影响月季花瓣呈色的理化因子及花色苷组分分析
2024-03-08袁美静康晓玲丁传雨
袁美静,马 誉,巫 瑞,康晓玲,丁传雨,杜 丽,3*
(1 南阳师范学院 生命科学与农业工程学院,河南南阳 473061;2 南阳师范学院 河南省月季花种质创新与栽培技术工程研究中心,河南南阳 473061;3 南阳师范学院分析测试中心,河南南阳 473061)
月季(Rosahybrida)是蔷薇科蔷薇属多年生直立或攀援木本植物[1]。作为中国的传统十大切花之一,月季具有极高的经济价值,其丰富多彩的花色、馥郁芬芳的香味、易于繁殖的特性使得月季在园林绿化和切花、盆花市场中占据重要地位[2]。花朵颜色是月季最重要的观赏特性[3],直接影响月季的商业价值。市场上的月季品种繁多,花色丰富,但由于月季自身缺乏能够编码合成蓝色色素的基因,导致缺少色泽纯正的蓝色系月季。因此,培育蓝色、复色等新颖花色的月季品种是当前育种工作者的主要目标之一。明确月季花色的形成机理是进行花色育种工作的前提。通过对影响月季花瓣呈色的各理化因子的定量评价及其相关性分析,对于探究花色形成机理和种质创新以及丰富月季市场具有重要意义。
前人研究发现,月季花色的形成具有复杂性,不仅受温度、光照、水分等外部因素影响,还受花瓣表皮细胞结构[4]、细胞液pH 值[5-6]、金属离子[7-8]、色素成分[9-10]等内部因素的调控,其中,花瓣中色素的种类和含量直接决定其花瓣颜色[11]。经研究发现,月季花瓣中主要含有的色素种类为类黄酮、类胡萝卜素以及少量的叶绿素[12]。其中,类黄酮是月季中主要的色素类群[13],主要包括花色苷和黄酮醇苷。花色苷则是由花青素通过与酰基、甲基或糖基的共价连接而形成更加稳定的结构,主要决定花瓣颜色[13-15];黄酮醇苷则是一类辅助色素,使花瓣颜色更加稳定[16]。已有研究表明,不同花色月季的形成受色素种类和含量的重大影响,Wan等[17]研究20个月季品种,并发现粉红色花瓣主要由矢车菊素/天竺葵素-3,5-葡萄糖苷和矢车菊素/天竺葵素-3-O-葡萄糖苷等花色苷赋予,黄色花瓣主要由类胡萝卜素赋予,橙色是由花色苷和类胡萝卜素的组合赋予。Yeon等[18]研究不同花色月季花瓣所含物质,其中粉色和红色花瓣以花色苷为主,黄色花瓣中含有以β-胡萝卜素为主,白色花瓣含有极少量类胡萝卜素。由此可见,不同花色苷的种类和含量使月季呈现出红、粉、紫等多种花色[19]。目前,对于花色苷的分析鉴定主要采用超高效液相色谱-飞行时间串联质谱(UPLC-TOF-MS/MS)技术,由于其能够提供精确分子数、对未知物质进行分析鉴定等优点,在花色苷的分析鉴定方面得到广泛应用。王琳等[20]在对葡萄风信子花色苷的研究中,建立了适用于葡萄风信子的花青素苷分析方法,利用HPLC-MS技术鉴定出6种花色苷。Li等[21]运用UPLC-TOF-MS/MS对15个水仙品种花瓣进行分析,鉴定出17种黄酮醇,16种类胡萝卜素,其中绿原酸首次在水仙中被鉴定,为水仙的花色育种奠定了物质基础。
目前,国内外关于月季花色的研究也有涉及,但多数是对于某一个品种或某一特定类群的研究[22-23],缺乏对多个色系月季花瓣及不同色系月季花瓣呈色机理的系统解析,对后续进行花色育种工作的指导价值不高。本研究选取8个不同花色的月季品种为试验材料,分别测定其花瓣颜色参数、花瓣细胞结构、细胞液pH 值、金属离子含量、色素含量等多个理化指标,并利用UPLC-TOF-MS/MS技术对其花色苷成分进行定量分析。随后将花瓣颜色参数与内部影响因子及花色苷组分进行相关性分析,明确造成不同花色月季形成的主要影响因子和花色苷成分,揭示花色形成机理,筛选花色改良受体品种,为后续利用分子育种手段丰富月季花色,培育符合消费者审美的月季花色新品种提供理论基础。
1 材料和方法
1.1 植物材料和取样
