3个李品种(系)芽需冷量的研究及休眠相关DAM基因的表达分析
2024-03-09唐孝富孟银银杜莹蒲靖王苗田锟邓群仙张慧芬
唐孝富,孟银银,杜莹,蒲靖,王苗,田锟,邓群仙,张慧芬
(四川农业大学园艺学院,成都 611130)
休眠是高等植物经过长期演化而获得的一种对环境条件及季节变化的生物学适应性,直接与开花、萌芽相关[1-2]。Lang G.A.等[3]于1987年对休眠进行了重新定义和分类,基于引起休眠的因素将其分为三大类:类休眠,指休眠结构以外的生理因素(如顶端优势、激素等)诱导产生的生长停滞现象;内休眠,指休眠结构本身的因素(如冷温需求、光周期影响)控制的生长停滞现象;生态休眠,是指由环境因子(如温度、水分胁迫、营养亏缺等)引起的休眠,一旦环境条件适宜,植物便可重新恢复生长。多年生果树通过芽休眠抵御自然界不利环境,而芽一般需要达到一定低温需冷量才能解除休眠进行正常生长发育[4-6]。陈茂铨等[7]对12个桃品种的花芽休眠需冷量进行研究发现,不同品种需冷量差异较大,其中高需冷量品种在犹他模型下达900~950 CU。近几年,汤志洪[8]、陈登文等[9]研究欧李冬季休眠特性发现,欧李花芽的需冷量低于叶芽,花芽与叶芽自然休眠过程中对有效低温的累积要求存在差异。
休眠是多基因协同控制的过程,近年来,关于芽休眠相关基因的研究较多。D.G.Blelenberg等[10]最早在桃的休眠障碍型突变体上发现MIKC-type MADS-box转录因子控制了芽休眠和分生组织生长,将其命名为dormancy associated MADS-box gene(DAM)[11-12]。DAM基因与植物体的开花及花器官的形成有密切关系,在桃、梨等蔷薇科果树中研究广泛[13-15],李海炎等[16]在桃的DAM基因调控需冷量的研究中发现,不同需冷量的桃在休眠过程中DAM基因的表达模式一致,随休眠解除DAM表达逐渐下降,且DAM基因在高需冷量的品种的表达量高于低需冷量品种的表达。杨高宇等[17]利用PacBio和high-c技术构建低需冷量梨品种的基因组组合,进一步确定梨的DAM基因对芽萌动的抑制作用。杨依维等[18]对欧李休眠诱导的分子机制研究发现,DAM5基因的表达随着休眠诱导进程逐渐升高,DAM5基因在诱导欧李芽休眠中发挥关键作用。近年来有研究者针对李的花芽休眠相关MADS-box基因的克隆及其对花芽发育和休眠的影响进行研究,结果表明,DAM转录因子在温暖季节开始积累,且随着温度的降低逐渐增加,从而诱导芽休眠[19]。
李为蔷薇科李属植物,作为一种重要的经济果树,在我国广泛种植,其中四川省主栽品种主要为‘羌脆李’和‘脆红李’。李树需冷量与芽休眠的解除是影响开花坐果的重要因素。本研究以‘脆红李’‘羌脆李’及‘羌脆李’的晚熟变异材料“晚熟羌脆李”为研究材料,对花芽与叶芽需冷量的进行比较,并用生物信息学方法对李DAM基因的序列结构和芽不同休眠状态的表达量分析,为四川李栽培生产中芽休眠调控奠定理论基础。
1 材料和方法
1.1 试验材料
