基于林木株数与竞争指数的马尾松退化林种群结构及数量动态分析*
2024-04-12申鹏白萍朱洲柴宗政
申鹏,白萍,朱洲,柴宗政
(贵州大学 林学院,贵州 贵阳 550025)
植物种群结构可以表征植物与环境之间的相互关系,种群数量动态能够解释种群的生存规律;研究植物种群结构及数量动态,对种群的保护与发展具有重要意义[1-2]。以往林木种群结构和数量动态研究多采用龄级结构、静态生命表、生存分析、动态指数和时间序列等方法,以种群数量为基础[3-6],却忽视了竞争对林木种群结构及数量动态的影响。自然界中,植物种群内部和群落之间普遍存在竞争关系[7]。植物间的竞争对植物个体生长、形态结构和种群空间分布产生重要影响[8-9]。对森林而言,林木间的竞争关系对林木的生长发育、形态结构、分布格局和演替更新都具有重要影响。研究林木种群竞争关系可以有效反映种群结构与动态,对维持种群结构稳定性和调节动态也具有重要作用[10]。因此,种群个体数量以及种内、种间竞争强度均与其种群结构、发展动态密切相关。但是,在以往林木种群结构与数量动态研究中,大多仅对林木株数特征或者竞争关系单方面分析,而很少见到有综合两者来分析种群动态的报道。
马尾松(Pinusmassoniana)是我国中部及南部地区分布最广的树种之一[11],也是我国亚热带退化生态系统中恢复与重建的乡土树种和先锋树种[12]。近些年来,我国南方地区大面积营造了马尾松纯林,但其树种组成单一、林分密度大、层次结构简单,林分抵御有害生物的能力差[13],生产力和效益不高[14]等问题突出,部分马尾松林生长逐渐衰退,呈现低质、低效状态,严重威胁林地的可持续发展[15]。因此,研究马尾松退化林种群结构及动态变化,对了解其退化现状并提出修复策略具有重要指导意义。
本研究选择贵州省紫云苗族布依族自治县国有浪风关林场作为研究区域,以典型马尾松退化林为研究对象,通过结合林木株数和Hegyi竞争指数,运用动态指数、时间序列预测等方法,综合分析研究区马尾松退化林种群结构及数量动态,以期为该林场马尾松退化林提质增效、科学管理和实现可持续良性发展提供理论依据。
1 研究区概况与研究方法
1.1 研究区概况
紫云苗族布依族自治县国有浪风关林场(106°05′~106°42′E、25°44′~40°11′N),位于贵州苗岭山脉中段、珠江流域红河水系,地势西南高、东北低,地形以山地为主,平均海拔1 185 m。该地区属中亚热带季风湿润气候,年均气温15.3 ℃,年均总积温4 559.5 ℃,年均降水量1 337.1 mm,年均无霜期288 d。土壤以砂页岩发育的黄壤为主,另有少量黄棕壤,土壤肥沃,适宜植物生长。乔木林主要以马尾松、湿地松(Pinuselliottii)、杉木(Cunninghamialanceolata)等人工针叶林为主。其中,马尾松林面积987.07 hm2,蓄积228 050.09 m3,面积较大且分布集中,约占林地面积91%。该区域部分马尾松人工林由于初植密度过大,立地条件差,加之在中幼龄阶段未及时抚育间伐,导致林木生长受到抑制,林木枯死严重,退化现象明显。
1.2 研究方法
1.2.1 样地设置与种群调查
以浪风关林场为研究区域,在全面踏查的基础上,选择马尾松退化林分布集中区域,采用典型抽样调查,共计设置20个样地,面积为20 m×20 m。对样地内所有胸径(DBH) ≥ 1 cm的乔木树种进行每木检尺,实测每株个体的胸径、树高、冠高、枝下高、冠幅、林木质量、病虫害状况、生活力、损伤状况,并详细记录样地编号、地理位置、样地面积、调查日期,同时测定海拔、地形、坡向、坡度、土壤类型、干扰状况、经营利用状况及生境概况。
1.2.2 年龄结构的确定和划分
马尾松平均个体寿命较长,研究马尾松的种群生物学特征相对困难。依据董鸣[16]的有关研究,同一树种在相同环境下,龄级和径级反应规律相一致。因此,我们采用径级结构代替年龄结构的分析方法来研究马尾松退化林种群结构特征。考虑到马尾松的生长规律和样地调查实际情况,采用上限排外法,将1 cm≤DBH<5 cm的植株定义为Ⅰ龄级,其后以4 cm为步长增加1龄级,即5 cm≤DBH<9 cm定义为Ⅱ龄级,依此类推,将该研究区的马尾松退化林共划分为12个龄级。
1.2.3 基于林木株数和竞争的种群动态分析理论基础
植物种群动态特征是种群内植物个体的生理特点与生境之间相互作用的结果[17]。通常,一个稳定的林木种群结构,其个体数量分布呈“倒J”型[18],即林木株数随着龄级的增大而逐渐减小,从种群生态学角度上来说,“倒J”型的种群结构意味着种群具有良好的繁殖能力和增长潜力[19-20],更有利于种群的更新与繁衍[21-22];竞争作为林木种群重要特征之一,随着胸径的增大而减小,也呈现“倒J”型分布,因为幼龄个体承受更大的竞争压力,随着龄级的增大,林木个体竞争能力越强,受到的竞争压力越小,多项有关林木竞争的研究均能证实这一结论[23-27]。可见,基于林木株数和竞争与龄级间具有相同的反应规律。因此,本研究尝试在传统以林木株数研究种群动态方法的基础上,引入竞争指数共同描述种群数量动态变化,以期更加科学完备地揭示种群结构及数量动态规律。
1.2.4 竞争指数的计算
采用Hegyi[28]1974年提出的单木竞争指数模型,其计算方法如下:
①
②
③
式中:Hegyii表示第i株对象木的竞争指数,Dj表示第j个竞争木的胸径,Di表示第i个对象木的胸径,dij表示第i个对象木与第j个竞争木之间的水平距离,n表示对象木的竞争木个数,Hegyi表示总竞争指数,Hegyi_m表示平均竞争指数。
以上竞争指数值越大代表竞争越激烈[29-30]。在计算竞争指数时,样地边缘的林木会缺乏参照树,因而存在边缘效应。为消除边缘效应影响,本研究采用基于拓扑学原理衍生的八邻域平移法进行边缘校正[23,31]。该方法依据拓扑学原理,将原始样地作为基准点,分别在原始样地的上、下、左、右、左上、左下、右上、右下共8个方向用平移的方式复制原样地,将原样地扩大为9个相同样地组成的大样地,以初始样地(20 m × 20 m)为核心区,将其内部林木作为参照树,并在样地周外围选取5 m的宽度作为缓冲区,处于缓冲区的林木只作为核心区林木的相邻木处理,从而达到消除边缘效应的目的。本研究中的竞争指数为种内平均竞争指数。
1.2.5 种群数量动态量化分析方法
参考陈晓德[32]的种群动态量化分析方法,描述马尾松种群动态变化。计算公式如下:
④

