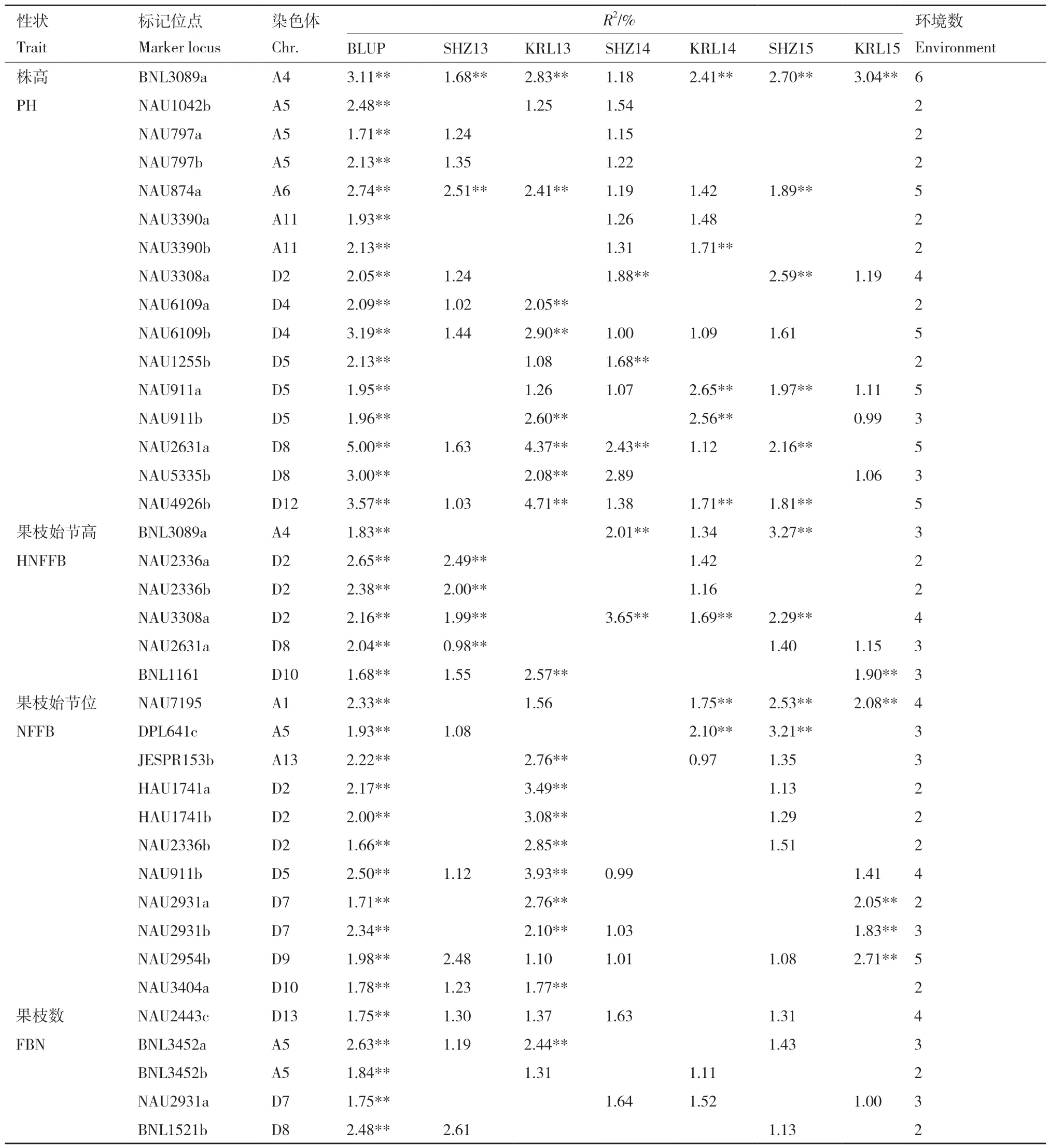
表3 与主要农艺性状关联的标记位点及其在多环境下表型变异的解释率Table 3 Marker loci associated with major agronomic traits and their explained portions of phenotypic variation in multiple environments
2.4 优异等位变异及典型材料挖掘
在上述关联分析结果的基础上,对上述标记位点进行等位变异表型效应的计算、统计和分析,并进一步结合育种实际,确定优异等位变异,共发掘含有目标性状的优良载体材料31 份(表4)。有6份材料同时携带多个优异等位变异,其中,同时携带2 个性状的优异等位变异载体材料有4 份,分别为09‑19‑7 含有株高和果枝始节高的优异等位变异(增效效应),BLUP 值分别为81.7 和25.8 cm;中棉所64 含有株高和果枝始节高的优异等位变异(减效效应),BLUP 值分别为44.2 和12.1 cm;DP2824‑092 含有株高和果枝始节高的优异等位变异(减效效应),BLUP 值分别为45.2 和13 cm;惠和28 含有果枝始节高和果枝始节位的优异等位变异(增效效应),BLUP 值分别为26.3 cm 和6.3。而德夏棉1号和中221 两个材料则同时携带株高、果枝始节高和果枝始节位等3 个性状的优异等位变异(减效效应),BLUP 值分别为39.5 cm、10.5 cm、4.2 和45.6 cm、11.3 cm、4.2。

表4 与农艺性状显著关联的位点等位变异对应的表型效应Table 4 Phenotypic effect corresponding to locus allelic variations significantly associated with agronomic traits
3 讨论
3.1 开展陆地棉株型育种的重要性
棉花具有多年生、无限生长的特性,具有复杂的分枝模式,侧枝较长、松散的株型不利于棉花单产提高和机械化采摘[30]。棉花株型性状的遗传解析是棉花理想株型育种的理论基础。对于新疆机采棉而言,要求株型紧凑,株高80-90 cm,果枝始节位高[21,31],才能达到农机农艺融合,提高机械采摘质量,减少原棉杂质污染,从而提高机采棉经济效益。本研究利用SSR 分子标记技术,对403 份陆地棉种质材料进行4 个主要株型性状因子的关联分析,发掘株型相关遗传位点及优异等位变异,为加强新疆棉花株型分子育种基础研究,以及加快新疆机采棉育种速度提供理论基础。
3.2 影响关联分析结果的主要因素
近年来,由于分子标记技术及统计分析技术的不断发展,使关联分析在植物研究中得到广泛应用,成为近年来研究的热点,本研究利用201 对SSR 标记位点在403 份材料中共检测到394 个等位变异,其中38 个等位变异与棉花主要株型性状相关联,为陆地棉株型分子育种提供了较好的理论基础。对关联分析而言,精准的表型性状值及广泛分布不同基因组的多态性标记可为关联分析研究提供更为准确的结果。本研究在表型鉴定上,采用多年多点多重复的表型鉴定,且在统计分析中进一步采用最佳无偏线性预测的BLUP 方法,提高了表型结果的准确性;在标记选择上选取均匀分布在不同染色体上的标记,进一步提高了检测效率。但要进行更为有效的全基因组关联扫描,按照陆地棉染色体图谱5 000 cM 的遗传距离计算,结合连锁不平衡衰减程度,需要1 000 个左右的多态性位点[32]。本研究201 个多态性标记共产生394 个多态位点,仅能达到粗略的基因组扫描,如要对控制重要育种目标性状基因进行精细定位,仍需增加更多标记。
