转抗虫基因107杨对根际和内源细菌群落的影响*
2024-04-12路文然任亚超赵津王进茂郭甫鑫张军
路文然,任亚超,赵津,王进茂,郭甫鑫,张军
(1.河北农业大学 林学院,河北 保定071000;2.河北省林木种质资源与森林保护重点实验室,河北 保定 071000)
转基因林木种植后对土壤微生物的影响是全面系统研究转基因林木对森林生态系统服务功能不可或缺的内容。Edwards等[1-3]认为在植物体内复杂的微生物群落中,存在着一些相互作用的菌群,这些菌群之间紧密联系,协同参与多种重要的代谢过程。因此,通过分析内生菌相对丰度上的相关性,能够了解微生物之间复杂的互作关系,有利于发掘潜在的功能菌群,可更有效地在农业生产中利用微生物资源。综合国内外的研究现状,转基因植物是否会对土壤微生物造成影响尚无定论[4]。金凌波等[5]研究表明,磷高效转基因大豆(Glycinemax)的种植不会对根际土壤可培养微生物产生明显影响。在巴西种植的转Bt 抗虫基因玉米(Zeamays)与非转基因玉米相比,氨氧化细菌和古细菌群落的丰富度差异明显,不过研究表明土壤微生物群落结构主要受土壤类型和季节的影响,而受植物基因型的影响较小[6]。Zhang等[7-8]也认为作物生育期、土壤理化性质和土壤类型等是影响根际微生物结构的主要因素。在转基因杨树(Populus)方面,李霞等[9]对转Bt基因欧洲黑杨(P.nigra)、张雁等[10]对转Bt基因南林895杨、马晓星等[11]对转AtSnRK2C基因杨树、吕秀华等[12]的研究结果中,均未发现转基因杨树对土壤微生物有负面影响。
杨树不仅具有生长速度快、轮伐期短、生命力强、用途广泛等特点,还具有净化空气、涵养水源、保持水土、防风固沙等生态作用[13],杨树已成为解决我国木材供应的重要用材树种,为国家经济建设提供了大量木材,发挥了巨大的经济效益、生态效益和社会效益。土壤被称为世界上最重要的能源之一,土壤生态系统与功能直接关系到整个生态系统健康。在转基因植物的生长周期中,外源基因表达产物主要通过植株残体和根系分泌物进入土壤,根系生长与土壤接触伴随整个转基因植物生长周期[14]。美国 EPA(Environmental Protection Agency)将转基因植物对土壤生态系统影响列为风险评价的重要组成部分,因此,转基因产品释放的生态风险之一就是对土壤生态系统影响的评估。此外,植物内生菌与其宿主在长期进化过程中形成一种共生或互生的关系,对植物的生长发育、病虫害防治、植物抗性、生物修复等方面都有积极的作用,能显著提高植物的产量和高度[15]。