沙塘鳢池塘和稻田养殖水质指标和环境生物的比较
2024-03-02梁克勤徐宏亮郭红喜朱晓鸣
李 礼 梁克勤 李 波 徐宏亮 郭红喜 王 曼 王 杰 朱晓鸣 张 磊
(1. 中国科学院水生生物研究所, 武汉 430072; 2. 淮安市水生生物研究中心, 淮安 223002; 3. 阿拉善盟农业技术推广中心,内蒙古 735499; 4. 武汉市农业科学院, 武汉 430207)
水质指标、水体浮游生物和底栖生物三者都是评价水体状态的重要参数。在水产养殖过程中,通过水质指标可以判断水质优劣情况、水体富营养化程度及污染状况。浮游生物和底栖生物作为水体生态健康评价指标被广泛应用[1,2]。水体生态环境中的初级生产者——浮游植物, 其变化直接影响了水体中物质循环和能量流动[3], 是养殖生态系统的重要基础; 浮游动物是浮游植物的主要消费者之一, 作为浮游植物与更高消费者之间的主要营养联系[4], 其群落组成和结构变化对于水体生态系统也具有重要指示作用[5]。底栖动物生活史长、运动性较弱, 摄食浮游生物的同时又被其他鱼类所食, 可以灵敏地反映环境变化和人为扰动造成的影响[6,7]。
通过探讨鱼类不同养殖模式的水环境变化, 可以有效评估和筛选鱼类适宜及经济实用的养殖模式。杨智景等[8]通过比较河蟹单养和青虾-河蟹混养两种养殖模式水质指标及浮游动物的差异, 表明青虾-河蟹混养模式水体状况优于河蟹单养模式。宋庆洋等[9]研究稻虾共作水体中的水质指标和浮游植物的群落特征, 得出浮游植物的生长主要受氮和磷元素的限制, 阐明了水质指标和浮游植物的内在关系。本文研究了沙塘鳢池塘和稻田两种养殖模式水质指标、浮游生物及底栖动物群落结构和组成, 考察了相关指标的动态变化规律, 阐明了沙塘鳢两种养殖模式下水生态系统的差异, 发展并丰富了稻渔综合种养的养殖对象, 对沙塘鳢稻渔共生模式实践具有重要的指导意义。
1 材料与方法
1.1 试验条件
共计6个试验点, 其中3个试验池塘和3个试验稻田, 每组3个重复。每个试验池塘和试验稻田面积均为6667 m2。试验池塘水深为1.2 m, 进排水方便。试验稻田四周具有环沟(宽3.5 m, 水深1 m); 内部有3条纵沟(40 m×3 m×0.8 m)。试验期间稻田水位保持在0.2 m左右, 以保证沙塘鳢可在整块稻田中活动。试验池塘和试验稻田中的水引自当地同一进水渠。试验于2022年6月25日至10月23日开展。整个试验期间不使用化肥及农药。
1.2 试验鱼
试验鱼的种类为中华沙塘鳢(Odontobutis obscura)。沙塘鳢苗种均为淮安市水生生物研究中心自行繁殖的同批次鱼苗, 待秧苗活棵后进行投放。试验池塘放养沙塘鳢密度为3 尾/m2、平均规格为(0.956±0.083) g/尾。试验稻田放养沙塘鳢密度为1尾/m2,每尾沙塘鳢的平均规格为(0.986±0.039) g/尾。试验池塘和试验稻田中均放养相同质量[平均规格(1.94±0.13) g/尾]的抱卵青虾, 每667 m2放养1.5 kg。
