吉兰-巴雷综合征患者外周血记忆性T 细胞和记忆性B 细胞亚群的变化及其临床意义
2024-03-16刘坛黄小雨张勇
刘坛,黄小雨,张勇
1.湖州市第一人民医院神经内科,浙江湖州 313000;2.徐州医科大学附属医院神经内科,江苏徐州 221000
吉兰-巴雷综合征(Guillain-Barré syndrome,GBS)是一类免疫介导的急性炎性周围神经病,主要病理生理机制是外周神经中抗体-抗原相互作用引起的补体介导的神经损伤[1-2]。既往研究发现体液免疫及细胞免疫共同参与此病的发生,然而国内外关于GBS 免疫细胞亚群的研究尚不充分,尤其是记忆性细胞亚群在GBS 患者外周血中的占比仍需进一步探讨。已有文献报道记忆性B 细胞参与了GBS 的发病[3];也有学者指出记忆性T 细胞可帮助记忆性B 细胞更快速地产生抗体反应[4]。由此推测记忆性T 细胞及记忆性B 细胞均可能参与GBS 的发病。本研究通过分析GBS 患者记忆性T细胞及记忆性B 细胞亚群的改变,为探索GBS 的发病机制提供新思路。
1 资料与方法
1.1 一般资料
回顾性选取2018 年6 月至2020 年12 月于徐州医科大学附属医院神经内科住院的16 例GBS 患者作为研究组,其中男8 例、女8 例,年龄28~78岁,平均(54.38±15.24)岁。选取同期16 名来院的健康体检者作为对照组,其中男7 例、女9 例,年龄29~69 岁,平均(46.56±11.18)岁。纳入标准:①符合《中国吉兰-巴雷综合征诊治指南2019》诊断标准[1];②年龄≥18 岁;③未接受静脉注射免疫球蛋白治疗。排除标准:①其他自身免疫性疾病,如白血病、类风湿疾病、甲状腺疾病、人类免疫缺陷病毒感染的患者;②器官移植的患者;③诊断不明确或临床资料不全、资料丢失的患者。所有研究对象均签署知情同意书。本研究经徐州医科大学附属医院伦理委员会审批通过(伦理审批号:XYFY2020-JS019-01)。
1.2 方法
所有研究对象均于入组后空腹抽取6ml 静脉血,其中2ml 静脉血用乙二胺四乙酸(ethylene diamine tetraacetic acid,EDTA)抗凝,余静脉血不抗凝直接分离血清。采用人淋巴细胞分离液(天津灏洋生物公司)分离得到外周血单个核细胞(peripheral blood mononuclear cell,PBMC),经磷酸缓冲盐溶液(phosphate buffer saline,PBS)洗涤离心后加入鼠抗人CD3-APC-Cy7 抗体、鼠抗人CD4-APC 抗体、鼠抗人CD8-Percp-Cy5-5 抗体、鼠抗人CD45RAV450 抗体、鼠抗人CCR7-BV510 抗体(美国BD 公司);同时设同型对照管,室温避光孵育15min 后,流式细胞仪检测记忆性T 细胞亚群,其中CD4+初始T 细胞(naive T cells,TN)表型为CD3+、CD4+、CD8-CD45RA+、CCR7+,CD4+中央记忆性T 细胞(central memory