RNA m6A甲基化在卒中后认知障碍中的研究进展
2024-04-01肖雨倩孙可心万俊陈淑颖陈丽敏王岩白艳杰
肖雨倩 孙可心 万俊 陈淑颖 陈丽敏 王岩 白艳杰
摘要:卒中后认知障碍(PSCI)主要表现为学习、记忆等方面的障碍。哺乳动物大脑中高度富集的RNA m6A甲基化修饰,参与神经胶质细胞介导的神经炎症。鉴于神经炎症是PSCI神经损伤以及空间和记忆能力下降的主要机制,推测RNA m6A甲基化修饰可调节脑卒中后神经胶质细胞炎症反应,进而改善PSCI。该文就RNA m6A甲基化修饰在PSCI发展中的作用及其调控神经胶质细胞介导的炎症的详细机制进行总结分析,为该领域的研究者提供参考。
关键词:卒中;认知障碍;炎症;小神经胶质细胞;星形细胞;m6A甲基化
中图分类号:R743.3文献标志码:ADOI:10.11958/20230780
Research progress of RNA m6A methylation in post-stroke cognitive impairment
XIAO Yuqian1, SUN Kexin1, WAN Jun1, CHEN Shuying1, CHEN Limin1, WANG Yan1, BAI Yanjie2△
1 Henan University of Chinese Medicine, Zhengzhou 450000, China; 2 the First Affiliated Hospital of Henan University of Chinese Medicine
△Correspongding Author E-mail: baiyj66@126.com
Abstract: Post-stroke cognitive impairment (PSCI) is mainly manifested as learning and memory disorders. Highly enriched RNA m6A methylation modification in mammalian brain is involved in glial cell-mediated neuroinflammation. Given that neuroinflammation is the main mechanism for neural damage and spatial and memory impairment of PSCI, it is speculated that RNA m6A methylation modification can regulate the inflammatory response of glial cells after stroke to improve PSCI. This review summarizes and analyzes the role of RNA m6A methylation modification in the development of PSCI and analyzes its detailed mechanism of regulating glial cell-mediated inflammation, which will provide reference for researchers in this field.
Key words: stroke; cognition disorders; inflammation; microglia; astrocytes; m6A methylation