大量研究表明,植物内生菌在植物生长发育过程起重要作用[16-22],杨树不同生态位内生菌菌群组成和形成机制尚不清楚,基因转化可能导致受体植物自身一些代谢物的非预期变异而影响体内环境,进而影响植物内生微生物定植。在微生物群落的研究方法中,高通量测序技术克服了平板培养、变性梯度凝胶电泳技术等方法的不足,测序速度快、覆盖度高,能够全面解析复杂的微生物群落[23]。16S r DNA 相对分子量大小适中,突变率小,基于16Sr DNA 的高通量测序技术已被广泛应用于微生物系统进化、分类及多样性研究中。
107杨是由中国林科院张绮纹研究员等从我国1984年引自意大利的“美洲黑杨×欧洲黑杨” (P.deltoides×P.nigra)无性系中选育而成。其材性良好,干型优美,树冠窄,侧枝细,叶满冠,抗病虫,抗风折,是重要的林业资源,在林业生产中具有重要的地位。本研究利用Illumina测序技术,对3 a生未转基因107杨、转Cry1Ac和Cry3A基因107杨高抗株系DB1和DB15树干韧皮部、根系及根际土壤进行16S V4 区测序,解析107杨根际土壤、根部、韧皮部细菌群组成与结构差异,获得全面的微生物信息,探讨转基因杨树对根际土壤及内源细菌群落可能造成的影响,以期为转基因林木产业化的自然生态风险评估提供参考。
1 材料与方法
1.1 样品采集
于2022年7月进行样品的采集,选定2个转基因株系DB1、DB15和对照株系(CK)为样本,每个小区选定3株,3次重复。同时在树高1.5 m处采集树干韧皮部样品,用灭菌手术刀轻轻刮开树皮表面,切取厚1 mm,20×10 cm大小的韧皮部装入无菌塑封袋中,置-80 ℃冰箱中保存备用。于采集韧皮部的同一株系,沿树基部从东、西、南、北4个方向挖掘深度约30 cm,防止挖断主根,待根系暴露后用无菌剪刀剪取须根,将根系周围大块土壤轻轻抖落后将根系及根系附带土壤装入无菌密封袋中。所有采集好的样品置于干冰环境中带回实验室备用。样品信息见表1。

表1 样品及编号
1.2 样品处理
根际土样品:使用灭菌的细软毛刷,轻轻刷掉根系表面的土壤,去除土壤表面毛根并通过1 mm灭菌滤筛,将过筛后的土壤置于50 mL试管中。根系样品:用无菌水清洗根系表面土8次,将洗净的根系放于15 mL PBSE中,使用超声波在50~60 Hz超声30 s,重复2次,用无菌吸水纸吸干根系表面水分后,用无菌手术刀将样品分成1~2 cm片段,装入无菌塑封袋。韧皮部样品:根据研究选取10 g新鲜植物组织,用无菌水冲洗2遍,后置于无菌塑封袋中。采集韧皮部样品组织放于50 mL离心管中并置于液氮中保存,干冰条件运输。以上所有处理好的样品置于-80 ℃保存。