T cells,TCM)表型为CD3+、CD4+、CD8-CD45RA-、CCR7+,CD4+效应记忆性T 细胞(effector memory T cells,TEM)表型为CD3+、CD4+、CD8-CD45RA-、CCR7-,CD4+终末分化效应记忆性T细胞(terminally differentiated effector memory T cells,TEMRA)表型为CD3+、CD4+、CD8-CD45RA-、CCR7-;CD8+TN 细胞表型为CD3+、CD4-、CD8+CD45RA+、CCR7+,CD8+TCM 细胞表型为CD3+、CD4-、CD8+CD45RA-、CCR7+,CD8+TEM 细胞表型为CD3+、CD4-、CD8+CD45RA-、CCR7-,CD8+TEMRA细胞表型为CD3+、CD4-、CD8+CD45RA-、CCR7-。在分离的PBMC 中加入鼠抗人CD45-Percp-Cy5-5 抗体、鼠抗人CD19-APC、鼠抗人CD27- BV510、鼠抗人CD38-FITC 抗体(美国BD 公司),同时设同型对照管。室温避光孵育15min 后,流式细胞仪检测记忆B 细胞亚群,其中记忆B 细胞表型为CD19+CD27+,浆母细胞表型为CD19+CD27+CD38+。
1.3 统计学方法
采用SPSS 19.0 统计学软件对数据进行处理分析。符合正态分布的计量资料用均数±标准差(±s)表示,组间比较采用独立样本t检验。不符合正态分布的计量资料以中位数(四分位数间距)[M(Q1,Q3)]表示,组间比较采用秩检验。计数资料以例数(百分率)[n(%)]表示,组间比较采用X2检验。采用Pearson 相关性分析CD4+TEM及CD8+TEM 占比与Hughes 残疾评分、脑脊液蛋白、脑脊液免疫球蛋白G(immunoglobulin G,IgG)及记忆性B 细胞的相关性。P<0.05 为差异有统计学意义。
2 结果
2.1 GBS 患者的一般资料及外周血记忆性T 细胞亚群占比分析
GBS 患者的一般资料分析见表1。与对照组相比,研究组患者的CD4+TN 及CD8+TN 均明显下降(P<0.05),CD4+TEM 及CD8+TEM 均明显升高(P<0.05),TCM 及TEMRA 在CD4 和CD8 上的比例差异无统计学意义(P>0.05),见表2。
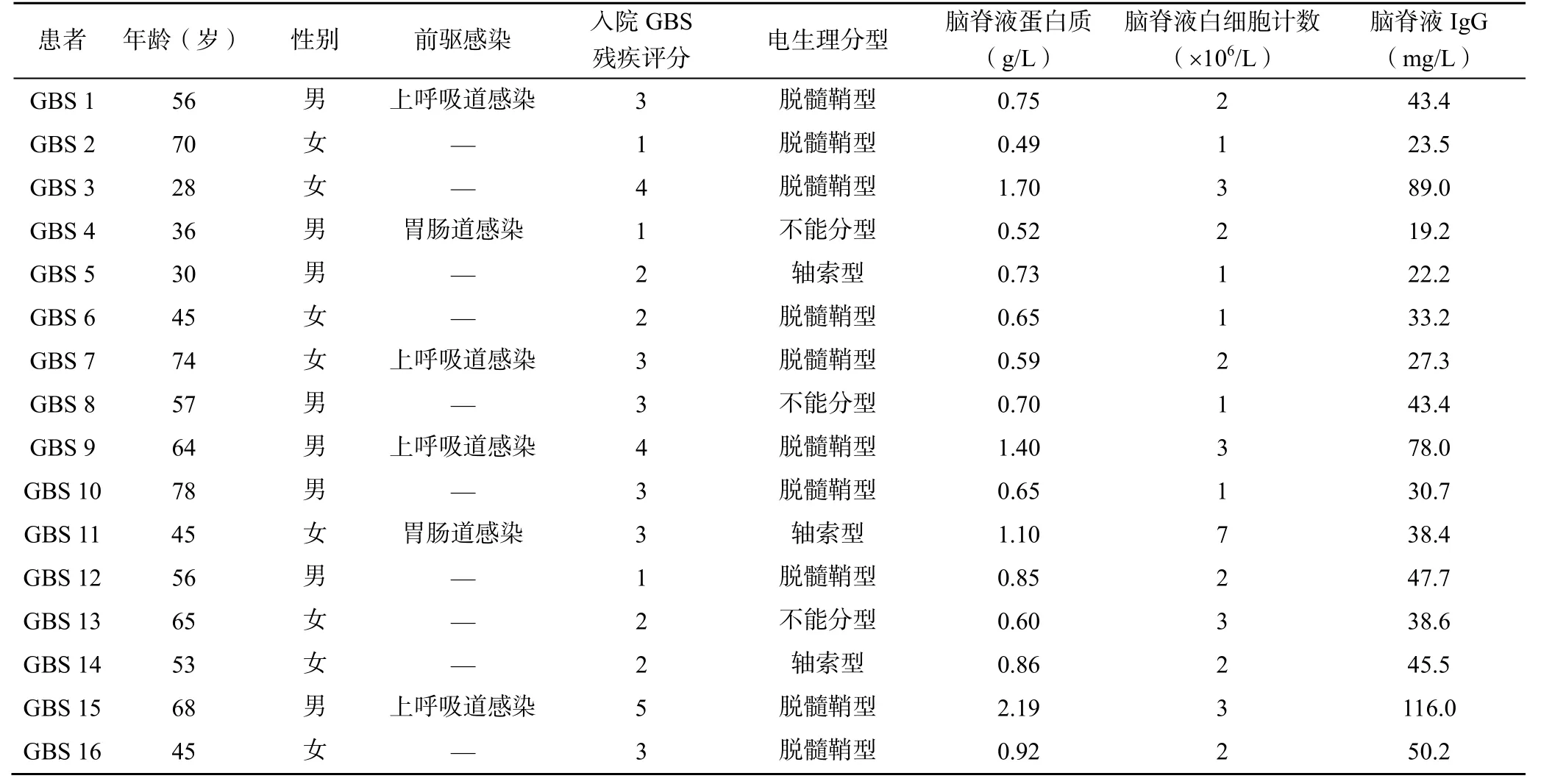
表1 GBS 患者的一般资料

表2 两组患者的外周血记忆性T 细胞亚群占比比较(n=16)
2.2 两组患者的外周血记忆性B 细胞占比比较
与对照组比较,研究组患者的记忆性B 细胞比例明显升高(P<0.05),浆母细胞的比例差异无统计学意义,见表3。

表3 两组患者的外周血记忆性B 细胞占比比较(n=16)
2.3 GBS 患者外周血CD4+TEM 和CD8+TEM 占比与GBS 患者临床严重程度的相关性分析