腦卒中是全球第二大常见死因,通常会导致运动、认知、语言和心理障碍。卒中后认知障碍(post stroke cognitive impairment,PSCI)主要表现为学习、记忆力、执行力等方面功能下降,包括运动和言语功能障碍等一系列症状。缺血缺氧引起的氧化应激、炎症反应、细胞凋亡、突触损伤、血管破坏等是PSCI的主要诱因[1]。神经胶质细胞在缺血性脑损伤的预后中起着至关重要的作用,其抗炎表型有助于减轻炎症反应。有研究报道,许多药物通过调节缺血或灌注不足模型中的小胶质细胞极化和炎症反应来减轻脑损伤[2]。RNA修饰是神经胶质细胞响应细胞外刺激而激活的一个关键细胞内机制,N6-甲基腺苷(N6-methyladenosine,m6A)甲基化修饰是研究最广泛的一种[3]。m6A于1974年首次在病毒RNA的内部修饰中被报道,是指在腺嘌呤碱基的第6号氮处选择性地添加甲基,通过调节RNA剪接、定位和翻译,影响各种生物进程[4]。最近研究发现,m6A修饰可以调节神经胶质细胞的促炎和抗炎反应[5]。鉴于神经胶质细胞在神经炎症过程中的关键调节作用,推测m6A修饰可以调节PSCI中神经胶质细胞炎症反应。本文旨在阐明RNA m6A甲基化修饰调控PSCI的详细机制及此修饰调控神经胶质细胞炎症反应参与PSCI的研究现状和未来方向。
1 RNA m6A甲基化修饰及其调控蛋白
1.1 RNA m6A甲基转移酶 RNA m6A甲基化修饰相关的酶和蛋白质可分为3类:m6A甲基转移酶、m6A去甲基转移酶和m6A结合蛋白。甲基转移酶和去甲基转移酶以高度动态的方式平衡m6A甲基化修饰水平。甲基转移酶负责在靶RNA上安装甲基,典型的甲基添加由多功能亚基组成的编码复合物催化。甲基转移酶样(METTL)3和METTL14形成异二聚体作为编码复合物核心,前者具有催化能力,而METTL14变构可激活METTL3并促进RNA结合[6]。最近的一项研究提出,METTL16也是一种独立的m6A甲基转移酶,并且在RNA剪接调节中起着关键作用[7]。研究还发现了一些与m6A甲基化相关的其他成分,例如Wilms肿瘤蛋白1相关蛋白(WTAP)、肾母细胞瘤1相关蛋白(VIRMA)、RNA结合蛋白15(RBM15)及其旁系蛋白RBM15B和一种含有CCCH序列的锌指蛋白13(ZC3H13)等[8]。
1.2 RNA m6A去甲基转移酶 目前,去甲基转移酶仅占m6A调节器的一小部分,也被称为“橡皮擦”。两种公认的去甲基转移酶是肥胖相关蛋白(fat mass and obesity associated,FTO)和ALKB同系物5(ALKB homologue 5,ALKBH5)。FTO在神经元中广泛表达和高度富集,其通过改变选择性剪接和翻译的模式来调节基因表达。FTO的缺失会导致海马体内神经元分化受损和脑源性神经营养因子(brain-derived neurotrophic factor,BDNF)的表达减少,从而增加焦虑并损害工作记忆[9]。ALKBH5是一种定位在核斑点的核蛋白,其介导的去甲基化活性影响核RNA输出和RNA代谢,从而调节基因表达。ALKBH5主要在神经元中表达,并在神经发育过程中动态增加,导致神经干细胞的增殖和分化[10]。ALKBH5、FTO和其他未定义的去甲基化酶很可能在去甲基化活性上重叠,在正常情况下可以部分替代彼此的功能。
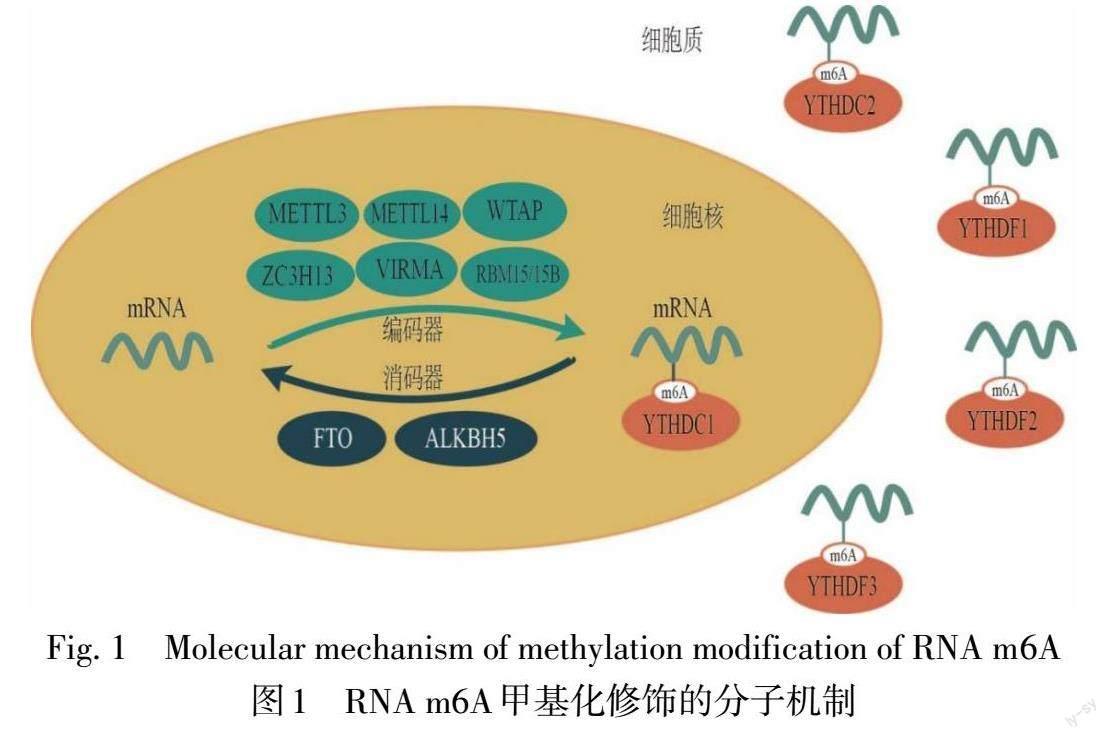
1.3 RNA m6A结合蛋白 RNA m6A结合蛋白也称为“阅读器”,在上述两种酶修饰后,通过识别和翻译不同转录物上的m6A位点来影响靶mRNA的剪接、核转运、稳定性、翻译和RNA衰变。根据其与m6A特异性结合的能力分为直接和间接结合蛋白。直接结合蛋白包括5种含YTH结构域的蛋白质:YTHDF1/2/3和YTHDC1/2。YTHDF1主要影响修饰基因的翻译,YTHDF2主要负责降解,YTHDF3可与前两者合作,加速靶转录物的翻译或降解[11]。间接结合蛋白主要是异质核糖核蛋白(heterogeneous nuclear ribonucleoprotein,HNRNPs),包括HNRNPC、HNRNPG和HNRNPA2/B1。此外,胰岛素样生长因子2 mRNA结合蛋白(insulin-like growth factor 2 mRNA-binding proteins,IGF2BPs)通过识别共有GGC序列靶向数千个mRNA转录物,在正常和应激条件下以m6A依赖的方式参与转录后RNA修饰[12]。在胞质中METTL3也可作为阅读器,通过与真核翻译起始因子3(eukaryotic initiation factor 3,eIF3)相互作用增强mRNA翻译[13]。RNA m6A甲基化修饰分子机制如图1所示。