选取河南省月季种质创新与栽培技术工程研究中心种植的8个不同花色的月季品种为供试材料,分别为白色品种‘藤冰山’、黄色品种‘朝圣者’、橙色品种‘蜂蜜焦糖’、粉色品种‘瑞典女王’、粉紫色品种‘薰衣草花环’和‘蓝色风暴’、红色品种‘嫦娥奔月’、紫红色品种‘雀之舞’。2022年4—5月收集盛花期(花萼、花瓣完全展开)花瓣,每个品种选择长势一致的植株,分散选取中层花瓣,部分新鲜样品立即用于花色表型测定和解剖结构观测,其余花瓣冻存于-80 ℃冰箱用于后续实验。
1.2 花瓣颜色测定
取盛花期生长良好的中层花瓣,在散射光源下根据国际照明委员会的CIEL*a*b*色空间系统,使用分光色差仪(CS-422,中国杭州彩谱公司)测定各个品种的花色参数,测定时将分光色差仪的集光口对准花瓣中央着色部位,获得花色的明度值L*和色相值a*、b*,并运用公式C*=(a*2+b*2)/2计算彩度C*。每个品种测量3 朵花,每朵花取5片花瓣,重复测定3次,取平均值。
1.3 花瓣形态结构观察
取盛花期花瓣中间部位,采用徒手切片法制作花瓣横切面、上下表皮的临时水装片,放置于光学显微镜(RX 50,舜宇光学)下进行观察,同时使用软件进行拍照,观察色素分布及表皮细胞形态,使用Image J软件测量花瓣横切面厚度及上表皮细胞面积,每个品种至少拍3张,每张照片至少测定5个细胞,取平均值。
1.4 花瓣pH 测定
参考唐前瑞[24]的方法并稍作修改,取盛花期新鲜花瓣0.5 g并加2 mL 双蒸水(不加石英砂)研磨成匀浆,置于离心管中,用平面笔式pH 计(pHS-3C,中国上海仪电科学仪器公司)测量花瓣匀浆pH值,每个品种生物学重复3次,技术重复5次,取平均值。
1.5 花瓣金属离子含量测定
各月季品种取适量花瓣,去离子水清洗3次后置于电热恒温鼓风干燥箱100 ℃杀青30 min,再置于70 ℃的温度环境中烘干10 h。称取0.5 g烘干样品,加入71%硝酸5 mL,于聚四氟乙烯消解罐中消解,后赶酸至近干,采用1%硝酸溶液稀释并定容。试样经消解后,由电感耦合等离子体质谱仪(ICAP RQ,赛默飞)测定,以元素特定质量数(质荷比,m/z)定性,采用外标法,测定各品种月季花瓣中钙(Ca2+)、镁(Mg2+)、铁(Fe3+)、锰(Mn2+)、铝(Al3+)5种金属离子含量,每个品种进行3次生物学重复,取平均值。
1.6 总黄酮含量测定
精确称取芦丁标准品,将其配制成1 mg/L 母液,分别取芦丁0,0.25,0.5,0.75,1.0,1.25 mg/mL置于25 mL 容量瓶中,采用NaNO2-Al(NO3)3法显色。首先加入5% NaNO2溶液1 mL,摇匀后放置6 min,再加入10% Al(NO3)3溶液1 mL,摇匀后放置6 min,加入4% NaOH 溶液10 mL,用70%乙醇定容至刻度,放置10 min。显色反应后,在510 nm 处测定不同浓度芦丁的吸光值A,以芦丁实际浓度为横坐标,以吸光值A为纵坐标绘制标准曲线得到线性回归方程为y=0.5148x+0.0062,R2=0.999 5。
分别取8个品种烘干的花瓣粉末0.1 g,加入70%乙醇10 mL,超声清洗仪100 W 提取1 h,取提取后的溶液2 mL,按照上述NaNO2-Al(NO3)3法进行显色反应,显色反应后,在510 nm 处测定不同品种月季花瓣供试溶液的吸光值A。不同品种月季花瓣的总黄酮含量按照上官琨瑶等[25]的方法计算,其中供试溶液的浓度按照芦丁标准曲线的回归方程计算。每个品种进行生物学重复3次,取平均值。
1.7 总花色苷含量测定
总花色苷含量测定采用pH 示差法[26]。取0.2 g各品种新鲜花瓣,剪碎后置于具塞试管中,向试管中加入1%盐酸甲醇溶液(1∶99,V/V)10 mL,盖上塞子超声清洗仪100 W 提取20 min,直至组织样品变白。取过滤后的各品种月季花瓣花色苷提取液1 mL,用pH 分别为1.0和4.5的缓冲液稀释定容至3 mL,平衡80 min后分别测定530 nm 及700 nm处的吸光值,每个品种3次生物学重复,取平均值。以1%盐酸甲醇溶液为空白对照,以矢车菊素-3-O-葡萄糖苷作为花色苷标准品,花色苷总量计算公式为:
