未折叠蛋白反应参与胰腺癌化疗耐药研究进展
2024-02-27朱克祥
陈 浩 朱克祥
胰腺癌是预后最差的消化系统肿瘤,有着“癌症之王”的称号。胰腺癌虽发生率较低,但在全球癌症相关病死率中排第4位,诊断晚、转移早和放化疗不敏感是导致其不良结局的主要原因。肿瘤细胞多来源于胰腺导管或腺泡,其中胰腺导管腺癌(pancreatic ductal adenocarcinoma,PDAC)占比超过90%,整体五年生存率不足10%[1]。早期胰腺癌首选手术切除后辅助化疗,标准联合化疗FOLFIRINOX方案(5-氟尿嘧啶、亚叶酸钙、伊立替康、奥沙利铂)及吉西他滨联合nab(白蛋白结合)-紫杉醇方案可以显著提高胰腺癌患者生存率,但晚期患者受益有限。化疗耐药是导致晚期患者不良结局的重要原因,耐药受肿瘤代谢、增殖和微环境等多方面因素调控[2]。本文就未折叠蛋白反应如何参与胰腺癌细胞逃避化疗杀伤为化疗增敏提供思路。
未折叠蛋白反应是细胞在代谢压力、化学毒物等有害因素刺激下做出的应激反应,此时内质网腔内未折叠或错误折叠蛋白的积累会破坏内质网稳态,进而启动内质网应激及下游未折叠蛋白反应通路[3]。未折叠蛋白反应3条信号通路(图1)有各自独立的感受器,分别为需肌醇酶1(inositol-requiring enzyme 1,IRE1)、活化转录因子6 (activating transcription factor 6,ATF6)和蛋白激酶样内质网激酶 (protein kinase-like ER kinase, PERK),未应激时,这3种跨膜蛋白与内质网腔内分子伴侣葡萄糖调节蛋白78(glucose-regulated protein 78/binding immunoglobulin protein,GRP78/BIP)结合处于失活状态;当发生应激时,内质网腔内未折叠蛋白数量增加,触发这3种蛋白感受器与GRP78解离,继而级联激活下游信号通路,缓解内质网代谢压力[4]。
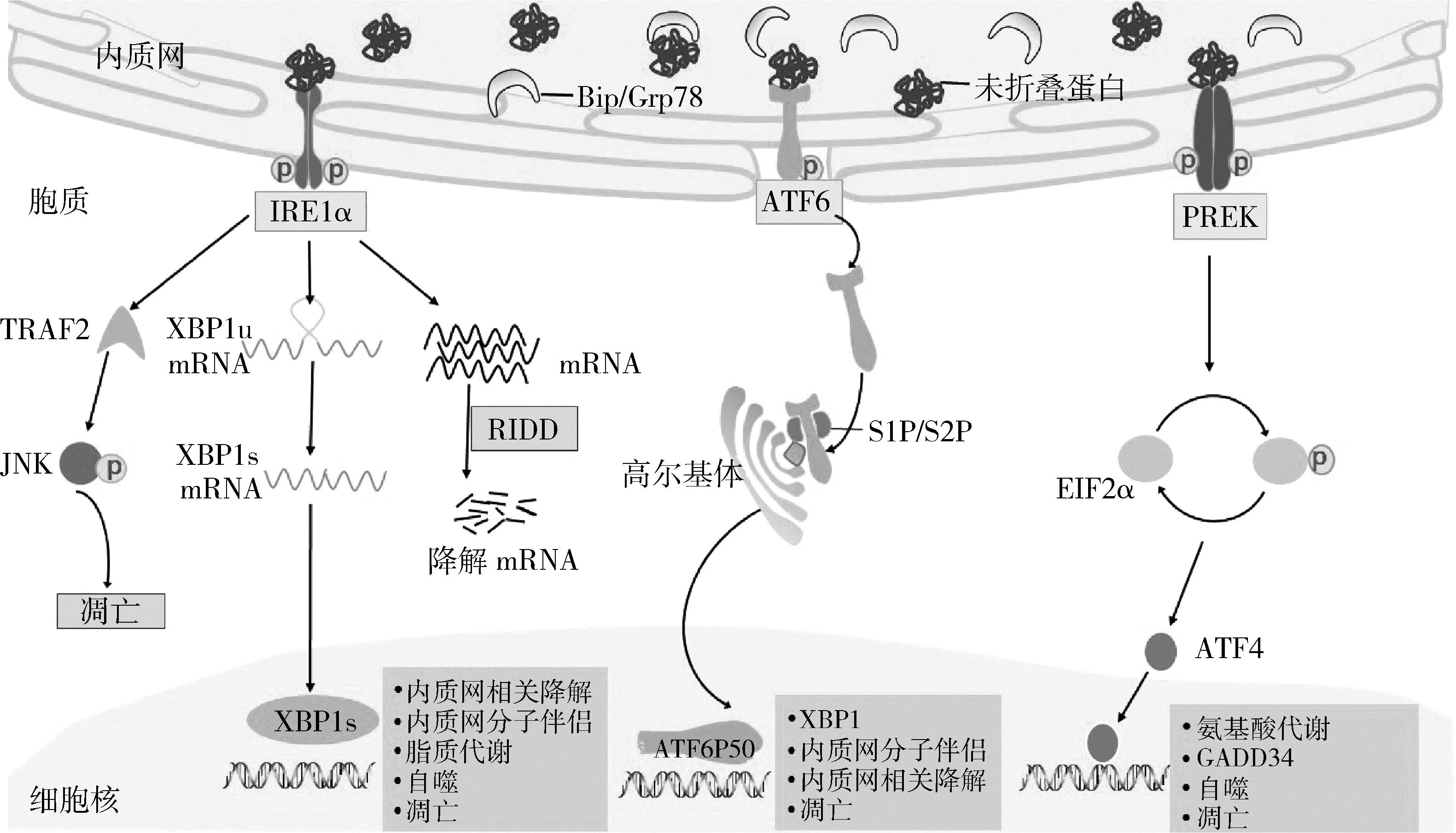
图1 未折叠蛋白反应激活3条信号通路示意图
未折叠蛋白反应能够在一定程度上减轻内质网负荷,促进细胞生存,化疗后肿瘤细胞通常处于高代谢负荷状态,未折叠蛋白反应为其提供了一条促生存通路。这一途径主要通过上调分子伴侣表达促进蛋白质折叠、增加异常折叠蛋白降解及减少蛋白翻译合成实现[5]。因此,深入研究未折叠蛋白反应与胰腺癌化疗耐药相关信号通路及机制具有重要临床意义,有望为提高胰腺癌化疗敏感度提供新思路。
一、未折叠蛋白反应信号通路