2 RNA m6A甲基化修饰在PSCI中的作用
2.1 调控细胞凋亡 m6A修饰在人类、小鼠和大鼠的大脑皮质中广泛存在,与其他组织相比,m6A修饰对脑的组织特异性最强。磷酸酯酶与张力蛋白同源物(phosphatase and tension homologous protein,PTEN)是一种常见的肿瘤抑制因子,敲低YTHDF1可通过降低PTEN稳定性阻止PTEN/蛋白激酶B(Akt)/雷帕霉素靶蛋白(mTOR)信号传导,抑制细胞凋亡,减轻脑缺血/再灌注(ischemia/reperfusion,I/R)损伤诱导的脑梗死和神经功能障碍,而YTHDF1过表达的结果相反[14]。
应激颗粒是由RNA结合蛋白和mRNA组成的无膜细胞器,可以立即且短暂地阻止mRNA翻译,以保护有价值的mRNA和蛋白质免受有害环境的伤害,从而提高早期急性缺血性脑卒中患者脑细胞的存活率。Si等[15]证明在糖氧剥夺/复氧(oxygen-glucose deprivation/reoxygenation,OGD/R)处理后,METTL3介导的m6A甲基化含量在原代培养的大鼠海马神经元中明显增多,通过增加产生成熟的miR-335促进卒中早期的应激颗粒生成,缓解神经元损伤和细胞的凋亡。B淋巴细胞瘤因子2(B-cell lymphoma-2,Bcl-2)作为抗凋亡蛋白在调节细胞凋亡中起重要作用,上调Bcl-2的蛋白和mRNA表达可以减少双侧颈总动脉闭塞诱导的大鼠海马细胞凋亡,改善其认知障碍[16]。敲低ALKBH5显著降低了原代神经元中Bcl-2的表达,导致神经元凋亡增强;相反,FTO表达增多时刺激Bcl-2水平上升,对神经元起到保护作用,m6A去甲基化可能通过升高Bcl-2缓解PSCI[17]。在体外,OGD/R诱导的SH-SY5Y细胞中,添加FTO后SH-SY5Y细胞ROS产生和超氧化物歧化酶活性降低;在体内,对大脑中动脉闭塞(MCAO)大鼠静脉注射FTO,FTO过表达通过介导核因子-E2相关因子2(nuclearfactor erythroidderived 2-like 2,Nrf2)mRNA的去甲基化,可降低YTHDF2依赖性mRNA降解并增加Nrf2表达,从而抑制氧化应激反应和减少细胞凋亡,最终缓解脑I/R损伤[18]。进一步研究发现,骨髓间充质干细胞来源的外泌体Krüppel样因子4通过靶向长链非编码RNA(lncRNA)-ZFAS1增加FTO水平,从而降低动力相关蛋白1的m6A修饰,对I/R诱导的线粒体损伤和细胞凋亡产生抑制作用[19]。
2.2 参与神经元和轴突的生长发育 PSCI是一个不可逆的过程,可直接对大脑感觉、运动和自主神经功能产生破坏,损害突触形态及生存环境。m6A甲基化调节神经发育过程中参与轴突引导和伸长的mRNA的翻译。FTO mRNA和蛋白水平在包括海马体在内的多个大脑区域中都非常丰富,敲除FTO会显著降低BDNF在小鼠海马体中的表达,降低学习和记忆表现[20]。维持轴突的正确导向是大脑中神经回路形成的关键一环,YTHDF1可促进小鼠背侧脊髓中轴突导向相关蛋白Robo3.1 mRNA的甲基化,增加Robo3.1蛋白水平,从而在控制轴突导向中起重要作用[21]。Shi等[22]研究发现,在海马神经元中,YTHDF1以神经元刺激依赖的方式促进目标转录物的翻译,增强蛋白质合成,从而改善小鼠的学习和记忆能力,YTHDF1的耗竭会损害海马突触的基础传递和长时程增强(LTP)。METTL3耗竭将抑制神经元增殖和分化,通过组蛋白甲基轉移酶Ezh2干扰神经干细胞向神经胶质谱系分化,抑制METTL3介导的m6A甲基化可以保护神经元免受I/R损伤[23]。miR-422a是大脑中富含的miRNA家族成员之一,在急性缺血性卒中患者脑组织中显著上调,介导神经元细胞死亡和凋亡。OGD/R刺激下,METTL3介导的Lnc-D63785 m6A甲基化是诱导miR-422a积累和神经元细胞凋亡的关键,对METTL3的抑制可以逆转Lnc-D63785造成的损伤[24]。