1.3 16S rDNA 测序
16S rDNA为原核生物的核糖体特征编码序列,常用于细菌以及古细菌的物种分类以及进化关系研究。从样本中提取基因组DNA后,用带有barcode的特异引物扩增rDNA的保守区,引物序列信息见表2。

表2 引物序列信息
对PCR扩增产物切胶回收,采用Quanti Fluor TM荧光计进行定量。将纯化的扩增产物进行等量混合,与测序接头连接,构建测序文库,Hiseq2500 PE250上机测序。通过测序得到raw reads之后,过滤低质量reads,然后进行组装和过滤,以保证利用最有效数据聚类,依次进行物种注释、α多样性分析、β多样性分析。在有效分组的情况下进行组间差异比较及差异检验。
2 结果与分析
2.1 聚类分析及稀释性曲线
聚类分析及稀释曲线又称OTU,即分类操作单元。根据不同的相似度水平,对所有序列进行OTU划分,通常来说,如果有效序列大于97%,即定义为一个OTU,每个OTU对应一种代表序列。使用QIIME(version 1.8.0)软件中的UCLUST对序列(Tags)在97%的相似度水平下进行类聚、获得OTU,并基于细菌和真菌分类学数据库对OTU进行分类学注释。方差分析结果显示(图1),不同部位之间转基因DB1、DB15株系与对照株系之间有显著差异,相同部位之间只有转基因DBs15株系与对照株系之间有显著差异。

图1 转基因107杨与对照杨的OTU数量柱状图
稀释性曲线是利用已测得序列中已知的OTU的相对比例,来计算抽取n个(n小于测得 tags序列总数)tags 时的α多样性指数期望值,然后根据一组n值与其相对应的α多样性指数期望值做出曲线。稀释性曲线用于评估序列数量是否足以覆盖所有类群,并间接反映样品中物种的丰富程度。当曲线趋向平坦时,说明测序数量渐进合理,反之则需要增加测序数据。如图2所示:每条曲线代表一个样品,用不同颜色标记。27个样品的稀释性曲线随着测序量的增加,所能得到的稀释性曲线逐渐趋于饱和,表明取样基本合理。
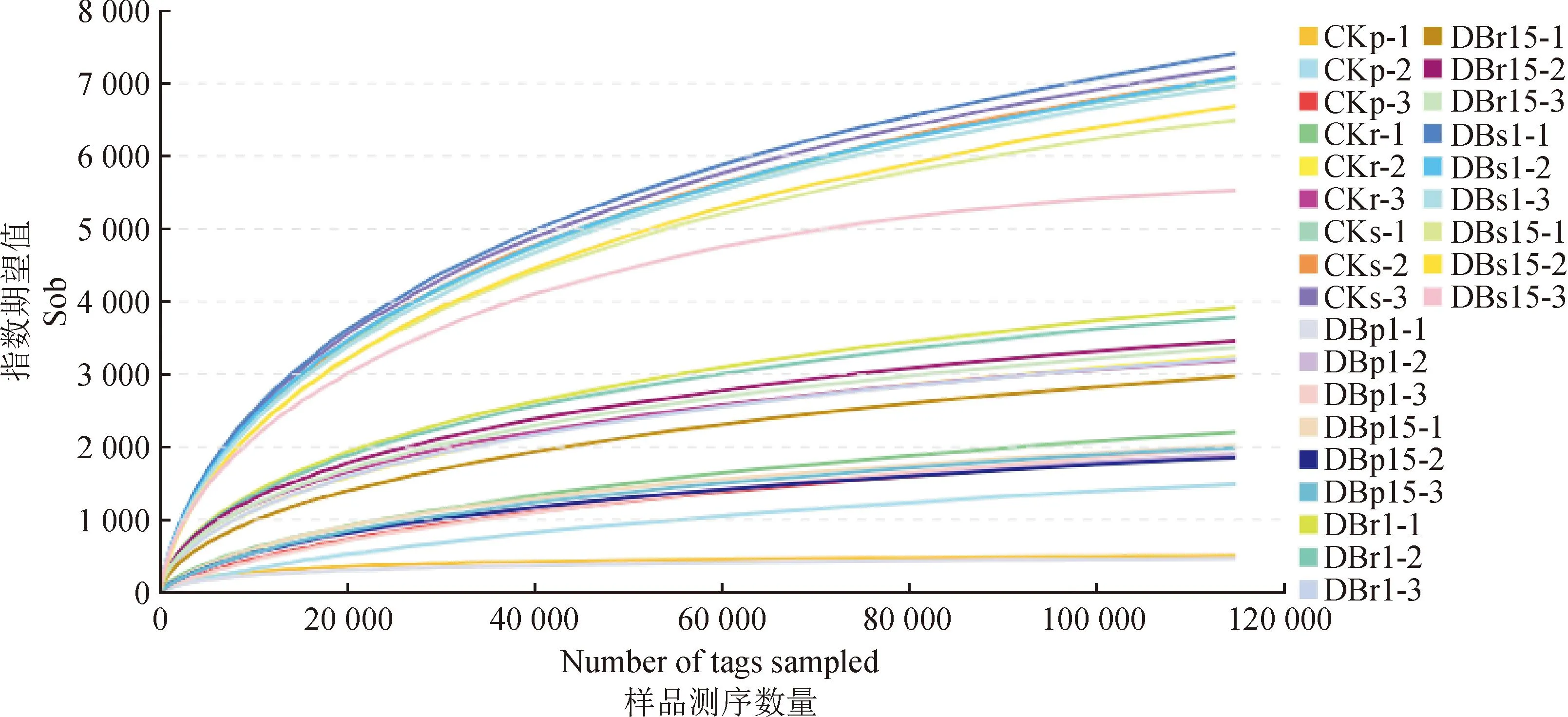
图2 转基因107杨与对照杨微生物群落多样性的稀释性曲线
2.2 物种注释及相对丰度