1.3 样本采集及种类鉴定
各类样本于7月25日、8月27日、9月16日和10月8日进行采集, 分别对应着水稻的四个生长阶段为分蘖期(Tillering stage, TS)、拔节期(Stem elongation stage, SE)、扬花期(Flowering stage, FS)和成熟期(Ripening stage, RS)。每个试验池塘和试验稻田均设置7个采样点。现场使用便携式水质分析仪(YSI Incorporated, Yellow Springs, OH USA)于水下0.2 m测量池塘和稻田(环沟)的温度、溶解氧和pH。其他水质指标检测样本采用5 L的采水器采集, 混合后收集1 L水样进行测定。浮游动、植物水样采集均使用2.5 L的采水器采集。浮游植物定性样本采集使用25号浮游生物网在水体表层收集; 定量样本取混合后的水样, 用1 L的白色水样采集瓶收集。浮游动物定量样本取20 L混合后的水样经25号浮游生物网过滤后收集到50 mL透明样品瓶中。浮游植物定量样本加入鲁哥氏液固定保存; 浮游植物定性样本和浮游动物定性、定量样本均使用4%福尔马林固定保存。底泥样本使用索伯网(底面采样框尺寸25 cm×25 cm×5 cm, 网径40目)采集后用筛网(孔径0.45 mm)洗涤。于实验室中将洗涤后的剩余物置于白瓷盘中活体分拣, 分拣样本经甲醛溶液(7%)固定后进行底栖动物鉴定。
浮游植物和动物的相关实验操作主要参照《淡水浮游生物研究方法》[10]。浮游植物鉴定参照《中国淡水藻类: 系统、分类及生态》[11]等。浮游动物鉴定参照《中国淡水轮虫志》[12]和《中国动物志(淡水枝角类)》[13]等。底栖动物鉴定参照《中国淡水生物图谱》[14]等书籍。浮游植物和轮虫生物量采用体积换算法; 枝角类和桡足类采用回归方程法;底栖动物测量其体重, 并将其换算为密度(ind./m2)。
1.4 数据分析
本实验数据的统计及计算采用Excel 2020软件进行。数据的单因素方差分析(One-way ANOVA)及多重比较(Duncan检验)采用SPSS 19.0软件进行分析处理, 以P<0.05作为差异显著水平。采用Graphpad prism 9.0进行作图。
2 结果
2.1 池塘和稻田水质指标
4次采样, 池塘和稻田的温度和pH无显著差异(P>0.05)。2种养殖模式的水温随采样时间呈现下降趋势, pH和溶解氧呈波动变化状态。稻田的溶解氧始终高于池塘(表1)。养殖期间池塘和稻田水体中硝态氮、氨氮和亚硝酸盐氮含量整体均呈现下降趋势(图1)。在4次采样中, 2种养殖模式水体的硝态氮、氨氮含量均无显著性差异(P>0.05)。首次采样, 稻田亚硝酸盐氮值显著高于池塘(P<0.05)。至采样后期, 稻田硝态氮、氨氮、亚硝酸盐氮均低于池塘。

图1 沙塘鳢池塘和稻田养殖模式硝态氮、氨氮和亚硝酸盐氮动态变化Fig. 1 Dynamics of nitrate nitrogen, ammonia nitrogen and nitrite nitrogen in pond and rice-fish culture of O. obscura