‘羌脆李’‘脆红李’及“晚熟羌脆李”枝条与芽均采样自四川省阿坝藏族羌族自治州汶川县茅岭村(31°28′ N,103°33′ E)果园。“晚熟羌脆李”为引种定植的100株‘羌脆李’中发现的芽变优株,该优株和同一批定植的‘羌脆李’相比,果实晚成熟约30 d左右。成熟期与当地‘脆红李’基本一致。“晚熟羌脆李”‘羌脆李’以及‘脆红李’芽休眠状态观察于2020年11月李全树落叶后开始,至2021年1月全部观察芽解除芽休眠,观察期间每间隔14 d从挂牌李树上选取生长发育良好、芽体饱满的1年生枝条5枝,用于花芽、叶芽取样,在液氮中预冷后立即放入-80 ℃超低温冰箱中保存用于RNA提取。
在NCBI网站下载与李相近的同源物种桃的DAM1-6基因序列,本地Blast在欧李基因组中查找到6个李DAM基因,分别命名为PdDAM1、PdDAM2、PdDAM3、PdDAM4、PdDAM5和PdDAM6。
1.2 萌芽率统计
2020年11月—2021年1月芽休眠观察期间,每隔14 d从3个品种(系)李树上各选取生长发育良好、芽体饱满的1年生枝条6枝,用于人工培养,统计萌芽率[20-21],每次采样各品种(系)用于观察统计的花芽与叶芽数量均为60个以上。将采回的枝条基部斜剪45°,放在盛有清水的烧杯中,以淹没枝条基部,立即放入人工气候箱培养。培养条件为:光照12 h,光照强度320 μmol/(m2·s),黑暗12 h;温度为(25±1.0)℃/(18±1.0)℃;空气相对湿度为75%;每2 d换1次水,每次换水剪除枝条基部少许。培养21 d时统计萌芽率,以芽顶端开裂露绿为萌芽标准,以萌芽率>50%时的观测日期为解除内休眠的日期[22-23]。参照Lang G.A.等[3]的方法[3],确定休眠的阶段:以萌芽率为0时,芽完全处于内休眠阶段;萌芽率为0~50%时,芽处于内生态休眠转换阶段;萌芽率为≥50%时,即认定芽解除内休眠进入生态休眠阶段。
1.3 需冷量计算
观察期间于田间装置温湿度计,设置为每隔15 min读数1次,实时记录田间温湿度。分别统计7.2 ℃模型、0~7.2 ℃模型和犹他模型的低温累积数。
7.2 ℃模型[24]:低温积累量以日平均温度稳定通过7.2 ℃的日期为起点,计算小于7.2 ℃低温小时数(chilling hour,CH),当每小时温度≤7.2 ℃时,计1个低温小时数。
0~7.2 ℃模型[25]:低温积累量以日平均温度稳定通过7.2 ℃的日期为起点,计算0~7.2 ℃的低温小时数(chilling hour,CH),当每小时温度处于0~7.2 ℃范围内,计1个低温小时数。
犹他模型[26]:低温积累量以负累积低温达到最大值时的日期为起点计算积累的冷温单位(chilling unit,CU),每小时温度与冷温单位转化如表1所示。

表1 温度冷温单位转化表Table 1 Transformation of temperature and chilling unit
1.4 李DAM基因进化树和保守motif分析
利用MEMEM在线软件对李DAM蛋白的保守基序进行分析,基序的最大数目设置为10,TBtools软件[27]对DAM基因结构进行分析与作图。利用Clusta1X进行氨基酸序列比对,MEGA11对李与其他植物的DAM氨基酸序列构建进化树。
1.5 李DAM 基因qRT-PCR分析