根据不同层级分类水平上不同样品的物种组成情况,用堆叠图的形式,直观展示不同样品在各个分类层级水平上的物种丰度的变化情况。无法注释到该水平的tags则被归类到Unclassified类别。由图3可知,各样品的优势细菌群落相同,主要分布于以下10个门:变形菌门(Proteobacteria)、蓝藻菌门(Cyanobacteria)、酸杆菌门(Acidobacteria)、拟杆菌门(Bacteroidota)、放线菌门(Actinobacteriota)、泉古菌门(Crenarchaeota)、绿弯菌门(Chloroflexi)、浮霉菌门(Planctomycetacia)、髌骨细菌门(Patescibacteria)和疣微菌门(Verrucomicrobiota)。10个门类细菌在所有细菌中所占百分比达97%,其中变形菌门为根部样品中的优势菌门,占比40.7%~58.8%。酸杆菌门和变形菌门为根际土壤样品中的优势菌门,占比16.6%~21.9%。蓝藻菌门和变形菌门为韧皮部样品中的优势菌门,占54.8%~59.2%。整体上看,转基因株系DB1、DB15菌类组成与对照株系CK极其相似,但各类细菌的数量存在差异。对照的根部变形菌门细菌数量高于转基因株系,对照和转基因株系蓝藻细菌数量无明显差异;转基因株系根部放线菌门细菌数量高于对照株系;转基因株系根际土各类细菌数量与对照基本无差异。

图3 微生物物种组成及分布(门水平)
微生物数目或种类发生变化,通常会导致样本性质发生改变。为了观测不同环境中物种的变化情况,可以对物种在不同环境下的丰度进行聚类分析。物种丰度作为观测微生物指标的重要变量,可以通过绘制微生物物种丰度聚类热图来观察不同样本的分布情况。对CK株系和转基因DB1和DB15株系的不同样品物种丰度的聚类分析(图4)可以看出,27个样品中物种丰度较高的细菌有24个门,不同株系的根际土样本形成一个分支,韧皮内微生物和根内微生物形成一个分支,但是根内微生物又独立形成一个分支,表明大部分根内微生物、韧皮内微生物和根际土微生物分别聚类到一起。其中,只有CKp-1没有与其它韧皮内微生物聚类到一起,而与根内微生物聚在一起,说明除CKp-1外,其余不同株系的相同部位微生物物种丰度差异不显著。梭杆菌门(Fusobacteria)、厚壁菌门(Firmicutes)、蓝藻门(Cyanobacteria)、栖热菌门(deinococcus-thermus)、拟杆菌门(Bacteriodetes)在韧皮部丰度最高,而放线菌门(Actinobacteria)在根内丰度最高,其余细菌在根际土内丰度最高。聚类结果表明转基因107杨对微生物物种丰度没有影响。