式中:TA为花色苷总量;A为2个pH 下吸光度的差值;MW为花色苷分子量(以矢车菊-3-O-葡萄糖苷计,449.2 g/mol);D为稀释倍数;e为摩尔消光系数[以矢车菊-3-O-葡萄糖苷计,29 600 L/(mol·cm)];L为光程(1 cm);G为花瓣鲜重(g);V为提取液体积(mL)。
1.8 总叶绿素及类胡萝卜素含量测定
取各品种新鲜花瓣0.2 g分别放入研钵中,加少量石英砂和碳酸钙粉以及10 mL无水乙醇溶液,研磨至组织变白,常温避光环境静置提取。将研磨的匀浆转移至离心管中,冷冻离心机13 000g离心10 min,取上层提取液,采用紫外-可见分光光度计测定665 nm、649 nm 及470 nm 处的吸光值,以无水乙醇溶液为空白对照,每个品种进行生物学重复3次,取平均值。
总叶绿素及类胡萝卜素含量计算公式[27]分别为:
式中:Ca为叶绿素a浓度;Cb为叶绿素b浓度;Ca+Cb为总叶绿素浓度;Cxc为类胡萝卜素浓度(mg/L)。
花瓣色素含量计算公式为:
式中:T为新鲜花瓣样品中花色素的含量(mg/g);C为色素浓度(mg/L);V为提取液体积(mL);N为稀释倍数;W为样品鲜重(g)。
1.9 月季花瓣中花色素成分的UPLC-TOF-MS/MS分析
取各品种月季花瓣,液氮速冻后快速研磨,精密称取0.5 g样品,以先前处理优化后的条件为基础,料液比1∶10,提取液为甲醇∶水∶甲酸=70∶29∶1(V/V/V),提取方式为冰浴超声提取30 min进行样品前处理,后将提取液转移至离心管中13 000g离心15 min,保留上清液用0.22 μm 有机滤膜过滤后待上机测定用。
色谱条件:Acquity UPLC BEH C18 色谱柱(100 mm×2.1 mm,1.7 μm);柱温30℃;进样量2 μL;流动相A 为0.1%甲酸水溶液,流动相B 为0.1%甲酸乙腈溶液,流速为0.4 mL/min。液相色谱梯度洗脱条件为0~8 min,95% A,5% B;8~13 min,86%~76% A,5%~14% B;13~16 min,76%~64% A,24%~36% B;16~17 min,64%~50%A,36%~50% B;17~20 min,50%~50% A,50%~50% B。质谱条件:电喷雾离子源;MSE 正负离子模式,质量扫描范围50~1 200 m/z;毛细管电压3 000 V;锥孔电压45 V,提取锥电压40 V;离子源温度100 ℃,脱溶剂气温度450 ℃;锥孔气流量50 L/h,碰撞能6~40 V。
1.10 数据分析
上述试验均生物学重复3 次,用Excel 2021、SPSS 22.0、Origin 2021软件进行数据处理与分析。用Masslynx V4.2 软件对花色苷组分分析,用UNIFI软件结合自建数据库鉴定花色苷化合物结构。
2 结果与分析
2.1 花色表型特征
为更好地了解不同花色月季花瓣表型特征的差异,选择不同色系的8种月季(图1),其花色表型的测定数据见表1。在CIEL*a*b*系统中,L*值代表花瓣明暗变化程度,与花色发育呈负相关,白色品种‘藤冰山’(A)的L*值最高,花色最明亮;紫红色品种‘雀之舞’(H)的L*值最低,花色较暗,且随花色逐渐变深,L*值逐渐减小。a*值代表花瓣红、绿变化程度,红色品种‘嫦娥奔月’(G)的a*值最高;白色品种‘藤冰山’(A)的a*值较低。随着花瓣红色加深,a*逐渐增高。b*值代表花瓣黄、蓝变化程度,黄色品种‘朝圣者’(B)及橙色品种‘蜂蜜焦糖’(C)的b*值显著高于其余品种,粉紫色系品种‘薰衣草花环’(E)‘蓝色风暴’(F)的b*值显著低于其余品种,且为负值。表明b*值从正值到负值的变化,代表黄色减弱蓝色增强。C*值代表花瓣色彩鲜艳程度,白色品种‘藤冰山’(A)C*值最低,红色品种‘嫦娥奔月’(G)的C*值最高。C*值在8个月季品种中的变化趋势与a*值相似,与L*值变化趋势相反。