3个品种(系)李花芽、叶芽RNA的提取参照宣继萍等[28]的CTAB法[28]。cDNA的合成使用Goldenstar™RT 6 cDNA Synthesis Ver.2(北京擎科生物公司)反转录试剂盒,具体操作参照说明书。引物设计采用Beacon Designer 7 (Version 7.9, Premier Biosoft, San Francisco, USA),具体引物信息见表2。以李TUB3为内参基因。使用BIO-RAD CFX96定量PCR仪进行qRTPCR 反应:体系为10 μL,包含2× TSINGKE®Master qPCR Mix (SYBR GreenⅠ) 5 μL,上下游引物(10 μmol/L)各0.4 μL,cDNA2 μL,ddH2O 2.2 μL。2步法扩增条件为:95 ℃预变性1 min,95 ℃变性10 s,60 ℃退火及延伸30 s,共40个循环,每个处理包含3个重复。采用2-∆∆Ct法计算基因相对表达量。
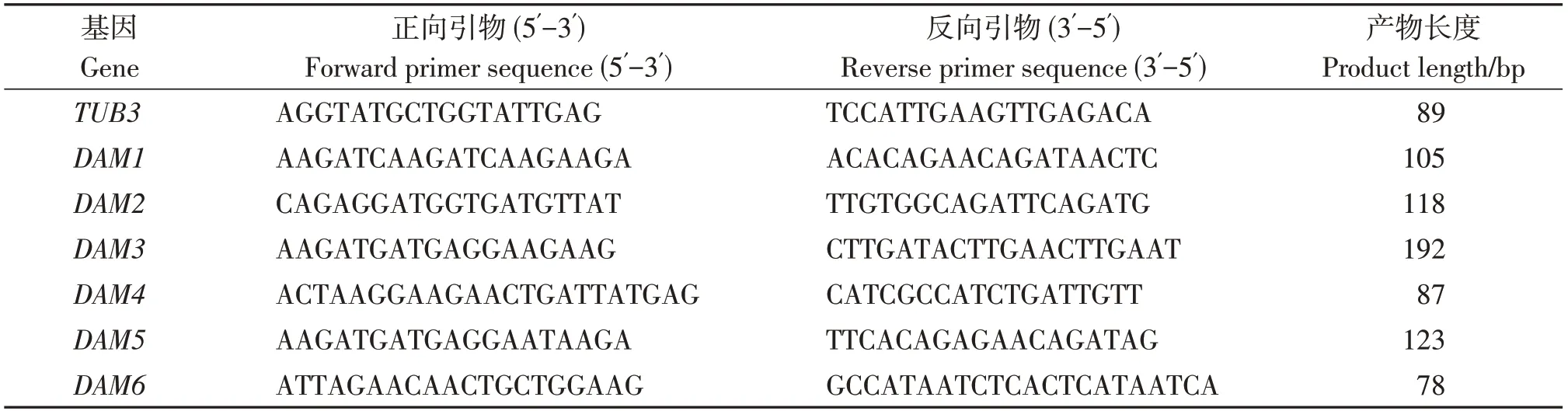
表2 DAM 基因qRT-PCR 引物序列Table 2 Primer sequences of DAM gene for real-time quantitative PCR
1.6 数据处理分析
使用Microsoft Excel 2021和Graph Pad Prism 8进行数据处理和作图。
2 结果与分析
2.1 萌芽率统计
由图1可知,“晚熟羌脆李”与‘羌脆李’‘脆红李’的花芽萌芽率差异较大,“晚熟羌脆李”花芽从11月5日—12月5日萌芽率未达到50%,为内休眠状态,12月20日时为生态休眠阶段,萌芽率为84.0%。‘羌脆李’‘脆红李’花芽萌芽率较一致,在11月5日到11月20日萌芽率小于50%,该阶段处于内休眠状态,12月5日开始萌芽率大于50%并且之后逐渐上升,达到解除内休眠的标准,处于生态休眠阶段,12月5日萌芽率高低依次为‘羌脆李’>‘脆红李’>“晚熟羌脆李”,“晚熟羌脆李”花芽的内休眠时间最长。3个品种(系)叶芽萌芽率相似,均表现为11月5日萌芽率小于50%,处于内休眠状态,从11月20日开始叶芽萌芽率大于50%内休眠状态解除。3个品种(系)叶芽休眠解除临界期均为11月5日—11月20日。3个品种(系)花芽休眠解除临界期不同,“晚熟羌脆李”为12月5日至12月20日,‘羌脆李’和‘脆红李’均为11月20日至12月5日,3个品种(系)叶芽解除临界期相同,为11月5日至11月20日。
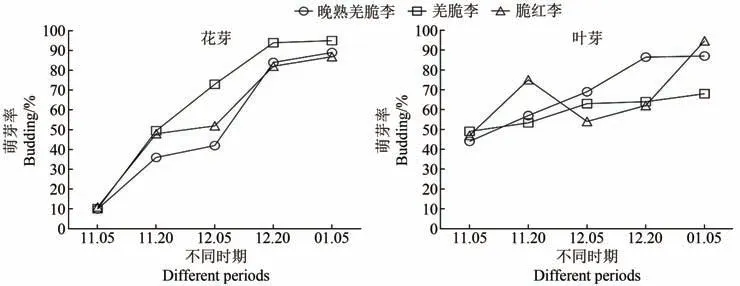
图1 花芽、叶芽不同时期萌芽率Figure 1 Germination rate of flower buds and leaf buds at different periods
2.2 休眠芽低温需冷量分析
结合田间温湿度记录仪温度记录与室内培养箱萌芽率统计,使用7.2 ℃模型、0~7.2 ℃模型和犹他模型分别计算3个李品种(系)的低温需冷量(表3)。结果表明,3种计算模型下,‘羌脆李’与‘脆红李’需冷量均相同且低于“晚熟羌脆李”,7.2 ℃模型下花芽需冷量最高,“晚熟羌脆李”需冷量为943 CH,‘羌脆李’与‘脆红李’需冷量为630 CH。同一模型下3个品种(系)叶芽需冷量均相同。“晚熟羌脆李”、‘羌脆李’和‘脆红李’的花芽、叶芽解除休眠的低温需冷量所反映的物候学模型与三者芽休眠状态调查结果一致。