图4 样品门水平的微生物物种相对丰度
2.3 扩增子—α多样性分析
α多样性是指微生物在特定生物环境或者生态系统内物种数目的多样性情况,主要利用不同生境内微生物种类与分布两个参数来计算。其中超1指数(Chao1)、基于丰度的覆盖估计指数(Abundance-based Coverage Estimator,简称ACE)、香农指数(Shannon)、辛普森指数(Simpson)是常用的α多样性指数。Chao1、ACE 指数主要反应样本的物种丰富度信息,数值越大,多样性越高;Simpson、Shannon综合体现物种的丰富度和均匀度,数值越大,多样性越高。如表3所示,转基因株系DB1和DB15在韧皮内、根内和根际土内,微生物在香农指数上与对照株系不存在差异;转基因株系DB1和DB15在韧皮内和根际土内,微生物在辛普森指数上与对照株系不存在差异,只有在根内存在差异。对照株系韧皮部和根部微生物的香农指数和辛普森指数低于转基因株系,说明对照株系韧皮部内和根内微生物丰富度和均匀度较低,其他指数无显著差异。总体而言,韧皮内生菌群落中物种的数量最低,而根际土和根内样品群落中的物种数量较高,根际土样品中细菌群落表现出最高的物种多样性。
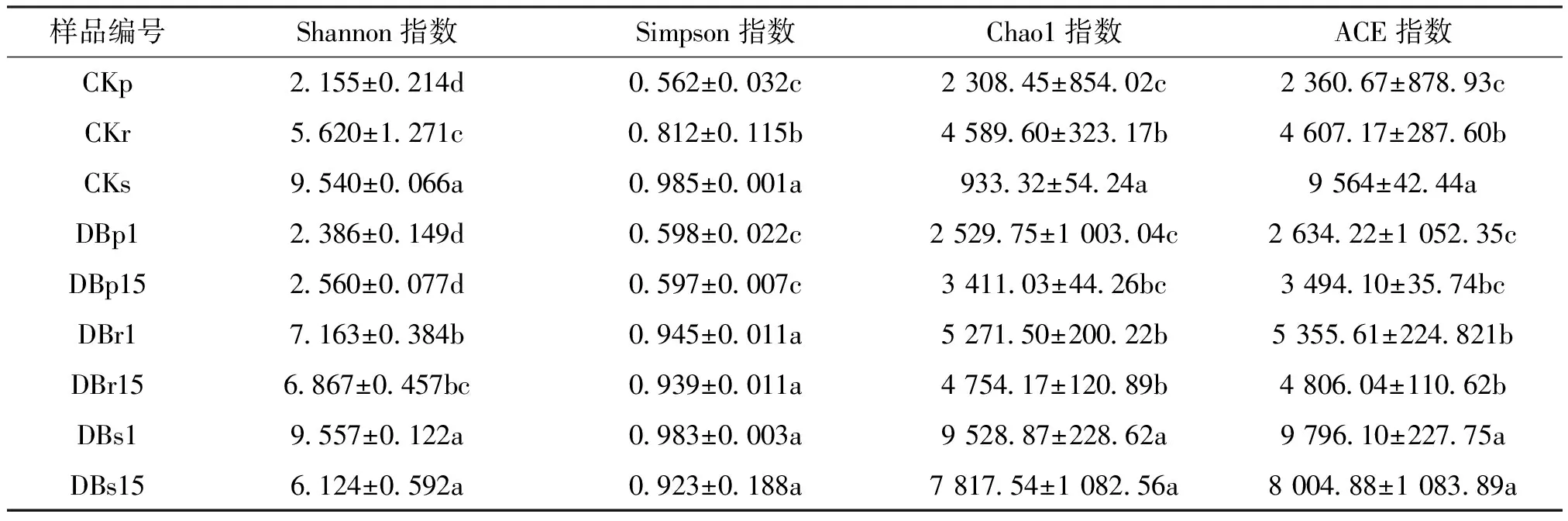
表3 各样品α多样性指数
2.4 等级丰度曲线分析
将样品中的OTUs按相对丰度(或者包含的序列数目)由大到小排序得到对应的排序编号,再以OTUs的排序编号为横坐标,OTUs中的相对丰度(也可用该等级OTU中序列数的相对百分含量)为纵坐标,将这些点用折线连接,即绘制得到等级丰度曲线(Rank Abundance)。该曲线可直观地反映样品中包含的分类丰富度和均匀度,即在水平方向,分类的丰度由曲线的宽度来反映,分类的丰富度越高,曲线在横轴上的跨度越大;在垂直方向曲线的平滑程度,反映了样品中分类的均匀程度,曲线越平缓,物种分布越均匀。由图5可以看出,韧皮内微生物在水平方向主要分布1 200~2 800之间,根内微生物在水平方向分布在2 800~4 800之间,根际土微生物主要分布在6 000~9 600,根际土微生物在横轴上的跨度最大,丰富度最高且在垂直方向上最平缓,说明物种分布均匀程度最高。对照组韧皮内微生物在垂直方向最陡,物种分布均匀度最低。说明在3个不同采样部位之间,微生物群落的丰富度和均匀度存在一定的差异和规律,但是在转基因株系和对照株系中没有明显的差异。