表1 8个月季品种的花色表型色差值Table 1 Color difference values of flower color phenotypes of 8 rose cultivars

图1 8个月季品种的花色表型(Bar=1 cm)A.藤冰山;B.朝圣者;C.蜂蜜焦糖;D.瑞典女王;E.薰衣草花环;F.蓝色风暴;G.嫦娥奔月;H.雀之舞。下同。Fig.1 Flower color phenotypes of 8 rose cultivars (Bar=1 cm)A.Climbing Iceberg.B.The Pilgrim.C.Honey Caramel.D.Queen of Sweden.E.Lavender Flower Circus.F.Shinoburedo.G.Chang'e Benyue.H.Dance of Peacock.The same as below.
2.2 花瓣解剖结构特征及色素分布
通过徒手切片法观察月季花瓣组织结构,其花瓣横截面见图2,花瓣上表皮细胞见图3。月季花瓣内部结构主要分为上表皮、栅栏组织、海绵组织和下表皮。在8个月季品种中,上表皮细胞均为锥形突起、排列紧密,能够吸收更多光线,加深花瓣颜色;下表皮细胞多为扁平状,能够反射光线,8个品种间具有相似的光吸收特征。

图2 8个月季品种花瓣的横切面结构(Bar=100 μm)Fig.2 Cross-sectional structure of petals of 8 rose varieties(Bar=100 μm)

图3 8个月季品种花瓣的上表皮结构(Bar=50 μm)Fig.3 Upper epidermal structure of petals of 8 rose varieties(Bar=50 μm)
但不同颜色的品种在色素分布等方面具有很大差异,在白色品种‘藤冰山’(A)中,各组织结构均未观察到色素分布,结合前人研究结果,认为白色花色是由于细胞间隙的微小气泡在光的照射下发生连续反射形成的一种光学颜色,受花色素的影响较小[9,28]。
在其余品种中,色素主要分布在上、下表皮。表皮细胞颜色与花色一致,说明花瓣结构与花色的形成紧密相关。不同月季品种盛花期花瓣厚度及上表皮细胞面积如图4所示,8个月季品种的花瓣厚度在185.52~256.15 μm,不同花色月季花瓣厚度之间存在显著差异。其中,‘藤冰山’、‘朝圣者’、‘蜂蜜焦糖’和‘嫦娥奔月’花瓣较厚,显著大于其他品种,粉紫色品种‘薰衣草花环’花瓣最薄。

图4 8个月季品种花瓣的横切面厚度及上表皮面积Fig.4 Transverse section thickness and upper epidermis area of petals of 8 rose cultivars
不同月季品种盛花期花瓣上表皮面积也有显著差异,其中‘瑞典女王’上表皮面积最大,‘雀之舞’上表皮面积最小。综合比较,浅色系月季上表皮面积较大,深色系较小。
2.3 细胞液pH 值测定
8个月季品种的细胞液pH 值测定结果为图5所示,细胞液pH 值范围为4.19~5.76,偏弱酸性,不同花色品种之间存在显著差异。其中,粉紫色品种‘蓝色风暴’细胞液pH 值最高(5.76),‘薰衣草花环’次之(5.75),红色品种‘嫦娥奔月’最低(4.19)。由于白色品种含有少量或不含色素,黄色品种多含有类胡萝卜素,而橙红色品种相较于黄色品种还通常含有胡萝卜素及花色苷两类色素,因此‘蜂蜜焦糖’相较于‘朝圣者’及‘藤冰山’pH 值较低。
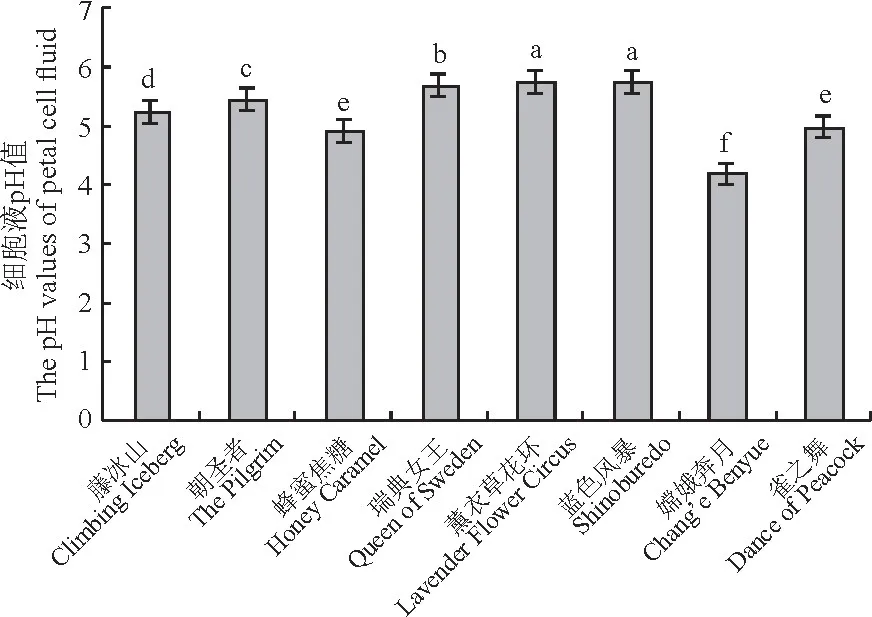
图5 8个月季品种花瓣细胞液pH 值Fig.5 pH values of petal cell fluid of 8 rose cultivars
综上所述,花色偏蓝紫色pH 值较高,花色偏红色pH 值较低,在8个月季品种中,由粉紫色到紫红色,pH 值逐渐降低。
2.4 花瓣金属离子含量测定