2.3 参与血管修复与生成 脑卒中后可迅速触发参与血管生成过程中重要血管生成因子的诱导和激活,促进脑血管生成是改善PSCI的有潜力的治疗策略[25]。m6A甲基化在包括血管生成在内的多种发育决策中起着重要作用[26]。研究表明,缺氧应激后内皮细胞中的m6A甲基化水平显著上调,并对血管修复和血管生成产生积极影响[27]。circSCMH1通过促进FTO泛素化增加了FTO的核易位,导致磷脂磷酸酶3 mRNA的m6A去甲基化,从而诱导内皮细胞中脂质磷酸磷酸酶3水平增加,随后增强血管修复,加快了小鼠卒中后功能恢复[28]。血脑屏障主要由脑微血管内皮细胞和紧密连接构成,是维持中枢神经系统稳态的重要物理屏障,通过对m6A修饰的抑制可减少基质金属蛋白酶(matrix metallopeptidase,MMP)3的表达,进而改善小鼠脑I/R损伤后血管内皮细胞的修复[29]。此外,脑内皮细胞中发现了一种新型m6A结合蛋白PRRC2B(proline rich coiled-coil 2B),可调节MMP14和解整合素金属蛋白酶19,促进缺氧诱导的内皮细胞迁移,PRRC2B敲除会加剧脑血管重塑,进而重新分配脑血流,改善缺氧诱导的小鼠认知能力[30]。
2.4 调控神经炎症 抑制炎症反应可以改善PSCI大鼠的认知功能[31]。异常的m6A修饰与神经炎症密切相关,脑卒中患者外周血的m6A甲基化水平均升高[32]。RNA甲基化免疫共沉淀表明,卒中后小鼠炎症相关基因m6A甲基化增加[33]。神经胶质细胞在缺血性脑卒中后的神经炎症起双向调节作用,研究发现,m6A甲基化修饰可以调控神经胶质细胞介导的炎症反应[3]。
2.4.1 调控小胶质细胞介导的神经炎症 小胶质细胞是主要的脑驻留细胞,几乎参与所有中枢神经系统病理过程。当被Toll样受体(Toll-like receptor,TLR)和其他刺激诱导时,稳态小胶质细胞(M0)被激活并极化为M1促炎表型和M2抗炎表型。促炎小胶质细胞会引发脑损伤,阻碍神经发生,干扰卒中后神经功能的恢复和修复。目前公认LTP是学习和记忆的基础,海马CA1中的LTP可被小胶质细胞释放的白细胞介素-1β(IL-1β)侵袭所损伤[34]。干扰素调节因子(interferon regulatory factor,IRF)5/IRF4调节轴是小胶质细胞促炎和抗炎激活的关键,小鼠IRF5敲除可导致M2表型激活增强并改善PSCI[35]。
m6A甲基化修饰与调节小胶质细胞炎症反应的基因表达具有动态和复杂的关系。有研究揭示了M0、M1和M2型小胶质细胞中mRNA和lncRNA中的m6A甲基化谱,发现87个lncRNA在M1型和M2型小胶质细胞中甲基化修饰有所差异,差异甲基化修饰的lncRNA通过改变多种信号转导途径来调节小胶质细胞介导的炎症反应[3]。Wen等[36]发现小胶质细胞中METTL3的水平与TNF受体相关因子6(TNF receptor associated factor 6,TRAF6)呈正相关,TRAF6/NF-κB通路可能在METTL3过表达条件下以m6A甲基化依赖性方式被激活,最终导致小胶质细胞炎症。m6A结合蛋白IGF2BP1通过增强Gbp11和Cp mRNAs的稳定性增强了小胶质细胞的炎症反应[37]。沉默信息调节因子(silence information regulator,SIRT)1是一种NAD依赖性脱乙酰酶,YTHDC1的沉默下调了SIRT1的表达,抑制M1小胶质细胞向M2表型的转化,诱导M1小胶质细胞活化并加剧炎症反应[38]。