⑤
⑥
⑦

1.2.6 时间序列模型预测
时间序列预测模型近年来在植物种群动态研究中大量应用,是预测种群未来发展趋势的重要手段[33]。本研究采用一次移动平均法对马尾松退化林种群的年龄结构动态进行预测,具体算法如下:
⑧

1.3 数据处理与分析方法
通过实地调查得到马尾松退化林种群相关数据,应用Excel 2010进行数据录入及存储,利用Origin 2022进行数据分析及图表绘制。
2 结果与分析
2.1 马尾松退化林种群结构特征
共计调查马尾松林木1 297株,包括12个龄级(图1)。马尾松种群个体数量随着龄级的增大呈先上升后下降的单峰趋势,Ⅰ 龄级个体数量只有52株,远低于Ⅱ、Ⅲ 龄级个体数,在Ⅲ 龄级后,个体数随着龄级增大而逐渐减少。由于I龄级个体过少,表明其天然更新能力较差。此外,种群竞争指数整体呈逐级下降的趋势,Ⅰ龄级数值最高,Ⅻ龄级最低,分别为2.383和0.098;Ⅶ龄级和Ⅸ龄级时,竞争指数出现波动,明显高于相邻龄级,分别为0.918和0.946;总体来看,马尾松种群以小径级个体为主,各龄级株数分布不合理,天然更新能力不足;Ⅶ龄级和Ⅸ龄级种内竞争过大,种群结构不稳定,呈衰退趋势。

图1 马尾松退化林种群数量及竞争指数
2.2 马尾松退化林种群动态变化
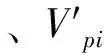

表1 基于林木株数和竞争指数的马尾松退化林种群数量动态变化指数
2.3 未来种群数量与竞争强度预测
基于不同龄级林木株数和竞争指数,应用移动平均法,预测2、4、6和8龄级后种群数量和竞争指数的动态变化。由图2可知,马尾松种群株数在未来2、4龄级时间内,Ⅴ~Ⅻ龄级个体数呈现增加趋势;经过6、8龄级时间后,Ⅸ~Ⅻ龄级个体数也开始不断增加,说明随着时间推移,种群中老龄级个体数量能够得到一定的补充。但经过2龄级时,Ⅱ~Ⅲ龄级数量呈现下降趋势,其中Ⅱ龄级个体数量下降了40.1%,说明由于Ⅰ龄级个体本身数量较少,将在短期内严重影响种群正常更新。从竞争指数变化上来看,经过2、4龄级时间后,Ⅶ和Ⅸ龄级种内竞争指数还存在波动,在未来6龄级以后竞争指数开始随龄级增大而逐渐减小,符合种群竞争理想规律,种群整体趋于稳定。