对8 个月季品种中Mg2+、Fe3+、Ca2+、Al3+、Mn2+含量进行测定(表2)。在测定的5种金属元素中,含量最高的为Mg2+,其次为Ca2+,Fe3+、Al3+和Mn2+在月季中仅微量存在。Mg2+含量在0.592~0.968 g/kg之间,其中在橙色品种‘蜂蜜焦糖’中含量最高,粉紫色品种‘薰衣草花环’‘蓝色风暴’含量较低。Ca2+的含量为0.552~1.240 g/kg,在紫红色品种‘雀之舞’中的含量最高,白色品种‘藤冰山’的含量最低。
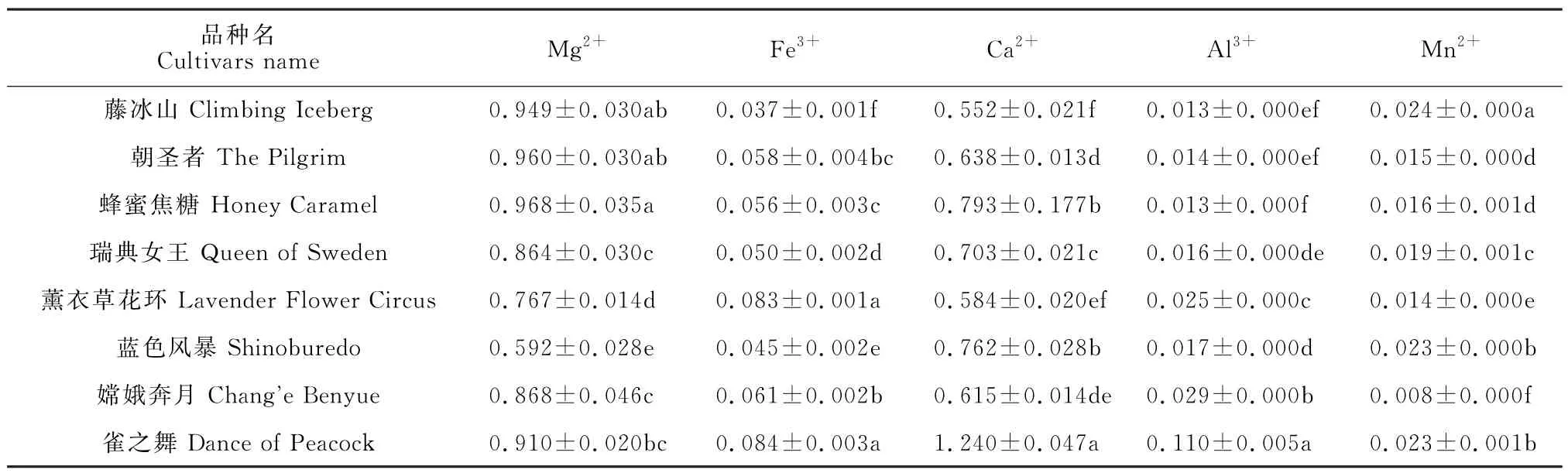
表2 8个月季品种的金属离子含量Table 2 Metal ion content of 8 rose cultivars g/kg
在Fe3+、Al3+、Mn2+3 种离子中,Fe3+的含量较高于其他2种,在0.037~0.842 g/kg之间,其中紫红色品种‘雀之舞’的含量最高,整体趋势随着花色变浅含量降低。Al3+含量最高的仍为‘雀之舞’,含量是其余品种的4~6倍;Mn2+在所有月季花瓣中含量均较低。总之,除了Mg2+外,花瓣颜色最深‘雀之舞’所含有的金属离子含量均显著高于其余品种。
2.5 花瓣色素含量测定
色素是影响花色的主要因素,包括总黄酮、总花色苷、总叶绿素及类胡萝卜素。通过对月季花瓣中色素含量的测定,结果(表3)表明,8个月季品种中均含有黄酮、花色苷以及叶绿素这三类色素,类胡萝卜素仅存在于黄色品种中,且同类色素在不同品种间的含量存在显著差异。总黄酮含量在各个品种月季花瓣中的占比最高,其中紫红色品种‘雀之舞’的总黄酮含量显著高于其余品种,黄色品种‘朝圣者’的总黄酮含量最低。其余品种随着花色逐渐变浅总黄酮含量逐渐降低,且存在显著差异。花色苷是决定花朵颜色的主要色素之一。
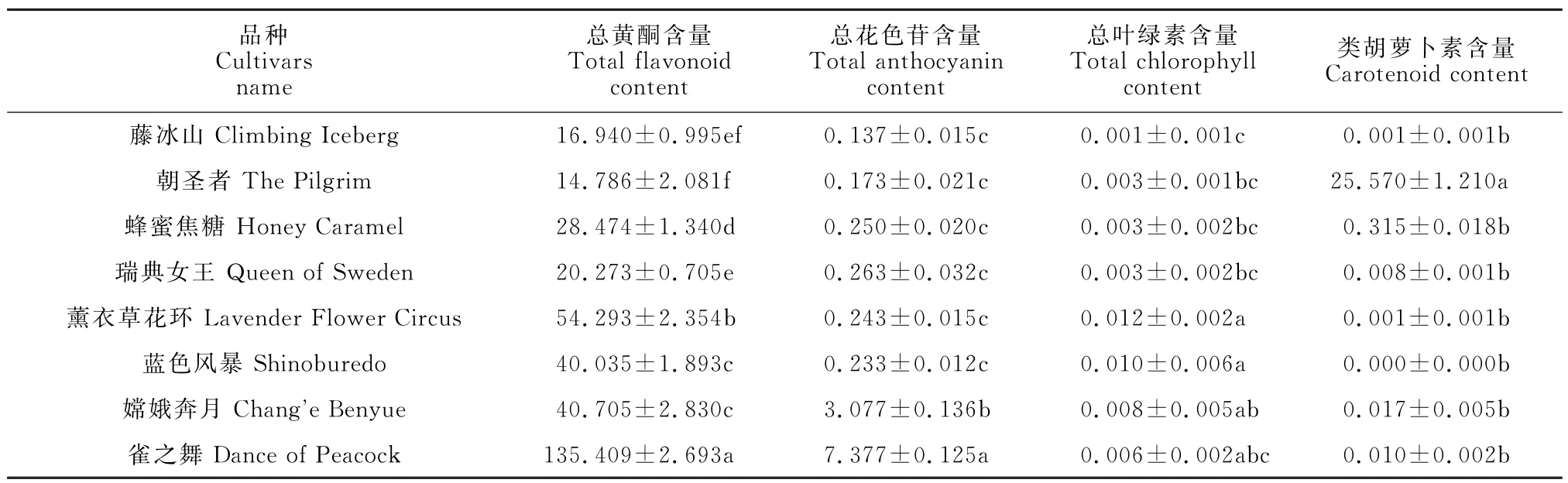
表3 8个月季品种的花瓣色素含量Table 3 Petal pigment content of 8 rose cultivars mg/g
‘雀之舞’的总花色苷含量最高,其次为‘嫦娥奔月’,两者的花色苷含量均与其余品种间存在显著差异,而白色品种‘藤冰山’的总花色苷含量最低。总黄酮与总花色苷的变化趋势一致,都是随着花色变浅含量降低。由于8 个月季品种中没有绿色系花朵,因此8个月季品种中的总叶绿素含量都微量存在。类胡萝卜素仅在黄色品种‘朝圣者’及橙色品种‘蜂蜜焦糖’中存在,且‘朝圣者’的类胡萝卜素含量远高于其余品种,但其他3 种色素的含量却较低。这说明,色素的种类和含量直接决定花瓣颜色,总花色苷和总类黄酮含量变化趋势一致,随着花色变浅含量降低,控制着粉、红、紫色花的形成。类胡萝卜素调控黄色花的形成,并与花色苷共同调控橙色花的形成。
2.6 花色表型测定值与不同呈色理化因子的相关性分析
为进一步明确影响月季花瓣呈色的主要因子,将测定的各理化指标与花瓣表型参数进行相关性分析,使用Pearson相关系数表示相关关系的强弱情况(表4)。经分析可知,花瓣颜色与金属离子含量和色素含量之间均存在一定的相关性。L*与a*、Fe3+、Al3+、总黄酮和总花色苷含量之间均存在显著负相关关系,其中L*与a*、总黄酮含量、总花色苷含量之间存在极显著负相关(P<0.01)。a*与pH 值及总花色苷含量之间存在显著相关关系,其相关系数分别为-0.787和0.787。Fe3+与总黄酮含量之间存在显著正相关(P<0.05);Ca2+与Al3+、总黄酮含量之间为极显著正相关(P<0.01),与总花色苷含量之间存在显著正相关;Al3+含量与总黄酮、总花色苷含量均存在极显著正相关。4种色素中,总黄酮与总花色苷之间呈现极显著正相关关系,类胡萝卜素与其余各指标间均不存在相关关系,总叶绿素与除Mg2+外的其他各指标之间均不存在相关关系。
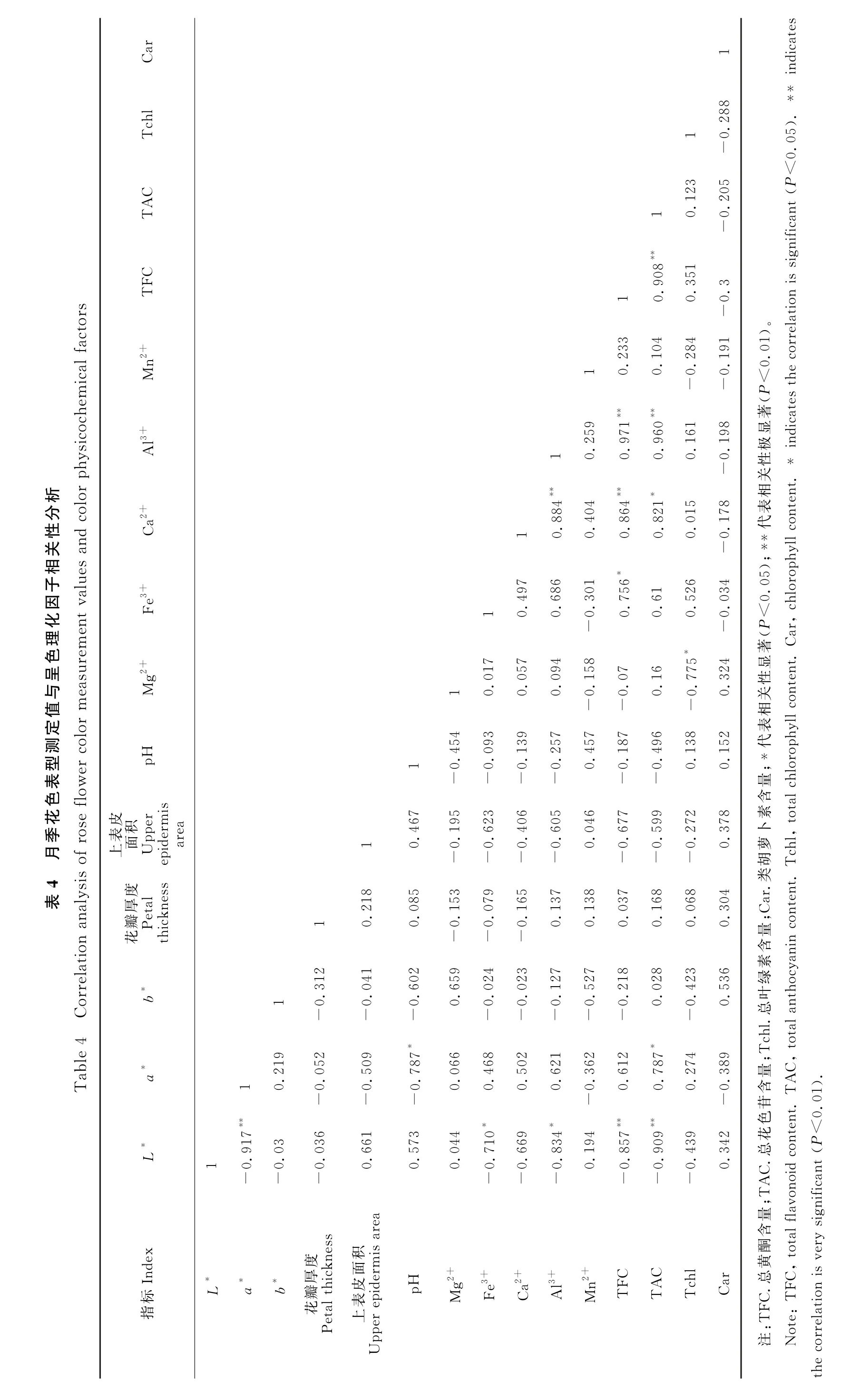
