盐胁迫对杠柳幼苗生长及次生代谢产物积累的影响
2024-03-08韩翠婷李先宽王广苹
韩翠婷,李先宽,王广苹,马 琳,张 坚
(天津中医药大学中药学院,天津 301617)
杠柳(PeriplocasepiumBunge)为萝藦科杠柳属植物,其根皮为传统中药“香加皮”,具有祛风湿、强筋骨的功效[1]。现代研究表明,杠柳的主要有效成分为甾体类成分,如强心苷类、C21甾类等,具有强心、抗肿瘤等药理作用[2]。此外,杠柳中的挥发性成分,如4-甲氧基水杨醛、异香草醛等也是杠柳的活性成分,其中4-甲氧基水杨醛还是药典中杠柳的唯一质量控制指标性成分[1]。
土地盐渍化是全世界面临的重要环境问题,据统计,目前全球约有9.5亿hm2的盐渍化土地,这已成为限制植物生长发育的主要非生物胁迫之一[3]。因此,探究盐渍化土壤对植物生长特性的影响,对盐渍化土壤的开发利用和土壤改良具有重要意义。前人研究表明,种植耐盐碱植物,可以在一定程度上降低土壤盐分,达到改良盐碱地的目的[4]。同时,有研究者提出中药材种植的“边缘效应”,表明适度的逆境胁迫能够明显提高药材的活性成分含量[5]。杠柳作为耐盐碱的水土保持物种,近年来对其研究多见于干旱或沙埋深度等胁迫下的杠柳生理特性及光合作用特征等[6,7]方面,而鲜见杠柳在盐胁迫下的应答机制及产量品质等相关报道。研究盐胁迫对药用植物次生代谢成分的影响有利于揭示逆境条件下其活性成分的合成机制,并为盐胁迫作为诱导手段刺激药用植物有效成分合成提供理论指导[8]。因此,本试验通过盐胁迫处理,考察杠柳幼苗的生长特性以及次生代谢产物总甾体、杠柳毒苷、杠柳次苷、杠柳苷元和4-甲氧基水杨醛等含量变化,揭示盐逆境与杠柳次生代谢产物积累的相关性,以期为杠柳的盐胁迫研究及产量品质调控提供参考依据。
1 材料和方法
1.1 供试材料与试验设计
试验于天津中医药大学中药学院进行,种子采自于天津市蓟州区山区,经笔者张坚副教授鉴定为萝藦科植物杠柳(PeriplocasepiumBunge)种子。选择适量均一饱满的种子,经催芽处理后均匀种植到育苗穴盘中,育苗基质由蛭石、珍珠岩(1∶1)混合而成,共15盆。置于光周期(光照/黑暗)16 h/8 h、光照度3 600 lx、恒温 (23±2) ℃、相对湿度为 (50±5)%的培养室内培养。每周喷施1/2 MS营养液1次,定期浇水。期间进行间苗处理,30 d后选取生长良好、长势一致的幼苗进行盐胁迫处理。
将15盆幼苗随机分为5个处理组,每个处理组3盆。盐胁迫浓度设置0.2%、0.5%、1.0%、2.0%等4个水平,同时设置空白对照组(CK)。其中,盐处理液由1/2 MS营养液和分析纯NaCl配制,空白对照组浇灌等量1/2 MS营养液。每隔3 d灌溉1次处理液,并使大量处理液从盆底渗出,以交换基质中的积余盐。胁迫处理10 d后,收获样品,记录不同处理组地上及地下部分的鲜质量数据,经真空冷冻干燥处理后记录其干质量,计算干湿比(干质量/鲜质量×100%),干湿比越大,代表植株含水量越大,反之则越小。同时将样品置于-20 ℃冰箱保存,用于后续各成分含量测定。
1.2 药用成分含量测定
杠柳中总甾体成分含量测定采用香草醛-浓硫酸法[9];杠柳毒苷、杠柳次苷、杠柳苷元及4-甲氧基水杨醛含量测定采用UPLC 法,具体实验条件参照姚鹏等[10]方法并进行改良。
1.2.1 UPLC色谱条件