GBS 患者外周血CD4+TEM 及CD8+TEM 占比与Hughes 残疾评分、脑脊液蛋白、脑脊液IgG 及记忆性B 细胞占比均呈正相关(P<0.05),见图1。

图1 GBS 患者外周血TEM 占比与GBS 临床严重程度相关性分析
3 讨论
GBS 发病与自身免疫功能障碍相关,各种类型的活化的T 细胞被认为是GBS 的发生和发展的协调者。研究表明记忆性T 细胞对B 细胞生发中心的形成及产生对抗髓鞘的抗体具有十分重要的作用,其中 TEM 细胞可通过产生细胞因子白细胞介素(interleukin-17,IL-17)和重组人干扰素(recombinant human interferon-r,IFN-γ)等炎症因子参与B 细胞的分化[5];同时记忆性B 细胞也与GBS 的发病相关[3]。因此,进一步明确记忆性B 细胞亚群、记忆性T 细胞亚群尤其是TEM 细胞在GBS 患者外周血中的占比,对解释GBS 的发病机制至关重要。
有学者根据是否表达CD45RA 区别初始T 细胞和记忆性T 细胞,并进一步依据趋化因子受体CCR7的表达将记忆性T 细胞分为TCM 和TEM[6]。与初始T 细胞和TCM 细胞相比,TEM 细胞更能快速被抗原识别,迅速激活免疫反应,且可立刻到达炎症部位并发挥作用[7]。既往多项研究表明,TEM 细胞与一些自身免疫性疾病如多发性硬化、系统性红斑狼疮 、结肠炎、1 型糖尿病等的发生、发展、复发及严重程度密切相关[8-12]。本研究结果也显示,与对照组相比,研究组患者外周血 CD4+TEM 细胞和CD8+TEM 细胞占比更高,差异有统计学意义。初始B 细胞成熟后,与抗原相遇被其激活,随后分化为CD19+CD27+记忆性B 细胞;当记忆性B 细胞再次与相同抗原相遇时就会增殖,分化为浆细胞,产生特异性抗体,从而发挥介导体液免疫应答的作用。据报道,记忆性B 细胞在一些自身免疫性疾病如系统性红斑狼疮、类风湿关节炎、自身免疫性溶血性贫血等疾病中发挥至关重要的作用[13-15]。Wang 等[3]发现记忆B 细胞在GBS 急性期显著增加,并随着病情的恢复而减少,进一步的Spearman 相关分析显示GBS 的临床严重程度与患者外周血中记忆B 细胞的占比呈正相关。本研究结果与之一致,研究组患者记忆性B 细胞比例高于对照组;此外,进一步Pearson相关性分析显示CD4+TEM 及CD8+TEM 细胞占比与Hughes 残疾评分、脑脊液蛋白、脑脊液IgG 及记忆性B 细胞占比均呈正相关,CD4+TEM 及CD8+TEM细胞与疾病临床严重程度呈正相关。在GBS 发病中发挥重要作用,并可能通过自身效应功能调节B 细胞的分化参与GBS 的发病。
综上,本研究发现GBS 患者存在记忆性T 细胞和记忆性B 细胞亚群免疫紊乱,外周血中CD4+TEM细胞、CD8+TEM 细胞及记忆性B 细胞占比升高可能是GBS 发病过程中的一个外周免疫机制。进一步深入探讨TEM 细胞参与GBS 发病的机制有望成为治疗GBS 的新靶点。
利益冲突:所有作者均声明不存在利益冲突。