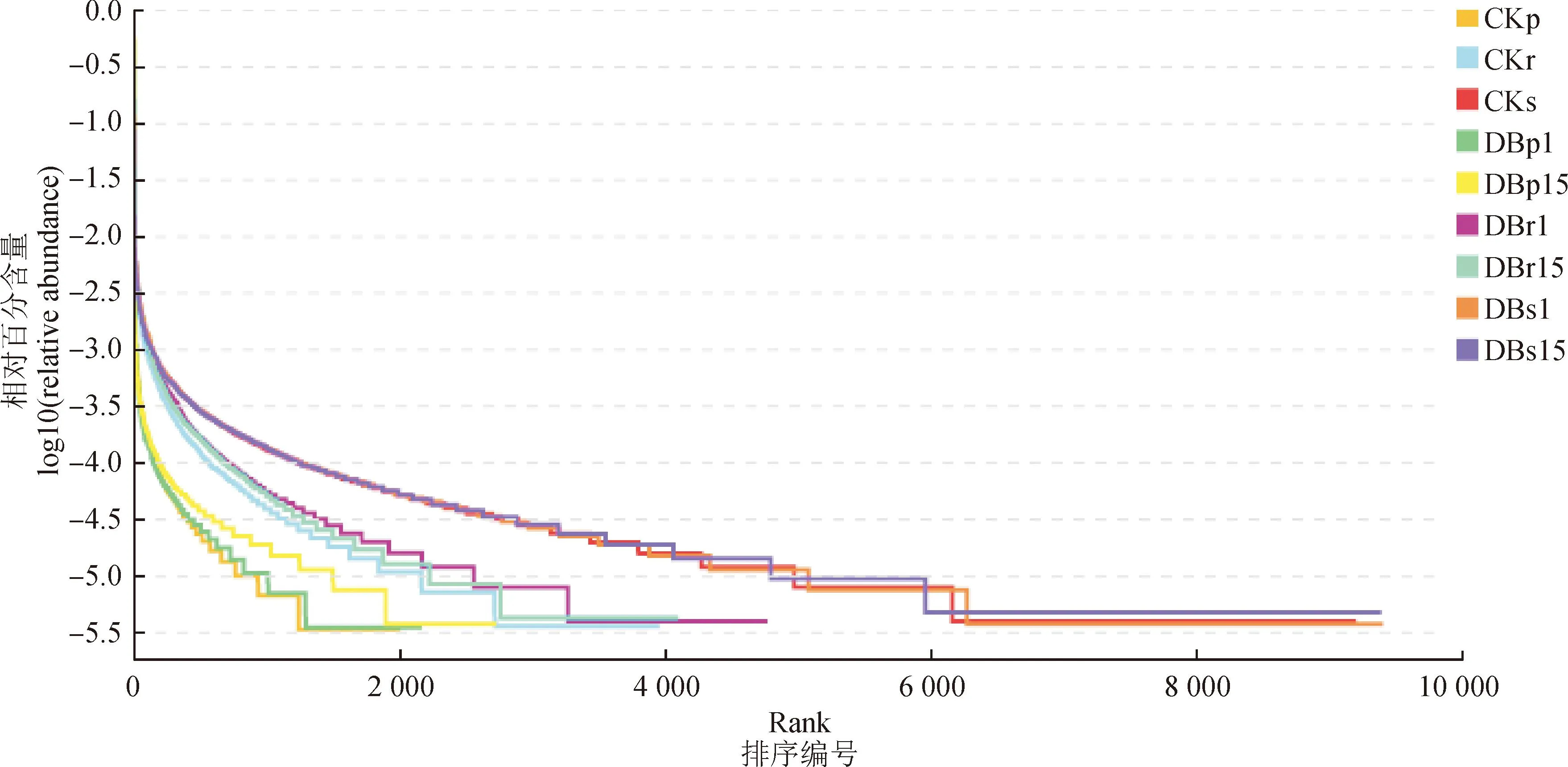
图5 样品微生物群落多样性等级丰度曲线
2.5 生境间的多样性分析
通过主坐标分析(PCoA,Principal Co-ordinates Analysis)来研究不同样品间微生物群落的相似性。该方法通过一种不受约束的降维方法找到主坐标与样本间的距离,从而判断群落相似性,结果如图6所示,主坐标PCo1坐标轴的解释度为53.3%,PCo2坐标轴的解释度为26.43%,两个主坐标轴的总解释度达到79.73%,说明PCoA聚类图可以很好地反映出细菌多样性差异。韧皮部、根际土和根内样本间并未出现聚类现象,说明样品具有良好的重复性。从PCo1维度上看,韧皮部可以与根、根际土更好地分离出来,说明根内和根际土壤微生物群落组成更相似,与韧皮部微生物群落组成差异较大。

图6 微生物分类群落PCoA分析
2.6 非加权配对平均法的聚类分析
非加权配对平均法分析(UPGMA,Unweighted Pair-group Method with Arithmetic Mean)是一种常用的聚类分析方法。基于Beta多样性分析得到的4种距离矩阵,通过R语言的Pheatmap包对样品进行UPGMA分类树分类,以判断各样品间物种组成的相似性。图7是基于bray计算方法得到的样品层次聚类树,其中越相似的样本将拥有越短的共同分支。由图7可知,根内、韧皮部微生物样品优先形成一个分支,根际土微生物样品单独聚类在一起,根内、韧皮部微生物再分别聚类到一起,即相同采样部位间微生物物种组成差异不显著。韧皮部微生物分支最短,物种相似度最高,其次为根际土微生物,根部物种组成分支最长,相似性最低,说明韧皮部和根际土物种组成相似度要略高于根部内微生物物种组成。
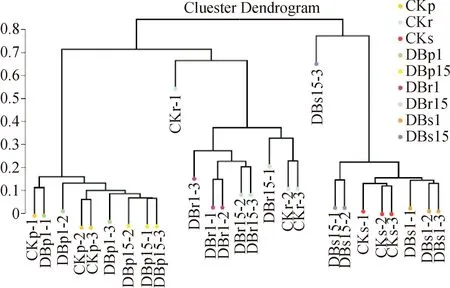
图7 样品微生物UPGMA聚类分析
2.7 指示物种分析
指示物种(biomarker)是指对分组等特征有指示作用的物种。指示物种的分析方法很多,比如韦恩图,统计检验,LEfSe等。此外,还有不依赖检验的随机森林模型、Indicator value计算等方法[24-25]。
2.7.1 韦恩图分析