色谱柱为ACQUITY UPLC BEH C18(50 mm×2.1 mm,1.7 μm),保护柱为Waters C18 柱(12.5 mm×2.1 mm,5 μm),以纯水(A)-甲醇(B)溶液为流动相梯度洗脱(0 min,90%A;1~4 min,85%~70%A;4~8 min,70%~67%A;8~9 min,67%~55%A;9~11 min,55%~48%A;11~17 min,48%~25%A;17~18 min,25%~90%A),流速为0.3 mL/min,柱温为30 ℃,进样体积为1 μL,检测波长为235 nm,分析时间为18 min。
1.2.2 对照品溶液制备
精密称取各对照品适量,加甲醇溶解并定容至5 mL,即得对照品母液。各成分质量浓度分别为杠柳毒苷1.02 mg/mL、杠柳次苷1.06 mg/mL、杠柳苷元0.94 mg/mL、4-甲氧基水杨醛1.02 mg/mL。然后,精密吸取各对照品母液适量,用甲醇配制成每1 mL分别含204 μg杠柳毒苷、212 μg杠柳次苷、188 μg杠柳苷元、204 μg 4-甲氧基水杨醛的混合对照品储备液,4 ℃冰箱存储备用。
1.2.3 供试品溶液制备
分别称取不同处理组杠柳地上及地下部分干燥粉末约 0.1 g,置于5 mL容量瓶中,用纯甲醇定容,超声提取45 min,静置冷却后,纯甲醇补足体积,8 000 r/min离心2次,每次10 min,上清液即为供试品溶液,过0.22 μm微孔滤膜后,于4 ℃冰箱存储备用。
1.2.4 方法学考察
(1)线性关系考察:精密吸取一定量的混合对照品储备液,按照不同稀释倍数用纯甲醇制成一系列浓度的混标对照品溶液,按照1.2.1节色谱条件依次进样,测定峰面积,以浓度(μg/mL)为横坐标(x),峰面积为纵坐标(y),绘制标准曲线(表1)。由表1可知,4 个化合物在各自线性范围内决定系数(R2) 均大于0.999 0,表明4个化合物在其线性范围内线性关系良好。

表1 杠柳毒苷、杠柳苷元、杠柳次苷及4-甲氧基水杨醛的标准曲线Table 1 Standard curves of periplocin,periplocymarin,periplogenin,and 4-methoxy salicylaldehyde
(2)精密度实验:在1.2.1节色谱条件下,取杠柳毒苷对照品溶液,连续进样6次,分别测定其峰面积。结果显示,其峰面积RSD 值均小于2%,表明仪器精密度良好。
(3)重复性实验:取同一处理组样品(0.5%盐浓度杠柳幼苗根)约0.1 g,按照1.2.3节方法制备供试品溶液,平行操作6份,在1.2.1节色谱条件下依次进样,测定峰面积。结果显示,各成分RSD 值均小于2%,表明本方法重复性良好。
(4)稳定性实验:按照1.2.3节方法制备供试品溶液(0.5%盐浓度杠柳幼苗根),分别于0,2,4,6,8,12,24 h内进样,测定峰面积。结果显示各成分RSD均小于2%,表明供试品溶液在24 h内基本稳定。
(5)加样回收率实验:取已知含量的同一处理组杠柳样品粉末,平行取样6份,分别加入对照品溶液适量,按上述提取方法制备提取液,进样测定各成分的峰面积,计算平均回收率均大于95%,RSD均小于2%。
1.2.5 供试品含量测定
在1.2.1节色谱条件下,分别进样不同处理组样品制备所得的供试品溶液1 μL,测定峰面积,计算各处理组样品中的杠柳毒苷、杠柳苷元、杠柳次苷及4-甲氧基水杨醛成分含量。在该色谱条件下,各成分的色谱峰分离良好,部分结果见图1。
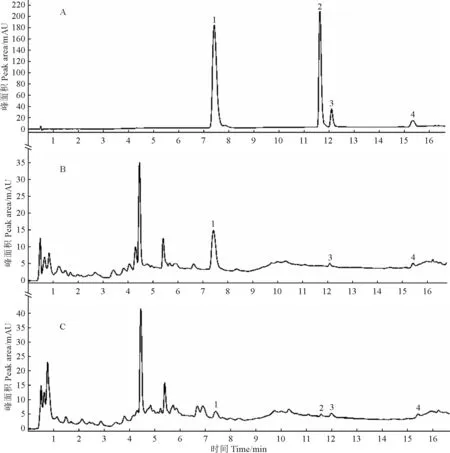