表3 3 个李品种(系)低温需冷量Table 3 Low temperature chilling requirement of 3 plum materialsCH
2.3 李DAM基因进化树和保守motif分析
利用李(Prunus domestica)、桃(Prunus persica)、拟南芥(Arabidopsis thaliana)、杏(Prunus armeniaca)、梅(Prunus mume)、苹果(Malus pumila)、葡萄(Vitis vinifera)和杨树(Populus talassica)DAM蛋白序列构建系统发育树发现,8个物种35个蛋白可聚类为3个亚族,李6个DAM在3个亚族内均有分布。将李DAM家族的保守基序及其系统进化树使用TBtools软件进行整/合可视化作图(图2)。系统进化分析发现,李的PdDAM1、PdDAM2、PdDAM3、Pd-DAM4、PdDAM5、PdDAM6分别与PpDAM6、Par-DAM5、PtMADS5、PmDAM3、PpDAM1、MdDAM1进化关系最近,序列相似性最高。MEME 网站在线分析预测35个DAM蛋白的保守motif,除苹果3个DAM与梅PmDAM5外,其他序列均含4个相对保守的motif: motif9-motif1-motif3-motif2,且motif之间的排列具有一定的规律性,6个李DAM蛋白序列motif排列顺序均为: motif9-motif1-motif3-motif5-motif2-motif6-motif4-motif8-motif7-motif10,而Pd-DAM4缺少motif8和motif7,PdDAM2基因缺少1个motif6。这些结果表明李DAM家族成员在序列较相似,PdDAM4与PdDAM2出现1~2个基因出现保守基序缺失或增加的现象,可能是进化过程中出现了序列结构差异。
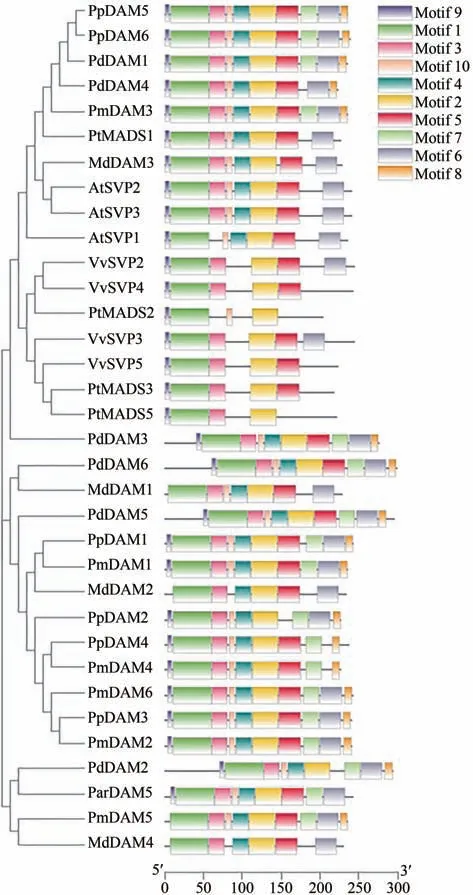
图2 李DAM基因进化树及保守基序Figure 2 Prunus DAM gene evolutionary tree and conserved motif order
2.4 休眠相关DAM基因qRT-PCR分析
休眠相关基因DAM在“晚熟羌脆李”‘羌脆李’和‘脆红李’休眠期花芽与叶芽的相对表达量的变化如(图3)所示,3个李品种(系)的花芽与叶芽内休眠解除临界期表达基因相同,且在内休眠解除临界期DAM基因的相对表达量基本呈下降趋势。“晚熟羌脆李”花芽与叶芽内休眠解除临界期均表现为DAM4和DAM5表达最高,其中DAM5在花芽与叶芽中的表达均呈下降趋势,而DAM4在花芽中为下降趋势,在叶芽中却呈缓慢上升趋势;‘羌脆李’花芽与叶芽内休眠解除临界期内随着休眠的解除DAM1和DAM5表达最高,呈逐渐下降趋势;‘脆红李’休眠解除临界期DAM2和DAM5表达最高,且呈逐渐下降趋势。总体而言,无论是“晚熟羌脆李”‘羌脆李’还是‘脆红李’的花芽还是叶芽在达到解除内休眠的过渡阶段均有DAM5为主要的调控基因,而在内休眠转变为生态休眠的阶段,DAM基因表达量均显著下调,DAM基因表达下降与休眠解除密切相关。