利用韦恩(Venn)图可以展示样品之间共有、特有OTU数目,直观地表现出样品间OTU的重合情况。结合OTU所代表的物种,可以找出不同环境中的核心微生物。对3种不同部位样品的微生物进行韦恩图分析,根据不同组分间OTU与物种丰度比值的分析,明确样品间共同存在或者特有的微生物数量及种类,结果如图8所示。
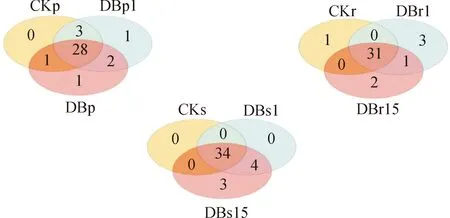
图8 样本细菌OTU物种韦恩图(门水平)
3个株系间韧皮部内微生物共有的物种数为28个,DB1独有物种1个,DB15独有物种1个;根内共有的物种数为31个,对照组独有1个,DB15独有2个,DB1独有3个;根际土共有34个物种,只有DB15独有3个物种,为螺旋毛虫(Spirochaetota)。转基因107杨DB1、DB15株系和对照杨共有微生物物种数均占总数的70%以上,3个部位独有的物种数量所占比例的总数均没超过20%,表明3个部位样品中的微生物种类差异较小。上述分析可以看出转基因杨树和非转基因杨树样品中的微生物种类差异较小。
2.7.2 富集三元图分析
当对照组有3个分组时,基于Kruskal-Wallis检验的差异分析结果,以三元图(R语言ggtern包)展示各分组显著富集(P<0.05)的物种数量,物种的相对丰度等信息,三元图用于展示3个组间物种差异检验结果,通过不同颜色表示物种所富集到的分组,直观展示每个分组中组间显著差异的物种。如图9所示,通过三元相关图(目水平)分析得知,3个株系的韧皮内微生物显著富集的物种只有1个目,放线菌目(Actinomycetes),集中富集在转基因株系DB15中;3个株系的根内微生物显著富集的物种有4个类群,均集中富集在转基因株系中,蓝藻门(Cyanophyta)、酸杆菌目(Acidobacteriales)和多囊菌目(Polyangiales)显著富集在DB1株系中,黄单胞菌目(Xanthomonadales)显著富集在DB15株系中;3个株系的根际土微生物显著富集的物种共10个目,只有巴氏杆菌科(Pasteurellaceae)和热土厌氧棒菌(Anaerobaculumthermoterrenum)显著富集在对照株系中。总体而言,相同株系不同部位间细菌群落组成和丰度在根和韧皮内差异较小,而与韧皮部群落组成差异较大,其中根际土微生物最丰富。
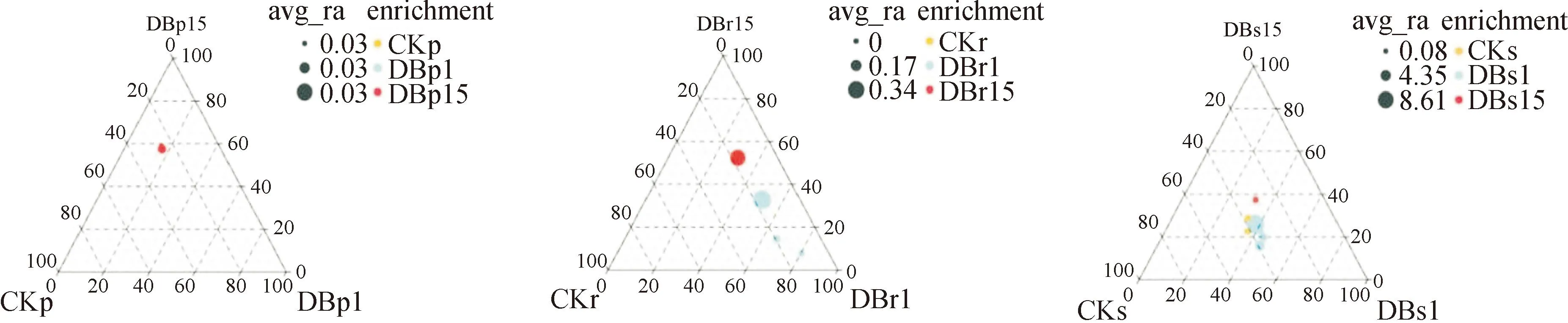
图9 微生物群落富集三元图
3 讨论与结论
3.1 讨论