图1 混合对照品(A)、0.5%盐处理组根样品(B)、0.5%盐处理组茎样品(C)的UPLC色谱图1.4-甲氧基水杨醛;2.杠柳苷元;3.杠柳毒苷;4.杠柳次苷。Fig.1 UPLC chromatogram of mixed reference substances (A),root samples of 0.5% salt treatment group (B),and stem samples of 0.5% salt treatment group (C)1,4-methoxy salicylaldehyde.2,periplogenin.3,periplocin.4,periplocymarin.
1.3 数据处理
利用SPSS 25.0软件进行单因素方差分析,采用LSD 和Duncan多重比较法进行显著差异性检验,采用Origin 85.0软件作图。
2 结果与分析
2.1 盐胁迫对杠柳幼苗生长状态和生物量的影响
随着盐处理浓度的增加,杠柳幼苗的长势受到的抑制逐渐加重,但其在0.2%~0.5%盐胁迫浓度下长势良好(图2)。

图2 盐胁迫下杠柳幼苗叶片表型和根系形态变化A-E correspond to the control group and treatments with 0.2%,0.5%,1.0% and 2.0% salt concentrations,respectively.Fig.2 Changes of roots and leaves of P.s epium under salt stressA~E 分别对应对照组及0.2%、0.5%、1.0%、2.0%的盐浓度处理。
同时,从表2可知,杠柳幼苗地上部分干鲜质量在各浓度盐胁迫下均较对照组有所提高,并随着盐浓度增加而先升后降,且均在0.5% NaCl盐胁迫下增幅达到显著水平;杠柳幼苗根系鲜质量随盐胁迫浓度的增加呈现逐渐显著下降趋势,而根系干质量则先增加后降低,并在0.2%~0.5% 盐胁迫下显著增加;地上和地下干湿比在各浓度盐胁迫下均比明显增加,且有随盐浓度逐渐增加的趋势。可见,适宜浓度盐(0.5% NaCl)胁迫处理更有利于杠柳幼苗地上部和根系干物质的积累。

表2 不同浓度NaCl处理下杠柳幼苗生长变化Table 2 Growth of P.sepium seedlings under NaCl treatments
2.2 盐胁迫对杠柳幼苗根和茎中总甾体含量的影响
如图3所示,随着盐胁迫浓度的提高,杠柳幼苗根及茎中的总甾体含量均呈先升高后降低的趋势,且均在0.5% NaCl处理时达最大值。与对照相比,杠柳幼苗根及茎中总甾体含量在0.5% NaCl处理时分别显著提高了18.2%、36.2%;茎中总甾体含量在0.2% NaCl处理时比对照组间显著提高(P<0.05)。即适宜浓度盐胁迫有利于杠柳幼苗根及茎中的总甾体积累。
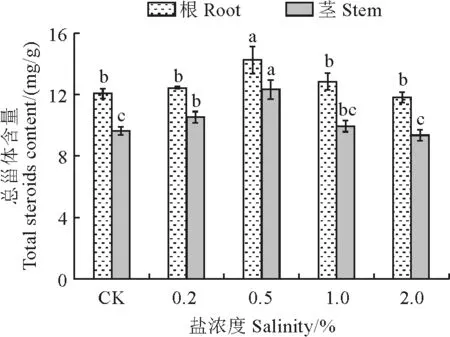
图3 盐胁迫下杠柳幼苗根和茎中总甾体含量变化不同小写字母表示处理组间在 5%水平差异显著。下同。Fig.3 Changes of total steroid content in roots and stems of P.sepium seedlings under salt stressDifferent lowercase letters indicate significant difference between the treatment groups at the 5% level.The same as below.