RNA m6A甲基化修饰相关的酶和蛋白可能通过增强小胶质细胞炎症反应参与PSCI。在MCAO小鼠和OGD诱导的小胶质细胞模型中,由于miR-421-3p表达的下调,导致YTHDF1表达增加,YTHDF1随后识别m6A甲基化修饰的p65 mRNA并促进其翻译和核转运,最终激活NF-κB信号通路并促进炎症反应[39]。此外,急性缺血性脑卒中时血清脂多糖(LPS)升高,LPS暴露会导致大脑中炎症介质急剧增加,认知能力下降[40-41]。研究发现,LPS刺激显著改变小胶质细胞的m6A修饰,TLR4被LPS激活,随后METTL3介导的m6A修饰上调了TLR4 mRNA的表达,通过TLR4/TLR相关的干扰素活化子(TRIF)-TLR相关分子(TRAM)/NF-κB途径介导M1小胶质细胞活化[42]。在老年小鼠模型中,海马体中的METTL3敲除可诱导神经变性和小胶质细胞数量的减少,可能是m6A修饰通过程序性死亡受体1/程序性死亡受體-配体1途径影响空间认知功能的基础[43]。环鸟苷酸腺苷酸合酶(cyclic guanosine monophosphate-adenosine monophosphate synthetase,cGAS)-干扰素基因的刺激因子(stimulator of interferon genes,STING)信号通路在促进小胶质细胞M1极化以加重脑缺血性脑卒中神经炎症方面起重要作用,FTO通过m6A修饰降低cGAS mRNA稳定性,抑制cGAS表达,缓解小胶质细胞介导的炎症反应,从而减轻脑I/R损伤中的神经炎症[5]。此外,FTO过表达减少总体RNA m6A修饰,并通过下调pri-miR-155的m6A修饰来抑制其成熟过程,从而减少神经炎症对脑I/R的损伤,发挥保护作用[44]。
2.4.2 调控星形胶质细胞介导的神经炎症 星形胶质细胞是中枢神经系统中数量最多和最特殊的分支细胞类型,主要负责维持大脑稳态,通过释放多种介质来影响中枢神经系统中各种细胞的功能。在不同条件下,星形胶质细胞可以被激活并分化为不同的亚型,包括A1神经毒性表型和A2神经保护表型。反应性星形胶质细胞中Na+/H+交换蛋白1型(Na+/H+ exchanger isoform 1,NHE1)的活化可导致星形胶质细胞肥大和肿胀,抑制NHE1表达可减少ROS产生和减轻炎症反应,保留白质和海马完整性,改善慢性脑灌注不足引起的认知功能障碍[45]。
在鏈脲佐菌素诱导的阿尔茨海默病(AD)模型中,当MO-I-500作为FTO抑制剂下调星形胶质细胞中FTO表达时,大脑中氧化应激和细胞凋亡明显降低,线粒体功能障碍和能量代谢紊乱明显改善,神经炎症反应减弱[46]。另一种去甲基化酶ALKBH5对星形胶质细胞具有相似效果。circSTAG1可以与小鼠海马中的ALKBH5结合,降低ALKBH5水平以改变星形胶质细胞中脂肪酸酰胺水解酶(fatty acid amide hydrolase,FAAH)的m6A甲基化水平,加速FAAH降解,最终导致星形胶质细胞功能障碍[47]。进一步研究发现,星形胶质细胞来源的IL-1β囊泡促进了m6A阅读器HNRNPC与淀粉样蛋白前体蛋白(APP)mRNA的结合,以增强APP翻译和淀粉样蛋白β(Aβ)的产生,从而加剧神经慢性炎症[48]。而Aβ沉积是认知能力下降的重要原因之一,可能与PSCI密切相关[49]。
3 小结和展望
脑卒中会改变大脑表观转录组,调节m6A甲基化修饰进而调控炎症、细胞凋亡、血管修复与生成以及神经元与轴突的生长发育过程,有望成为改善PSCI的有效策略。同时,动态和可逆的RNA m6A甲基化可以调控神经胶质细胞的活化和极化,也可在脑卒中后神经胶质细胞诱导的炎症反应中发挥作用,对神经胶质细胞甲基化的控制可能减少炎症引起的继发性脑损伤,进而改善认知障碍。
研究m6A甲基化与PSCI间的相互作用有助于更好地了解PSCI的发病机制,并发现PSCI治疗的新靶点。然而,很少有学者在神经胶质细胞的背景下探索m6A甲基化修饰与PSCI的关系,故仍需深入研究m6A甲基化修饰在PSCI发生发展过程中如何参与神经胶质细胞的精确调控。
参考文献
[1] LEE K P,CHANG A Y W,SUNG P S. Association between blood pressure,blood pressure variability,and post-stroke cognitive impairment[J]. Biomedicines,2021,9(7):773. doi:10.3390/biomedicines9070773.