表1 沙塘鳢池塘和稻田养殖模式水质指标动态变化Tab. 1 Dynamics of water quality indicators in pond and rice-fish culture of O. obscura
在整个种养周期内, 池塘和稻田水体总氮和总磷含量无显著差异(P>0.05), 2组水样的总氮含量呈现不断增长的趋势, 总磷含量波动性较大, 于第3次采样达最大值(图2)。池塘和稻田的叶绿素a含量呈现下降趋势, 且池塘水体中叶绿素a含量始终显著高于稻田(P<0.05; 图3)。

图2 沙塘鳢池塘和稻田养殖模式总氮、总磷含量动态变化Fig. 2 Dynamics of total nitrogen and total phosphorus in pond and rice-fish culture of O. obscura
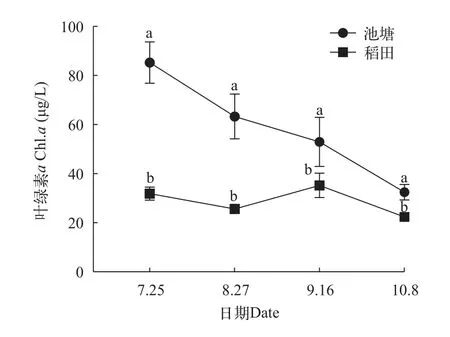
图3 沙塘鳢池塘和稻田养殖模式叶绿素a (Chl.a)含量变化情况Fig. 3 Dynamics of chlorophyll-a in pond and rice-fish culture of O. obscura
2.2 池塘和稻田浮游植物
4次采样, 池塘和稻田共鉴定出浮游植物6门40属。池塘有6门32属, 硅藻门7属、蓝藻门3属、绿藻门15属、甲藻门2属、裸藻门4属、隐藻门1属;D组有6门31属, 硅藻门8属、蓝藻门3属、绿藻门14属、甲藻门1属、裸藻门4属、隐藻门1属。其中23属在两种模式中都能鉴定出(表2)。在两种养殖模式下, 均是绿藻门占比最高, 其次是硅藻门。池塘和稻田独有的浮游植物均有8种, 其中根管藻(Rhizololeniasp.)、卵形藻(Cocconeissp.)、拟配藻(Spermatozopsissp.)、弓形藻(Schroederiasp.)、空球藻(Eudorinasp.)、实球藻(Pandorinasp.)、新月藻(Closteriumsp.)和裸甲藻(Gymnodiniumsp.)为池塘特有的浮游植物; 稻田特有的为辐节藻(Stauroneissp.)、菱板藻(Hantzschiasp.)、双菱藻(Surirellasp.)、多突藻(Polyedriopsissp.)、集星藻(Actinastrumsp.)、肾形藻(Nephrocytiumsp.)、蹄形藻(Kirchneriellasp.)和十字藻(Crucigenniasp.)。

表2 沙塘鳢池塘和稻田养殖模式浮游植物群落种类组成Tab. 2 Species composition of phytoplankton communities in pond and rice-fish culture of O. obscura