许多研究表明,变形菌门一般为各土壤样品中的优势菌门[26-27],在土壤中变形菌门、浮霉菌门、拟杆菌、厚壁菌门、放线菌、酸杆菌门、蓝藻菌门、绿弯菌门等是较为常见的菌门,一般在林地土壤中可占总比例的90%以上,这与本研究的结果一致。转基因株系与非转基因株系在菌类物种组成门水平下没有明显差异,而相同株系不同部位样品之间的物种组成存在一定的差异。影响植物体内微生物种类和数量变化的因素有多种,首先是植物生长环境的稳定性和复杂性,有研究认为根际生境由于可提供大量的内在碳源,如糖类、氨基酸、有机酸等,因而成为更加复杂的微生物栖息地[28],受到外界环境影响也就更大了。有研究揭示植物基因型是根际菌群落形成的一个重要决定性因素[29-30],大量的研究证明土壤类型、种植年限和田地的异质性等因素,要比外源基因插入植物会更多地干扰根相关微生物群落的组成[31-32],其次是植物自身的生长需要,内生菌普遍存在于高等植物中,植物内生菌可以刺激植物生长或提高宿主植物的自然抗性[33-34]。本研究中转基因株系与对照间微生物存在少量差异物种,通过统计分析,仅有个别菌种显著富集,且富集物种丰度极低。不同种类的植物,甚至是生长在相同环境的同种类植物都会具有独特的内生菌类型[35-37],多种因素共同影响着植物体内微生物群落的组成。
本研究中转抗虫基因107杨的根系与韧皮部内生细菌Shannon指数和Simpson指数差异相对较小,在物种均匀度方面也没有表现出明显差异。Shannon指数分析说明了转基因株系和非转基因株系差异不大,不同部位之间差异较显著;Simpson指数在根际土壤、根部和韧皮部之间都没有显著差异,物种分布均匀,多样性高;转基因株系和非转基因株系之间也没有显著差异。
3.2 结论
对转基因107杨DB1、DB15株系和对照株系的不同部位(韧皮部、根、根际土)的微生物群落进行高通量测序分析可知,在根际土壤、根部和韧皮部转基因株系和对照之间的细菌群落组成没有表现出明显差异,不同部位之间差异显著。韧皮部和根部的细菌群落组成相似度极高,二者与根际土壤的细菌群落组成差异较大。根际土壤和韧皮部细菌的丰富度高于根部。多样性指数分析发现转抗虫基因107杨内生细菌种类丰富,没有对韧皮、根、根际土中的微生物群落多样性产生影响。综上所述说明转基因杨树未对自身和土壤中的细菌群落产生显著的影响。研究结果为转基因杨树的生态安全性评价提供了参考,同时也为转基因杨树环境释放后的自然生态系统风险评估提供了更加系统的认识,对其生物安全性全面、系统的研究有待进一步深入。