2.3 盐胁迫对杠柳幼苗根和茎中主要强心苷类成分含量的影响
3种强心苷类成分(杠柳毒苷、杠柳次苷和杠柳苷元)在杠柳幼苗茎中均有分布,且三者含量随着盐胁迫浓度的增加均呈先上升后下降的趋势(图4)。其中,杠柳毒苷与杠柳苷元含量在0.5% NaCl处理组下达到最大值,比相应对照组分别显著提高了35.1%、10.3%(P<0.05);杠柳次苷含量则在1.0%NaCl处理组下达到峰值,各盐浓度处理含量均显著高于对照组,分别达到对照组的1.28,3.97,5.49,2.51倍(P<0.05)。

图4 盐胁迫下杠柳幼苗根和茎中杠柳毒苷、杠柳次苷及杠柳苷元成分含量变化Fig.4 Changes of periplocin,periplocymarin,and periplogenin contents in roots and stems of P.sepium seedlings under salt stress
与杠柳幼苗茎中表现不同,在同样的检测条件下,杠柳根中未检测到杠柳苷元成分。杠柳幼苗根中杠柳毒苷、杠柳次苷含量随盐浓度的变化趋势均与茎中一致,但在盐胁迫前后均明显高于相应茎中含量;同时,与对照相比,各处理组根中杠柳毒苷和杠柳次苷含量均不同程度提高,且大多达到显著水平,杠柳毒苷含量分别达到对照组的1.32,2.08,1.70,1.38倍(P<0.05),杠柳次苷含量分别达到对照组的1.53,2.15,2.66,1.25倍。
2.4 盐胁迫对杠柳幼苗中4-甲氧基水杨醛含量的影响
图5显示,随着盐胁迫浓度的增加,杠柳幼苗根及茎中4-甲氧基水杨醛含量呈先升后降再升的趋势,并均在2.0%浓度下达到峰值,分别为0.841,0.323 mg/g,此时相较对照组分别显著提高了68.9%、317.6%。在不同盐浓度处理下,根及茎中的4-甲氧基水杨醛含量相比对照组均显著增加(P<0.05)。此外,在盐胁迫前后,杠柳幼苗根中4-甲氧基水杨醛含量均远远高于茎中含量。
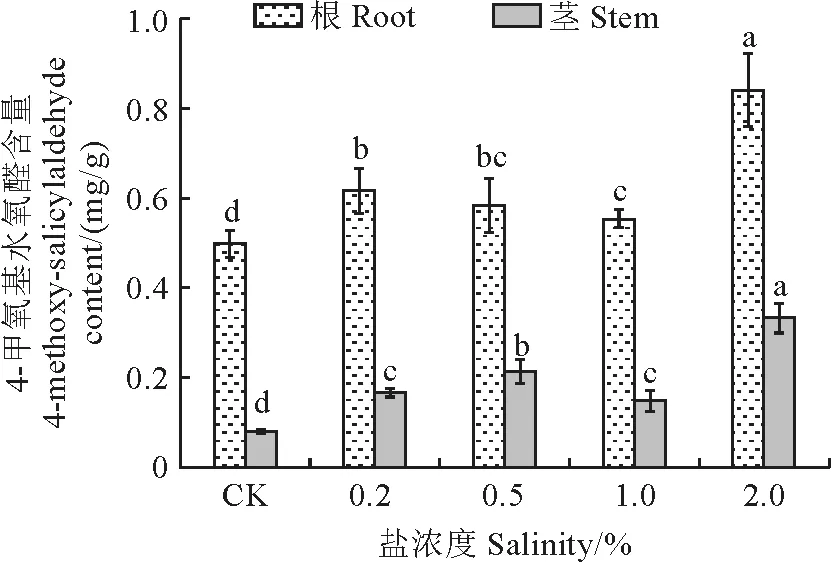
图5 盐胁迫下杠柳幼苗根和茎中4-甲氧基水杨醛含量变化Fig.5 Changes of 4-methoxy salicylaldehyde content in roots and stems of P.sepium seedlings under salt stress
3 讨论