综上所述,降低花瓣中总黄酮、总花色苷、Fe3+、Al3+含量可增加花瓣亮度,反之花瓣颜色变暗加深;降低花瓣细胞液pH 值或增加花瓣中花色苷含量可提高月季红色程度,而随着月季花瓣红色程度增加,花瓣亮度降低;Fe3+、Ca2+和Al3+含量都随着总黄酮含量的升高而升高,Al3+含量还与总花色苷含量极显著正相关。4种色素中,总黄酮含量与总花色苷含量相辅相成,主要决定花朵颜色,随着总黄酮含量增加,总花色苷含量也呈递增趋势;而总叶绿素及类胡萝卜素与其余指标间仅呈现弱相关性,未达显著水平。因此,不同花色月季呈色的理化因子之间关系复杂,相互影响,共同作用决定花瓣颜色,其中总花色苷含量和总黄酮含量占主要地位,直接影响花色呈现,金属离子及pH 值等因素通过改变花色素结构来改变花色。
2.7 月季花瓣中花色苷组分与花色表型相关性分析
本团队在前期工作[29]中已建立稳定的月季花色苷组分定量检测的方法,并对本试验中的8个品种月季的花色苷组分含量进行精准测定,其结果见表5。由此可知,不同花色月季花瓣中花色苷及黄酮类物质含量存在显著差异,花色由紫红色到白色,矢车菊类色素含量逐渐降低,其中矢车菊类色素在紫红色品种‘雀之舞’中含量最高,黄色品种‘朝圣者’含量最低,且矢车菊-3,5-葡萄糖苷的含量高于矢车菊-3-O-葡萄糖苷含量;天竺葵类色素在红色品种‘嫦娥奔月’和橙黄色品种‘蜂蜜焦糖’中含量较高,在其余品种中仅微量存在;二氢杨梅素及飞燕草-3-O-葡萄糖苷在8个品种月季中仅微量存在,而芦丁的含量在粉紫色系品种中含量相对较高,其中‘薰衣草花环’的含量最高。在前期工作中,明确了8个月季品种中花瓣的花色苷及黄酮类物质的含量,但对于月季花瓣颜色的形成是否与花色苷及黄酮存在一定关系及与哪类花色苷的关系最为密切尚不清楚。故本文引用团队前期测定的结果,与上文测定的花瓣颜色参数及总色素含量等指标进行相关性分析,进一步明确造成月季花色差异的小分子代谢物,明确月季花瓣呈色机理,同时也为后期利用分子手段进行花色育种奠定基础。

通过计算月季花瓣颜色参数与花色苷含量、黄酮含量之间的Person相关系数,对其相关性进行分析,并以热图形式显示(图6)。由此可知,不同颜色参数与花色素之间存在很强的相关性。L*与a*、总黄酮含量及总花色苷含量之间呈极显著负相关(P<0.01),并与单个花色苷含量之间也存在相关性,与单个黄酮类化合物间无相关性。L*与矢车菊-3,5-葡萄糖苷和矢车菊-3-O-葡萄糖苷的相关系数分别为-0.97和-0.79,相比之下,L*与矢车菊-3,5-葡萄糖苷的相关性较矢车菊-3-O-葡萄糖苷强。

图6 月季花瓣颜色测定值与花色苷成分的相关性分析Cy3D5G,cyanidin-3,5-glucoside.Pe3D5G,pelargonidin-3,5-diglucoside.Cy3G,cyanidin-3-O-glucoside.Pe3G,pelargonidin-3-O-glucoside.De3G,delphinidin-3-O-glucoside.* The correlation is significant (P<0.05);** the correlation is very significant (P<0.01).Fig.6 Correlation analysis of rose flower color measurement values and anthocyanin componentsCy3D5G.矢车菊素-3,5-葡萄糖苷;Pe3D5G.天竺葵素-3,5-葡萄糖苷;Cy3G.矢车菊素-3-O-葡萄糖苷;Pe3G.天竺葵素-3-O-葡萄糖苷;De3G.飞燕草素-3-O-葡萄糖苷;Ampelopsin.二氢杨梅素;Rutin.芦丁。*代表相关性显著(P<0.05);**代表相关性极显著(P<0.01)