在整个采样周期中, 除第1次采样时, 池塘浮游植物密度和生物量显著高于稻田外(P<0.05), 其余3次采样, 池塘和稻田浮游植物密度和生物量均无显著差异(P>0.05)。从整体来看, 2种模式水体浮游植物密度和生物量的变化趋势基本相同, 于分蘖期达到最大值, 到拔节期出现显著下降, 于扬花期又出现增高, 最后在成熟期再次下降(图4)。

图4 不同时期沙塘鳢池塘和稻田养殖模式浮游植物密度、生物量的动态变化Fig. 4 Dynamics of phytoplankton density and biomass in pond and rice-fish culture of O. obscura at different periods
2.3 池塘和稻田浮游动物
池塘和稻田共鉴定出浮游动物3大类21属41种,其中轮虫12属24种(58.6%), 枝角类6属11种(26.8%),桡足类3属6种(14.6%)。池塘鉴定出轮虫20种、枝角类10种、桡足类5种; 稻田鉴定出轮虫18种、枝角类10种、桡足类6种。其中28种在两种模式中都能鉴定出。池塘和稻田独有的浮游动物种类均是6种, 其中直额弯尾溞(C.rectirostris)、角突臂尾轮虫(B.angularis)、壶状臂尾轮虫(B.urceus)、月形腔轮虫(L.luna)、凹顶腔轮虫(L.papuana)和螺形龟甲轮虫(K.cochlearis)为池塘特有的浮游动物; 稻田特有的浮游动物有秀体尖额溞(A.diaphana)、跨立小剑水蚤(S.kuali)、至点球轮虫(T.haerasolstitialis)、前节晶囊轮虫(A.priodontav)、盖氏晶囊轮虫(A.girodi)和四角平甲轮虫(P.quadricornis; 表3)。

表3 沙塘鳢池塘和稻田养殖模式浮游动物群落种类组成Tab. 3 Zooplankton community species composition in pond and rice-fish culture of O. obscura
在水稻分蘖期和拔节期, 池塘浮游动物密度和生物量均显著高于稻田(P<0.05)。在水稻成熟期时, 稻田浮游动物的密度和生物量高于池塘组(P<0.05)。从整体来看, 两种模式浮游动物的密度、生物量的变化趋势基本相同, 池塘和稻田浮游动物密度、生物量均在拔节期达到最高(图5)。分蘖期和拔节期池塘浮游动物生物量均显著高于稻田(P<0.05), 其后两次无显著性差异(P>0.05)。
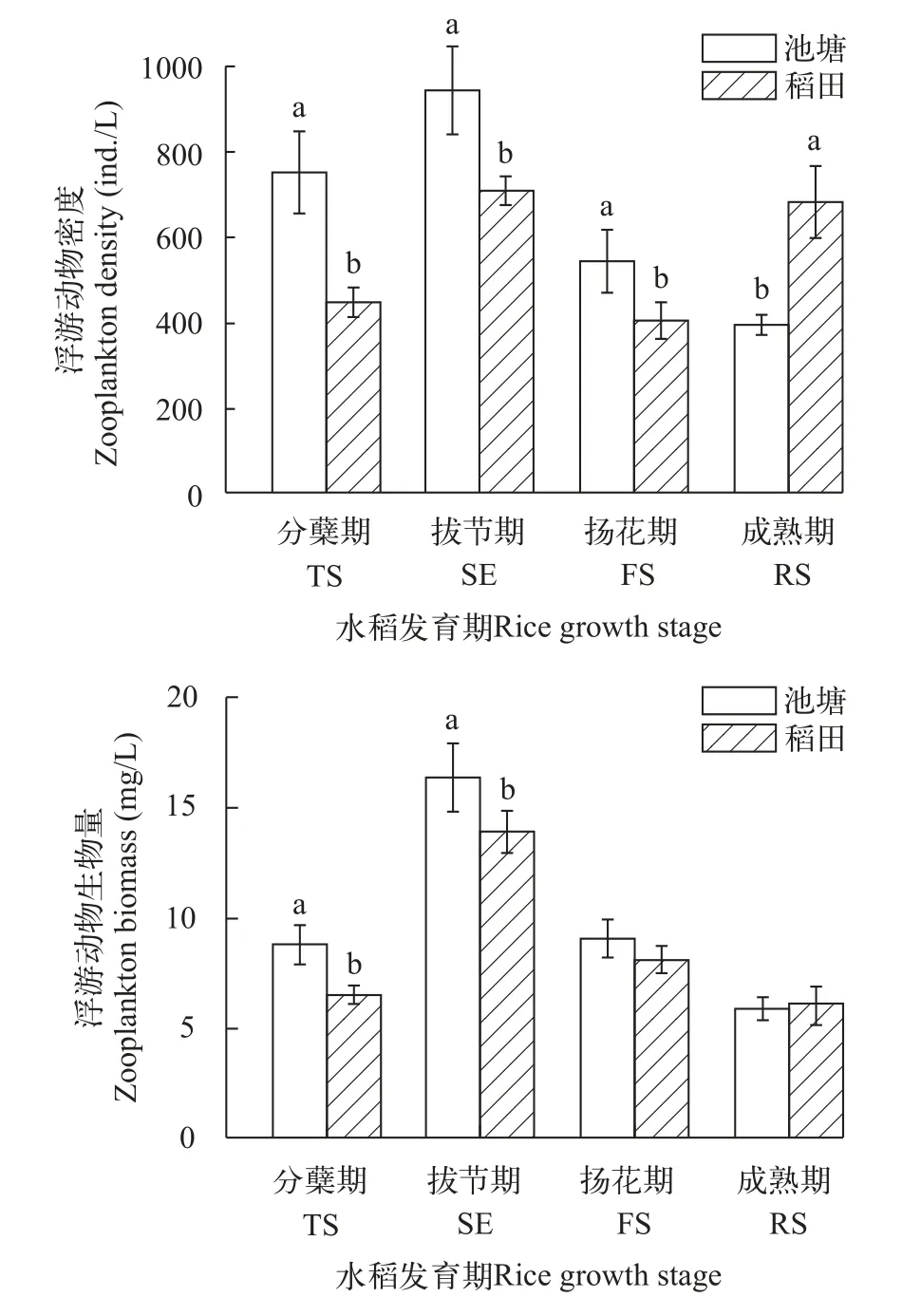
图5 不同时期沙塘鳢池塘和稻田养殖模式浮游动物密度、生物量的动态变化Fig. 5 Dynamics of zooplankton density and biomass in pond and rice-fish culture of O. obscura at different periods
2.4 池塘和稻田底栖动物