在前期试验中[11]笔者发现杠柳耐盐性较强,表现在:(1)幼苗在中等盐(0.5%)胁迫强度下生长良好;(2)能够耐受1.0%的高强度盐胁迫,虽然高强度盐胁迫下根系生长受到一定程度的抑制,但其顶芽仍然能够萌发;(3)可以短期耐受2.0%的超高浓度盐胁迫,生长虽停滞但植株并未死亡。在本试验中,中等盐胁迫(0.5% NaCl)下杠柳幼苗地上干、鲜质量均显著高于其他处理组和对照组,说明中等盐胁迫更有利于杠柳地上部分生物量的积累。但在本试验中同时发现,杠柳幼苗根系发达程度及生物量鲜质量随盐胁迫强度增加呈现下降趋势,而干质量则在中低盐浓度下增加,且盐胁迫下根系的干湿比均明显高于对照组。这可能是因为盐胁迫导致植物生理干旱,耐盐植物通过盐离子区隔化和产生初生或次生代谢等渗透调节物质来调节细胞渗透势,重新建立细胞内稳态;其次有研究表明植物体在盐胁迫下,细胞壁通过木质化或木栓化起到细胞拒盐的作用,如有研究显示一些植物通过根部皮层或凯氏带细胞壁木化增厚达到拒盐目的[12],这在一定程度上增加了根系干质量和干湿比。张潭等[13]对枸杞幼苗的研究中也发现此现象。
次生代谢产物不是细胞正常生命活动所必需的物质,不直接参与植物生长和发育过程,但在植物提高自身保护和生存竞争能力、协调与环境关系上发挥着重要作用,且很多次生代谢产物也是药用植物的主要有效成分或活性成分[14]。相关试验表明,植物在逆境胁迫下可以通过次生代谢产物的积累来抵御受到的伤害,以保证植物的正常生长发育[15]。杠柳中的主要药效成分有C21甾体类及强心苷类等,两者均属于甾体类化合物。本试验对总甾体成分含量测定结果显示,盐胁迫处理对杠柳幼苗根及茎中的总甾体成分有明显诱导作用,在0.5%盐浓度下其含量相比对照组显著增加,这说明一定强度的盐胁迫有利于杠柳幼苗中甾体类成分的合成积累。
杠柳毒苷、杠柳次苷及杠柳苷元是存在于杠柳中的甲型强心苷类成分,既是属于总甾体中的重要成分,同时也是杠柳中的主要药效成分[16]。其中杠柳苷元在糖基转移酶的催化作用下,可与加拿大麻糖及葡萄糖相结合,从而形成糖苷类化合物杠柳次苷及杠柳毒苷。研究表明糖基转移酶可能广泛参与植物的各种胁迫应答,在植物的生长发育中发挥重要作用[17]。由糖基转移酶催化的糖基化反应,是植物细胞生长发育、代谢平衡过程中所必需的一种修饰反应,经修饰获得的糖基化产物具有不同的生物学功能,使得糖基化反应成为影响及调节植物生长代谢平衡的重要机制[18]。植物中的总甾体类成分包括植物甾醇、胆固醇和胆固醇代谢过程中的下游产物,如C21甾体和强心苷等。本试验中适当浓度的盐处理可以显著提高甾体类化合物的含量,同时也提高了甾体类下游代谢产物强心苷类成分含量。这说明适度的盐胁迫可以激活杠柳中甾体合成的整个代谢路径,参与逆境下的胁迫响应。但强心苷中的这3种主要成分被诱导出现的峰值所处盐胁迫浓度并不完全一致,这可能与不同盐胁迫下强心苷类产物合成下游路径中的糖基转移酶或水解酶活力变化有关。本试验还发现,杠柳不同部位中杠柳毒苷和杠柳次苷含量均表现为根中明显高于茎中,而杠柳苷元成分却未在根中检测到。在姚鹏等[10]研究结果中,同样显示其根中杠柳苷元含量很低或检测不到,这与本试验结果相一致。这可能是由于杠柳根中的糖基转移酶活性更强,从而导致其糖基化反应更迅速,导致苷元含量过低几乎检测不到;而在茎中杠柳苷元含量更高,则可能是茎中糖苷水解酶活力高或糖基转移酶活力低导致,这有待后续通过分子生物学方法进一步验证。