而a*恰好相反,a*与大部分单个花色苷含量间存在显著正相关,其中与矢车菊-3,5-葡萄糖苷的相关性最强。而b*与各指标间均不存在相关性。总花色苷含量与3种花色苷含量呈显著正相关性,其中与矢车菊-3,5-葡萄糖苷呈极显著正相关(P<0.01),与天竺葵素-3,5-葡萄糖苷、天竺葵-3-O-葡萄糖苷间均存在显著正相关性(P<0.05)。飞燕草-3-O-葡萄糖苷只与二氢杨梅素之间存在显著正相关关系,飞燕草-3-O-葡萄糖苷的含量随着二氢杨梅素的变化而变化。此结果揭示了月季花瓣花色苷和黄酮代谢之间的相关性。矢车菊类色素之间和天竺葵类色素之间均存在显著正相关性,但两类色素之间无相关性,且矢车菊素、天竺葵类色素和飞燕草类色素之间均不存在相关关系,且芦丁与花瓣表型及花色苷均不存在相关性。这说明三类色素在花色苷合成通路中处于不同的合成方向,调控不同花色的形成,而芦丁不是影响花瓣颜色变化的主要因子。
综上所述,花色苷是影响月季呈色的主要因子,芦丁等黄酮类物质辅助月季呈色。矢车菊-3,5-葡萄糖苷在月季花色苷组分中占据主体地位,是影响月季花瓣亮度L*和红度a*的主要因素,其中对L*产生负影响,对a*产生正影响。在8个品种月季中,除橙黄色品种‘蜂蜜焦糖’外,矢车菊-3,5-葡萄糖苷在5种花色苷组分中占比最高,且在紫红色品种中含量最高。由此发现,矢车菊-3,5-葡萄糖苷能够调控月季花瓣向紫红色发展。天竺葵类色素与L*、总花色苷含量间也存在显著正相关性。天竺葵-3,5-葡萄糖苷占比高于天竺-3-O-葡萄糖苷,且在红色品种‘嫦娥奔月’中最高,其次是橙黄色品种‘蜂蜜焦糖’,其显著高于其他品种。这说明天竺葵色-3,5-葡萄糖苷对于月季花瓣红色花朵的形成有一定的影响,并与类胡萝卜素一起调控月季形成橙红色的花朵。
飞燕草-3-O-葡萄糖苷在月季中仅微量存在,但在粉紫色系品种 ‘薰衣草花环’中发现极高的芦丁含量。这说明月季中极少量的飞燕草素不足以使花朵变蓝,但无色的黄酮醇可能与花色苷发生作用,从而使月季呈现出一定程度的蓝紫色。
2.8 8个品种月季花瓣花色素组分的聚类分析
根据所测得的花色苷和黄酮类物质的含量,对8个品种月季进行聚类分析。由图7 可知,当欧式距离在400时,将所有样本聚类为3类,即白色品种‘藤冰山’、粉色品种‘瑞典女王’及紫红色品种‘雀之舞’为一类;黄色品种‘朝圣者’、橙黄色品种‘蜂蜜焦糖’、粉紫色品种‘蓝色风暴’及红色品种‘嫦娥奔月’为一类;粉紫色品种‘薰衣草花环’单独为一类;且还可发现‘薰衣草花环’始终单独为一类。
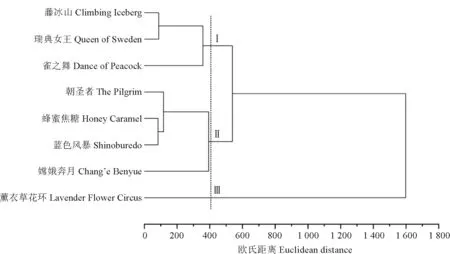
图7 8个品种月季花瓣花色素成分的聚类分析Fig.7 Cluster analysis of 8 cultivars of rose based on anthocyanin components
通过对比各品种月季花色苷组分的含量,发现第一类中3个品种的月季所含的芦丁含量在7个组分中占比最高,而‘藤冰山’和‘瑞典女王’不含或含量较少花色苷组分,但‘雀之舞’中矢车菊-3,5-葡萄糖苷的含量较高,因此当欧式距离为88时,其又与‘雀之舞’区分开来。第二类中4个品种中均含有较丰富的矢车菊-3,5-葡萄糖苷和少量的芦丁,故聚为一类;但与其他3个品种相比,‘嫦娥奔月’中还含有较高的天竺葵-3,5-葡萄糖苷,因此在欧氏距离为118时其也与其他3 个品种区分开来。第三类中‘薰衣草花环’以极高的芦丁含量和较低的花色苷含量显著区别于其他品种,单独成类。
3 讨论
3.1 总花色苷和总黄酮含量是影响月季花瓣呈色的主要因子
花色是植物最重要的观赏性状之一,受多种呈色因子影响,对各因子进行量化评价并分析其相关性对于探究花色形成机理及种质创新具有重要意义。在对8个不同花色月季品种的花色表型进行测定时,随着花色加深,L*值逐渐减小,a*值、C*值呈现增大的趋势,L*值与a*值、C*均呈负相关,L*值与b*值之间无显著相关性这与刘艺平等[30]对荷花花色表型和李盈侠等[31]对牡丹花色表型研究的结果一致。花瓣亮度L*值及花瓣彩度C*值均与花瓣红度a*值相关,相较于b*值,a*值对于花瓣表型的参考价值更大,这与吴艳梅等[32]对丽格海棠花色表型研究的结果一致。花瓣结构主要是影响光线反射从而影响花色。研究结果表明,白色花瓣中无色素分布,其余品种色素在花瓣上下表皮中均有分布。