[2] RAN Y,SU W,GAO F,et al. Curcumin ameliorates white matter injury after ischemic stroke by inhibiting microglia/macrophage pyroptosis through NF-κB suppression and NLRP3 inflammasome inhibition[J]. Oxid Med Cell Longev,2021,2021:1552127. doi:10.1155/2021/1552127.
[3] LI Q,WEN S,YE W,et al. The potential roles of m6A modification in regulating the inflammatory response in microglia[J]. J Neuroinflammation,2021,18(1):149. doi:10.1186/s12974-021-02205-z.
[4] ZACCARA S,RIES R J,JAFFREY S R. Reading,writing and erasing mRNA methylation[J]. Nat Rev Mol Cell Biol,2019,20(10):608-624. doi:10.1038/s41580-019-0168-5.
[5] YU Z,ZHENG L,GENG Y,et al. FTO alleviates cerebral ischemia/reperfusion-induced neuroinflammation by decreasing cGAS mRNA stability in an m6A-dependent manner[J]. Cell Signal,2023,109:110751. doi:10.1016/j.cellsig.2023.110751.
[6] SCH?LLER E,WEICHMANN F,TREIBER T,et al. Interactions,localization,and phosphorylation of the m6A generating METTL3-METTL14-WTAP complex[J]. RNA,2018,24(4):499-512. doi:10.1261/rna.064063.117.
[7] AOYAMA T,YAMASHITA S,TOMITA K. Mechanistic insights into m6A modification of U6 snRNA by human METTL16[J]. Nucleic Acids Res,2020,48(9):5157-5168. doi:10.1093/nar/gkaa227.
[8] ZOU J,LIU H,TAN W,et al. Dynamic regulation and key roles of ribonucleic acid methylation[J]. Front Cell Neurosci,2022,16:1058083. doi:10.3389/fncel.2022.1058083.
[9] SPYCHALA A,R?THER U. FTO affects hippocampal function by regulation of BDNF processing[J]. PloS One,2019,14(2):e0211937. doi:10.1371/journal.pone.0211937.
[10] DU T,LI G,YANG J,et al. RNA demethylase Alkbh5 is widely expressed in neurons and decreased during brain development[J]. Brain Res Bull,2020,163:150-159. doi:10.1016/j.brainresbull.
2020.07.018.
[11] SHI R,YING S,LI Y,et al. Linking the YTH domain to cancer:the importance of YTH family proteins in epigenetics[J]. Cell Death Dis,2021,12(4):346. doi:10.1038/s41419-021-03625-8.
[12] SUN C Y,CAO D,DU B B,et al. The role of insulin-like growth factor 2 mRNA-binding proteins(IGF2BPs)as m(6)A readers in cancer[J]. Int J Biol Sci,2022,18(7):2744-2758. doi:10.7150/ijbs.70458.
[13] CHOE J,LIN S,ZHANG W,et al. mRNA circularization by METTL3-eIF3h enhances translation and promotes oncogenesis[J]. Nature,2018,561(7724):556-560. doi:10.1038/s41586-018-0538-8.
[14] LI X,AN P,HAN F,et al. Silencing of YTHDF1 attenuates cerebral stroke by inducing PTEN degradation and activating the PTEN/AKT/mTOR pathway[J]. Mol Biotechnol,2022,65(5):822-832. doi:10.1007/s12033-022-00575-0.
[15] SI W,LI Y,YE S,et al. Methyltransferase 3 mediated miRNA m6A methylation promotes stress granule formation in the early stage of acute ischemic stroke[J]. Front Mol Neurosci,2020,13:103. doi:10.3389/fnmol.2020.00103.
[16] LIU Y,YAN Z,REN Y,et al. Electroacupuncture inhibits hippocampal neuronal apoptosis and improves cognitive dysfunction in mice with vascular dementia via the JNK signaling pathway[J]. Acupunct Med,2022:9645284221136878. doi:10.1177/09645284221136878.
[17] XU K,MO Y,LI D,et al. N(6)-methyladenosine demethylases Alkbh5/Fto regulate cerebral ischemia-reperfusion injury[J]. Ther Adv Chronic Dis,2020,11:2040622320916024. doi:10.1177/2040622320916024.