图3 “晚熟羌脆李”花芽(A1)、“晚熟羌脆李”叶芽(A2)、‘羌脆李’花芽(B1)、‘羌脆李’叶芽(B2)、‘脆红李’花芽(C1)、‘脆红李’叶芽(C2)中DAM基因的表达分析Figure 3 Expression analysis of DAM genes of "Wanshuqiangcuili" flower buds (A1), "Wanshuqiangcuili" leaf buds (A2),'Qiangcuili' flower buds (B1), 'Qiangcuili' leaf buds (B2), 'Cuihongli' flower buds (C1), and 'Cuihongli' leaf buds (C2)
3 讨论与结论
落叶果树具有明显的季节性生长规律,休眠为果树生长发育过程中的重要时期[29-30]。休眠除了受外界环境的影响之外,通常还受解除休眠需冷量、休眠相关DAM基因的多重控制[31]。一般不同的树种,或者同一树种不同品种品系的低温需冷量不同,休眠期时间长短也不一致,如柿、苹果等树种之间休眠不同[32-35]。枣树研究中,低温需冷量较小的“京枣39”在12月上旬结束休眠,“辣椒枣”“七月鲜”等低温需冷量中等的在1月下旬结束休眠,低温需冷量较大的“大树冬枣”在2月初结束休眠[36]。本研究中3个李品种(系)花芽需冷量不同,“晚熟羌脆李”较‘羌脆李’需冷量高,果实成熟期也延迟。陈菲菲[37]在桑葚休眠与果实熟期的研究结果也表明,不同休眠时期对产量无显著影响,但休眠提前解除,果实熟期也提前。因此在生产上利用休眠期的合理管理来进行产期的调控。本研究对3个不同李品种(系)的花芽和叶芽的休眠状态、低温需冷量分析发现,“晚熟羌脆李”花芽从12月20日萌芽率大于50%,解除了内休眠,‘羌脆李’和‘脆红李’在12月5日处于解除深休眠状态;对于叶芽,3个品种(系)均在11月20日达到解除内休/眠状态。需冷量的差异与休眠状态一致,且3者叶芽需冷量无差异,而解除休眠最晚的“晚熟羌脆李”花芽的需冷量最高。
DAM基因是果树休眠变化的重要调控基因,在桃树中发现存在6个DAM基因[10],中国樱桃中发现3个DAM基因[13],本研究通过与李近缘物种桃的DAM1-6比对,以欧李基因组为参考查找到6个李DAM基因。H.Yamane等[38-39]研究了需冷量高低的不同品种的桃,结果表明,休眠期PpDAM5和Pp-DAM6均存在相对较高的表达。本研究发现,无论是“晚熟羌脆李”‘羌脆李’还是‘脆红李’,均表现为随休眠状态解除DAM的相对表达逐渐下调,在休眠解除临界期表达下调最显著,且DAM5可能是3个品种(系)中共有的调控休眠的关键基因,这与欧李DAM5促进休眠,抑制萌发[18]研究结果一致。“晚熟羌脆李”花芽中的DAM基因表达高峰均晚于‘羌脆李’与‘脆红李’,与花芽需冷量及休眠观察结果一致。需冷量与DAM表达分析发现,“晚熟羌脆李”的内休眠解除要晚于‘羌脆李’,这也可能是“晚熟羌脆李”较‘羌脆李’果实熟期推迟的原因之一,休眠解除较迟,萌动也会相应有所推迟,同时树体所消耗的营养也随之增多,体内提供生长养分变少,影响花期、坐果甚至果实熟期[40]。