图2 基于林木株数和竞争指数的马尾松退化林种群时间序列分析
3 讨论与结论
3.1 讨论
3.1.1 马尾松退化林种群结构不稳定的原因分析
林木种群结构特征是种群内部林木个体发展过程中的具体表现,反映了林木种群自身的发展特性和对外界环境的适应能力[34]。幼龄个体数量是种群更新发育的物质基础,对维持种群稳定性具有重要作用[35]。本研究中,马尾松种群龄级结构表现为单峰状,属衰退型,种群结构不稳定。Ⅰ龄级株数远低于Ⅱ、Ⅲ龄级,表明种群更新能力有限。从竞争指数来看,马尾松种群的竞争指数总体随龄级的增大而逐渐减小,幼龄个体竞争指数较高,尤其Ⅰ龄级竞争指数最高,竞争最为强烈,Ⅶ龄级和Ⅸ龄级内部竞争指数高于相邻龄级,不符合种群竞争规律。尽管通过现场踏查和资料显示,马尾松具有一定的天然更新能力,能在养分贫瘠的土壤中生长,且低覆盖度有利于马尾松种子发芽,但是,由于研究区马尾松林初植密度过大,4 444株/hm2(株行距1.5 m×1.5 m),从栽植后任其自由生长,未进行任何抚育经营,造成林分密度过大、郁闭度过高、种内竞争激烈,加之马尾松成林后占据主林层,对光照和水分的竞争具有绝对优势,林下荫蔽条件不利于种子的萌发,幼苗个体竞争力较弱,在生长过程中易受到环境因素的影响处于不稳定状态,个体死亡率高[36],强烈的环境筛选作用延缓了种群更新进程[37]。此外,Ⅶ龄级和Ⅸ龄级林木株数相对较多,龄级内部竞争强度过大且明显高于相邻龄级,主要是由于主林层林分密度过大,Ⅶ龄级和Ⅸ龄级内林木竞争激烈,强烈的竞争导致生境提供给马尾松的营养物质严重不足[38],造成研究区枯立木较多,林木枯死现象明显。可见,幼苗幼树匮乏和成树个体的死亡,是马尾松退化林种群结构不合理,退化现象严重,难以持续稳定发展的主要原因。
3.1.2 马尾松退化林种群动态变化与发展趋势分析
3.1.3 马尾松退化林种群保护与调控对策
研究区马尾松退化林种群结构不合理,难以持续稳定发展。其主要原因是初植密度过大,栽植后任其自由生长,未进行任何抚育经营,导致竞争激烈,退化现象明显,幼苗幼树匮乏以及成树个体死亡严重制约该种群的天然更新和种群发展。建议采取以下措施,提高幼苗存活率和减少成树死亡率,促进退化林恢复和改善:(1)对主林层间伐,通过“去劣留优、去密留疏、去针留阔”的方式,减小林分密度,改善林内光照,促进种群天然更新,同时伐除竞争强度较大的Ⅶ龄级和Ⅸ龄级的部分林木,减缓竞争。间伐可以改变林分组成结构,同时加速自然更新[39],促进保留木的生长[40]。间伐还可以改变微环境因素,如光照、温度和土壤性质等[41],这些条件的改变都可能增大林下幼苗的存活率。(2)对幼龄级个体进行保护性经营,通过伐除周边影响其生长的灌草植被,改善生存环境,提高幼苗幼树的存活率。(3)通过林下补植的方式补充马尾松幼龄级个体,有效弥补前期幼苗幼树因生长缓慢、存活率不高造成的数量严重不足[42],增强种群结构稳定性。
3.1.4 未来研究方向
传统研究种群结构及数量动态多采用对种群原始数据匀滑处理进行分析,但匀滑处理数据后可能会掩盖种群数量波动中产生的部分生态现象[43],不能最真实的反映种群原有的特征。因此,本研究没有对原始数据进行匀滑处理,而是直接从数据本身进行分析,更能真实反映种群动态特征。传统种群动态研究仅从种群数量动态变化方面对种群动态进行分析,而竞争作为林分结构动态变化的内在驱动因子[44],在掌握森林结构和动态的演变趋势,揭示种群和群落的发育演替进程中具有重要的生态学意义[45-47]。本研究以林木株数结合竞争指数的多角度、多方位的量化分析方法,在关注种群数量动态变化的同时,从种群现阶段个体间的竞争状况出发,用竞争去揭示种群数量变化的本质,对种群数量动态分析结果进行有效补充,有助于更加准确地解释和预测整个种群的动态变化过程,建议在后续同类研究中可以借鉴推广。
3.2 结论
综上所述,贵州省紫云苗族布依族自治县国有浪风关林场的马尾松林种群天然更新能力不足,导致种群呈现衰退趋势且种群结构不稳定,总体呈增长型种群,但种内竞争较激烈,面临较大的衰退风险。推测在未来6个龄级以后,种群中老龄级株数逐渐增多,幼龄级株数下降,种内竞争强度逐渐趋于稳定。建议采取主林层间伐配合林下割灌除草和补植的方式来调整种群结构,尽快实现稳定持续发展。