本试验共获得底栖动物9种(属), 其中包含环节动物3种、软体动物2种和节肢动物4种。尾腮蚓属(Branchiurasp.)在整个采样周期中均可检测到, 属于常见种; 而日本医蛭(Hirudo nipponia)只在少数采样周期能检测到, 属于偶见种, 也是稻田特有物种(表4)。整体上, 两种养殖模式底栖动物的种类和数量均呈下降趋势, 尾腮蚓属占比持续升高(表5)。

表4 沙塘鳢池塘和稻田养殖模式底栖动物群落种类组成Tab. 4 Species composition of zoobenthic communities in pond and rice-fish culture of O. obscura

表5 沙塘鳢池塘和稻田养殖模式底栖动物数量的动态变化Tab. 5 Dynamics of zoobenthic populations in pond and rice-fish culture of O. obscura (ind./m2)
3 讨论
3.1 池塘和稻田养殖模式的水质指标对比分析
水温、pH、溶解氧、TN、TP等是评价水体状态, 判断水质优劣的重要指标。水温受气候变化影响较大, 过高或过低的温度会改变水体中浮游植物的群落结构, 降低生态系统中浮游植物的稳定性[15]。例如水温较高时, 只有部分蓝藻能够存活并生长[16],当蓝藻成为水体中的优势种时, 会造成水华, 进而水中溶解氧含量下降, 导致水体恶化和水质变差。本研究中两种养殖模式的水温持续下降, 组间无显著差异。水体维持较高的pH和溶解氧, 有利于水环境充分发挥自身的净化能力。研究表明, 较高的pH(7.0—10.0)有利于提高氨氮的降解速率, 适宜与水质净化相关的微生物生长[17]; 较高的溶解氧可以减少水体中有害物质的产生、抑制氮磷等营养要素的过度释放[18]。本研究中两种养殖模式的pH无显著差异, 但稻田养殖模式的溶解氧含量始终高于池塘养殖模式, 这表明稻田养殖模式水体的自净能力可能高于池塘养殖模式, 稻田的水质更好。总氮和总磷是衡量水体污染的指标, 本研究中两种养殖模式的总氮含量除第一次采样外, 均无显著差异, 且随着时间的推进不断增加。而氨氮、亚硝酸盐氮和硝态氮的含量均不断降低, 表明养殖水体中有机氮含量在不断积累, 可能是由于随着沙塘鳢和青虾生长, 其排泄物的增加导致的。叶绿素a是评价水体富营养化的重要依据[19]。在两种模式下, 叶绿素a含量均呈下降趋势, 沙塘鳢池塘养殖模式的叶绿素a含量始终显著高于稻田养殖模式, 这表明池塘养殖模式的营养化水平可能高于稻田模式。通过水质指标的分析可以得出本研究中沙塘鳢稻田养殖模式的水质更佳, 营养化水平低于池塘养殖模式。
3.2 池塘和稻田养殖模式浮游生物及底栖动物的对比分析