与程怡等[22]对月季‘仙境’的研究结果一致。月季与芍药[33]、葡萄风信子[34]的色素分布模式一致,与长春花[27]等不同。而花瓣厚度不是决定花朵着色的主要因素,与刘金芝[35]的研究结果相同。细胞液的酸碱程度影响着花青素的结构稳定和存在方式,在观赏植物花瓣中,细胞液大多偏酸性,本研究中的8个月季品种pH 值为4.19~5.76,偏弱酸性,蓝紫色系花瓣pH 值最高;随着花瓣颜色逐渐加深,pH 值逐渐降低,且pH 值与红度a*值呈显著负相关,可通过降低pH 值增加花瓣红度,与余智城等[36]对野牡丹花色的研究结果一致。研究金属离子对花瓣呈色的作用,发现Fe3+、Ca2+、Al3+与花瓣表型颜色均具有相关性,在一定程度上影响着月季花瓣呈色,且Al3+与总花色苷之间呈极显著相关,因此推测Al3+可能会与花青素苷β环上的二羟基结合形成高度着色的络合物[37],从而影响花朵呈色。唐东芹等[38]在对风信子花色进行研究时也证实了这一点。相关性研究显示,尽管pH 值、Fe3+、Ca2+、Al3+、总黄酮含量、总花色苷含量与月季花瓣呈色密切相关,但总黄酮及总花色苷含量与花色表型的相关系数较高,是主要影响因子,这与谢燕等[39]对腊梅花色研究的结果一致。
3.2 花色苷对月季花瓣呈色起主要作用,黄酮类物质起辅助作用
月季花瓣中主要存在的花色苷主要为矢车菊类及天竺葵类色素,连接的糖苷包括葡萄糖及芸香糖。与王峰[1]和Mikanagi等[40]对月季花色苷物质鉴定的研究结果一致。月季花瓣中黄酮类物质主要为槲皮素及山奈酚的糖苷化、酰基化衍生物,在观赏植物中也多次被报道。张玲等[41]在对‘紫枝’玫瑰进行花青素苷检测时,类黄酮苷多以槲皮素和山奈酚的糖苷化、酰基化的衍生物为主。钟培星等[42]研究发现芍药中槲皮素-3-O-葡萄糖苷、槲皮素-7-O-葡萄糖苷等黄酮醇的含量与芍药花色呈现明显线性关系。在对8个品种月季花瓣颜色参数与花色苷组分进行相关性分析时发现,月季花瓣的颜色多由花色苷的种类和含量决定,黄酮类物质辅助花瓣呈色。矢车菊-3,5-葡萄糖苷是月季花瓣中主要的花色苷成分,这与之前的报道[43-44]相吻合。这是因为月季中含有一种特殊的矢车菊糖基转移酶,其经常在花色素苷元的2个不同位置上发生糖基化[19]。矢车菊-3,5-葡萄糖苷调控月季花瓣向紫红色发展,天竺葵-3,5-葡萄糖苷对橙、红色花的形成有一定的影响,这与李想等[28]的研究结果一致。8个品种月季中均含有丰富的芦丁,但相关性显示芦丁与花色表型并无相关性,这说明黄酮醇可能通过与花青素发生共色作用而产生色素沉着,从而辅助花瓣呈色。这与Gordillo等[45]的报道一致,其也发现花青素与黄酮醇共着色可以使花青素更稳定。
3.3 粉紫色品种‘薰衣草花环’为最适合进行花色改良的月季受体品种
Katsumoto等[46]在培育蓝色月季时,通过分子手段进行花色改良时能够积累蓝色色素的受体品种通常是一些花瓣中含有较多黄酮醇却积累少量的矢车菊类色素,并且具有较高的细胞液pH 值的月季品种。同时,结合之前的研究中也发现矢车菊素、天竺葵素和飞燕草素位于花色苷合成途径的不同方向,芦丁等黄酮醇类物质与花色苷发生共色素沉着,辅助花朵呈色,导致花瓣颜色向蓝紫色偏转。故选择含有丰富黄酮醇类前体物质以及对于存在竞争关系的色素含量较少的月季品种成为花色育种的理想品种。而在本研究中发现粉紫色品种‘薰衣草花环’正因为积累了高含量的芦丁和少量的矢车菊类色素而显著区别于其他品种,且其pH 值也显著高于其他品种,故后续可通过调控内源基因表达或转入外源基因的手段来改良花色。而白色品种‘藤冰山’和粉色品种‘瑞典女王’也含有适量的芦丁和少量的花色苷以及较高的细胞液pH 值,因此成为第二个合适的选择,其虽含有一定量的黄酮醇,但由于不含或仅含有少量的花色苷,导致可能缺少一些能够编码合成花色苷的酶基因,故推测需转入多个外源基因,重新构建花色苷合成通路,因此改良过程和‘薰衣草花环’相比较为繁琐,但具体的花色改良方案还需进一步研究。