[18] HOU L,LI S,LI S,et al. FTO inhibits oxidative stress by mediating m6A demethylation of Nrf2 to alleviate cerebral ischemia/reperfusion injury[J]. J Physiol Biochem,2023,79(1):133-146. doi:10.1007/s13105-022-00929-x.
[19] WANG Q S,XIAO R J,PENG J,et al. Bone marrow mesenchymal stem cell-derived exosomal KLF4 alleviated ischemic stroke through inhibiting N6-Methyladenosine modification level of Drp1 by targeting lncRNA-ZFAS1[J]. Mol Neurobiol,2023,60(7):3945-3962. doi:10.1007/s12035-023-03301-2.
[20] LI L,ZANG L,ZHANG F,et al. Fat mass and obesity-associated(FTO)protein regulates adult neurogenesis[J]. Hum Mol Genet,2017,26(13):2398-2411. doi:10.1093/hmg/ddx128.
[21] ZHUANG M,LI X,ZHU J,et al. The m6A reader YTHDF1 regulates axon guidance through translational control of Robo3.1 expression[J]. Nucleic Acids Res,2019,47(9):4765-4777. doi:10.1093/nar/gkz157.
[22] SHI H,ZHANG X,WENG Y L,et al. m(6)A facilitates hippocampus-dependent learning and memory through YTHDF1[J]. Nature,2018,563(7730):249-253. doi:10.1038/s41586-018-0666-1.
[23] CHEN J,ZHANG Y C,HUANG C,et al. m(6)A regulates neurogenesis and neuronal development by modulating histone methyltransferase Ezh2[J]. Genomics Proteomics Bioinformatics,2019,17(2):154-168. doi:10.1016/j.gpb.2018.12.007.
[24] XU S,LI Y,CHEN J P,et al. Oxygen glucose deprivation/re-oxygenation-induced neuronal cell death is associated with Lnc-D63785 m6A methylation and miR-422a accumulation[J]. Cell Death Dis,2020,11(9):816. doi:10.1038/s41419-020-03021-8.
[25] SUN P,MA F,XU Y,et al. Genetic deletion of endothelial microRNA-15a/16-1 promotes cerebral angiogenesis and neurological recovery in ischemic stroke through Src signaling pathway[J]. J Cereb Blood Flow Metab,2021,41(10):2725-2742. doi:10.1177/0271678x211010351.
[26] MATHIYALAGAN P,ADAMIAK M,MAYOURIAN J,et al. FTO-dependent N(6)-methyladenosine regulates cardiac function during remodeling and repair[J]. Circulation,2019,139(4):518-532. doi:10.1161/circulationaha.118.033794.
[27] YAO M D,JIANG Q,MA Y,et al. Role of METTL3-dependent N(6)-methyladenosine mRNA modification in the promotion of angiogenesis[J]. Mol Ther,2020,28(10):2191-2202. doi:10.1016/j.ymthe.2020.07.022.
[28] LI B,XI W,BAI Y,et al. FTO-dependent m(6)A modification of Plpp3 in circSCMH1-regulated vascular repair and functional recovery following stroke[J]. Nat Commun,2023,14(1):489. doi:10.1038/s41467-023-36008-y.
[29] LIANG E,XIAO S,ZHAO C,et al. M6A modification promotes blood-brain barrier breakdown during cerebral ischemia/reperfusion injury through increasing matrix metalloproteinase 3 expression[J]. Heliyon,2023,9(6):e16905. doi:10.1016/j.heliyon.2023.e16905.
[30] LI S,HU W,GONG S,et al. The role of PRRC2B in cerebral vascular remodeling under acute hypoxia in mice[J]. Adv Sci(Weinh),2023:e2300892. doi:10.1002/advs.202300892.
[31] ZHANG X,YUAN M,YANG S,et al. Enriched environment improves post-stroke cognitive impairment and inhibits neuroinflammation and oxidative stress by activating Nrf2-ARE pathway[J]. Int J Neurosci,2021,131(7):641-649. doi:10.1080/00207454.2020.1797722.
[32] ZHU L,LIU S,LIAO F,et al. Comprehensive analysis of blood-based m6A methylation in human ischemic stroke[J]. Mol Neurobiol,2023,60(2):431-446. doi:10.1007/s12035-022-03064-2.
[33] CHOKKALLA A K,MEHTA S L,KIM T,et al. Transient focal ischemia significantly alters the m(6)A epitranscriptomic tagging of RNAs in the brain[J]. Stroke,2019,50(10):2912-2921. doi:10.1161/strokeaha.119.026433.
[34] HOSHINO K,HASEGAWA K,KAMIYA H,et al. Synapse-specific effects of IL-1β on long-term potentiation in the mouse hippocampus[J]. Biomed Res,2017,38(3):183-188. doi:10.2220/biomedres.38.183.