在两种养殖模式下, 水体中浮游植物种类数没有差异, 共检测出浮游植物6门40属。池塘养殖模式检测出浮游植物6门32属, 高于路浩浩等[20]对中华绒螯蟹土池中鉴定出的5门16属19种。稻田养殖模式检测出浮游植物6门31属, 低于宋庆洋等[9]对稻虾共作水体鉴定出的7门124种, 高于Yuan等[11]对稻鱼共生系统水体检测出的6门38属。地域、所用水源及养殖生物等都可能造成浮游植物种类数的差异。绿藻门为池塘和稻田养殖模式水体中种类数最多的浮游植物种类, 与张露等[21]对稻虾共作水体浮游植物群落特征的研究结果和杨智景等[8]、龚进玲等[22]对池塘浮游植物种类的研究结果相同。池塘和稻田养殖模式下, 共检测出浮游动物种类有3大类21属41种。池塘养殖模式共鉴定出35种, 高于青虾-河蟹养殖池塘鉴定出的25种[8]; 稻田养殖模式共鉴定出34种, 低于吴嫔等[23]在赣北稻田水体中检测出的109种。池塘和稻田养殖模式共鉴定出底栖动物9种, 日本医蛭既是偶见种也是稻田养殖模式的特有物种。本研究检测出的底栖动物种类数低于李岩等[24]对稻蟹共作水体中鉴定出的10种, 但常见种为尾鳃蚓属的结论与李岩[24]的研究结果相同。
部分浮游生物与底栖动物可作为判断水体状态的指示物种, 例如盘星藻、席藻、小环藻等代表着水体处于富营养化状态[25]; 臂围轮虫、暗小异尾轮虫等也指征着水体富营养化[26]; 摇蚊幼虫、尾腮蚓、水丝蚓等为耐污型物种[27], 指示水体处于污染状态。两种养殖模式的浮游植物(盘星藻、席藻和小环藻)及底栖动物(尾鳃蚓属)都鉴定出指示水体污染的物种, 结合两种养殖模式总氮和总磷的指标,说明沙塘鳢池塘和稻田养殖模式都处于富营养化状态或高度污染状态。角突臂尾轮虫、壶状臂尾轮虫作为池塘养殖模式中的特有物种, 分别属于中污型和多污型水体指示物种[28], 而在稻田养殖模式下的特有浮游动物中未发现关于水体污染的指示物种, 这可以从侧面说明池塘养殖模式的水体污染更加严重。
两种养殖模式浮游植物的密度、生物量的变化趋势相同(下降、上升再下降)且均在分蘖期最高;浮游动物的密度、生物量在拔节期升至最高, 然后持续下降。前期大量的浮游植物为浮游动物提供了充足的食物来源, 导致其在拔节期不论是密度还是生物量均为最高。浮游植物密度和生物量首次下降可能是因为沙塘鳢和青虾的生命活动以及水稻对稻田水面的覆盖范围增加, 抑制了浮游植物的生长。在第3次采样时, 沙塘鳢对青虾及浮游动物的摄食, 导致浮游动物数量下降, 减轻了对浮游植物的摄食压力, 沙塘鳢养殖稻田及池塘浮游植物生物量有所回升。至水稻成熟期, 水温降低导致氮、磷等营养素的释放减少, 浮游植物密度及生物量显著下降, 继而浮游动物的生物量也下降。在本研究中, 稻田养殖模式的浮游动物生物量的变化趋势(先上升后下降)和常见种——尾腮蚓属, 与郑斯迪等[29]对浙江稻鱼青田的研究结果相同。在本研究中, 养殖后期, 池塘养殖模式和稻田养殖模式底栖动物的密度和生物量均下降, 可能是由于沙塘鳢和青虾的摄食导致的, 但由于环境因子和养殖鱼类对底栖动物的影响较为复杂, 是否因为养殖生物的生命活动造成的两种模式底栖动物的数量下降还未确定, 有待进一步的研究。
沙塘鳢池塘养殖和稻田养殖模式的水质指标和环境生物都指示两者处于富营养化状态, 但稻田养殖模式的水质略优于池塘养殖模式。