3.3 多个关联位点的一因多效性分析
基因连锁和多效性是生物体遗传学研究中常见的现象。本研究检测到的38 个位点中,其中有5个标记位点同时与多个性状相关联,NAU2336b 同时与果枝始节高和果枝始节位相关联,NAU2931a同时与果枝始节位和果枝数相关联,NAU2631a、NAU3308a 同时与株高和果枝始节高相关联,NAU911b 同时与株高和果枝始节位相关联。说明这些标记具有一因多效性,有同步改良多个育种目标性状的潜力,以实现多个育种目标性状的聚合育种研究。自然和人工表型选择可以解释棉花驯化过程中的基因连锁和一因多效性现象。另外,本研究检测到38 个关联位点中发现,有4 个位点与前人通过连锁分析定位得到的株型及相关性状的QTL 结果一致(表5),说明这些标记具有较好的可重复性和稳定性。上述标记可为进一步理解棉花株型性状的遗传机理提供理论依据,并可为后期棉花株型性状的分子标记辅助选择育种提供实践指导。

表5 本研究检测到的主要育种目标性状关联位点与前人研究的比较Table 5 Comparison of QTL associated with main breeding target traits detected in this study to the previous studies
3.4 优异等位变异及相应载体材料的应用
挖掘与育种目标性状相关的优异种质是棉花育种研究的重要内容之一。本研究在关联分析基础上,挖掘得到含有目标性状的优良载体材料31 份。结合育种实际,4 个性状可根据不同的育种目标确定其相应的优异等位变异,例如,对株高而言,如进行矮秆育种,可选择表型效应为负值的,如NAU911b是减效(‑7.27 cm)表型效应最大的等位变异,典型材料有德夏棉1 号、中221 和中棉所50;对果枝始节高而言,如进行适宜机采的株型育种,可选择表型效应为正值的,如NAU2336b 是增效(+0.33 cm)表型效应最大的等位变异,典型材料有09‑19‑7、恵和28、恵远14‑19。上述材料可作为优异亲本用于棉花株型育种的性状改良。另外,在株高性状的研究中,本研究发掘到11 个表现为株高性状增效效应的优异等位变异,典型材料主要有09‑19‑7、金垦69‑2、Y11、110‑47、C2、10‑1,发现这6 份材料均来源于西北内陆棉区,分析其原因可能为来源于其他棉区的材料由于对缩节胺较为敏感,对棉花生长有一定的抑制作用,表现为株高低于80 cm,不适宜机械采收。因此建议长江流域、黄河流域等棉区品种在新疆棉区种植应降低缩节胺的使用量。
4 结论
在陆地棉品种(系)群体中共鉴定到38 个与4个株型性状关联的标记位点,其中,有5 个标记位点同时与多个性状相关联;发掘出31 份含有优异等位基因的典型材料,其中6 份材料同时携带多个优异等位基因。
文章所有附表数据请到本刊官网下载(http://biotech.aiijournal.com/CN/1002‑5464/home.shtml)。