[35] AL MAMUN A,CHAUHAN A,QI S, et al. Microglial IRF5-IRF4 regulatory axis regulates neuroinflammation after cerebral ischemia and impacts stroke outcomes[J]. Proc Natl Acad Sci U S A,2020,117(3):1742-1752. doi:10.1073/pnas.1914742117.
[36] WEN L,SUN W,XIA D,et al. The m6A methyltransferase METTL3 promotes LPS-induced microglia inflammation through TRAF6/NF-κB pathway[J]. Neuroreport,2022,33(6):243-251. doi:10.1097/wnr.0000000000001550.
[37] DING L,WU H,WANG Y,et al. m6A reader Igf2bp1 regulates the inflammatory responses of microglia by stabilizing Gbp11 and Cp mRNAs[J]. Front Immunol,2022,13:872252. doi:10.3389/fimmu.2022.872252.
[38] ZHOU H,XU Z,LIAO X,et al. Low expression of YTH domain-containing 1 promotes microglial M1 polarization by reducing the stability of sirtuin 1 mRNA[J]. Front Cell Neurosci,2021,15:774305. doi:10.3389/fncel.2021.774305.
[39] ZHENG L,TANG X,LU M,et al. microRNA-421-3p prevents inflammatory response in cerebral ischemia/reperfusion injury through targeting m6A reader YTHDF1 to inhibit p65 mRNA translation[J]. Int Immunopharmacol,2020,88:106937. doi:10.1016/j.intimp.2020.106937.
[40] TAN C,WU Q,WANG H,et al. Dysbiosis of gut microbiota and short-chain fatty acids in acute ischemic stroke and the subsequent risk for poor functional outcomes[J]. JPEN J Parenter Enteral Nutr,2021,45(3):518-529. doi:10.1002/jpen.1861.
[41] LIAO S,WU J,LIU R,et al. A novel compound DBZ ameliorates neuroinflammation in LPS-stimulated microglia and ischemic stroke rats: role of Akt(Ser473)/GSK3β(Ser9)-mediated Nrf2 activation[J]. Redox Biol,2020,36:101644. doi:10.1016/j.redox.2020.101644.
[42] QI L,HU H,WANG Y,et al. New insights into the central sympathetic hyperactivity post-myocardial infarction: roles of METTL3-mediated m(6)A methylation[J]. J Cell Mol Med,2022,26(4):1264-1280. doi:10.1111/jcmm.17183.
[43] HU W,XIE H,ZENG Y,et al. N6-methyladenosine participates in mouse hippocampus neurodegeneration via PD-1/PD-L1 pathway[J]. Front Neurosci,2023,17:1145092. doi:10.3389/fnins.2023.1145092.
[44] JIANG Z,SHI L,HUANG H,et al. Downregulated FTO promotes microRNA-155-mediated inflammatory response in cerebral ischemia/reperfusion injury[J]. Neuroscience,2023:526:305-313. doi:10.1016/j.neuroscience.2023.07.012.
[45] LIU Q,BHUIYAN M I H,LIU R,et al. Attenuating vascular stenosis-induced astrogliosis preserves white matter integrity and cognitive function[J]. J Neuroinflammation,2021,18(1):187. doi:10.1186/s12974-021-02234-8.
[46] COCKOVA Z,HONC O,TELENSKY P,et al. Streptozotocin-induced astrocyte mitochondrial dysfunction is ameliorated by FTO inhibitor MO-I-500[J]. ACS Chem Neurosci,2021,12(20):3818-3828. doi:10.1021/acschemneuro.1c00063.
[47] HUANG R,ZHANG Y,BAI Y,et al. N(6)-methyladenosine modification of fatty acid amide hydrolase messenger RNA in circular RNA STAG1-regulated astrocyte dysfunction and depressive-like behaviors[J]. Biol Psychiatry,2020,88(5):392-404. doi:10.1016/j.biopsych.2020.02.018.
[48] LI Z,MONIRUZZAMAN M,DASTGHEYB R M,et al. Astrocytes deliver CK1 to neurons via extracellular vesicles in response to inflammation promoting the translation and amyloidogenic processing of APP[J]. J Extracell Vesicles,2020,10(2):e12035. doi:10.1002/jev2.12035.
[49] OUYANG F,JIANG Z,CHEN X,et al. Is cerebral amyloid-β deposition related to post-stroke cognitive impairment?[J]. Transl Stroke Res,2021,12(6):946-957. doi:10.1007/s12975-021-00921-5.
(2023-05-23收稿 2023-08-01修回)
(本文編辑 李志芸)