4-甲氧基水杨醛作为药典中规定的唯一指标性成分,其含量高低对于杠柳的药用质量评价具有重要作用。在植物中,4-甲氧基水杨醛合成的前体物质有4-羟基苯甲醛,两者结构中都具有酚羟基和醛基2种还原性基团,均属于多酚类次生代谢产物,能够中和逆境胁迫下植物体内产生的活性氧自由基,发挥抗氧化作用,减轻植物受到的氧化胁迫伤害[19]。本试验结果显示,杠柳幼苗根茎中4-甲氧基水杨醛含量随盐胁迫程度的增加呈先升后降而后再升的趋势,同时所有盐胁迫组中4-甲氧基水杨醛的含量均显著性高于对照组。由此可见,高盐强度(1.0% NaCl)胁迫虽然也能够显著促进植株中4-甲氧基水杨醛的诱导,但含量却低于中低强度盐胁迫组,这可能是由于高盐胁迫下植物体内氧化还原平衡被打破,产生了更多的活性氧自由基,而植物体产生的4-甲氧基水杨醛等多酚类物质被活性氧大量消耗,从而导致4-甲氧基水杨醛在杠柳地上和地下均呈现出下降的趋势。而超高强度盐(2.0% NaCl)胁迫下杠柳幼苗无论地上还是地下部分4-甲氧基水杨醛含量均显著增加(明显高于其他各组),这说明超高盐胁迫下,植物体通过次生代谢产生小分子还原性多酚类物质可能是应对极端恶劣条件下氧化胁迫的重要措施。前期试验表明杠柳幼苗中抗氧化酶体系发挥中和活性氧的作用是在中低浓度盐胁迫范围内,而黄酮类成分的含量则在高盐胁迫下显著提高,这进一步说明小分子还原性物质可能是杠柳幼苗在极端逆境条件下抵御氧化胁迫对其植株造成伤害的重要措施。卫乐等[20]研究也发现不同盐浓度下玫瑰中部分多酚物质含量呈先升后降而后再升的趋势,这与本试验结果具有一致性。此外,本试验中由于选择的研究对象为杠柳苗期(1个月龄),次生代谢产物积累时间较短,因此次生代谢产物含量均相对较低,但盐胁迫下次生代谢产物诱导的变化趋势显著,说明适度的盐胁迫可以促进杠柳植物体中次生代谢产物的积累。
4 结论
在中等浓度(0.5%)盐胁迫处理下,杠柳幼苗地上部分的生物量积累显著增加,同时根系干质量也显著增加;杠柳幼苗根及茎中的主要次生代谢产物如总甾体、强心苷类等含量在0.5%盐胁迫浓度作用下均明显上升,而4-甲氧基水杨醛含量则在2.0%的高浓度盐胁迫下显著增加。这为杠柳在中等强度盐渍化土壤上引种栽培提供了理论依据,对盐渍化土壤改良利用及杠柳资源开发提供了新思路,同时也为杠柳道地药材产地的划分提供了参考。