1.需肌醇酶1通路:需肌醇酶1(inositol-requiring enzyme 1,IRE1)为Ⅰ型跨膜蛋白,胞质端具有丝氨酸/苏氨酸蛋白激酶和核糖核酸内切酶活性。未折叠蛋白反应激活时,IRE1首先自身磷酸化形成二聚体,之后催化剪切X盒结合蛋白1(X-box binding protein 1,XBP1)mRNA,产生活化XBP1s。XBP1s是一种转录因子,进入细胞核后会启动一系列基因转录[6]。轻度应激时,其主要激活靶基因参与内质网相关降解(ER-associated degradation, ERAD)和IRE1依赖的蛋白降解(regulated IRE1-dependent decay, RIDD)途径,从转录、翻译等多个层次缓解内质网应激。严重内质网应激时,IERE1募集肿瘤坏死因子受体相关因子2(tumor necrosis factor receptor-associated factors 2,TRAF2),进一步诱导凋亡信号调节激酶1(apoptosis signal regulating kinase 1,ASK1)及其下游c-jun氨基末端激酶(c-Jun N-terminal kinase,JNK)的激活,诱导细胞凋亡[7,8]。
2.活化转录因子6通路:活化转录因子6(activating transcription factor 6,ATF6)是碱性亮氨酸拉链(basic leucine zipper,bZIP)家族转录因子,未折叠蛋白反应激活时转移到高尔基体上切割活化,剪切作用由高尔基体膜蛋白酶S1P(site 1 protease) 和S2P(site 2 protease) 介导,产生活性片段ATF6P50,ATF6P50随后进入细胞核与内质网应激反应元件(ER stress response element, ERSE)相互作用,激活GRP78、蛋白质二硫键异构酶(protein disulfide isomerase, PDI)、GRP94、钙联蛋白等下游分子伴侣基因,加强对未折叠蛋白处理能力。另外ATF6能协同IRE1增加XBP1转录,强化其降解未折叠蛋白能力。严重内质网应激时,ATF6能够激活CHOP(C/EBP-homologous protein)凋亡通路,诱导细胞凋亡[9,10]。
3.蛋白激酶样内质网激酶通路:蛋白激酶样内质网激酶(protein kinase like endoplasmic reticulum kinase,PERK)同样为Ⅰ型跨膜蛋白,胞质端具有丝氨酸/苏氨酸蛋白激酶活性。未折叠蛋白反应激活时,自身磷酸化形成二聚体,同时磷酸化真核翻译起始因子2α (eukaryotic initiation factor,eIF2α),磷酸化后eIF2α失活,后续mRNA翻译停滞,减轻内质网负荷。磷酸化PERK选择性激活活化转录因子4(ATF4),ATF4介导的细胞适应性反应称为整合应激反应 (integrated stress response,ISR),激活靶基因GADD34和CHOP,STAT4/GADD34通路为一条负反馈通路,能够去磷酸化P-EIF2α,导致整合应激反应信号转导终止,细胞恢复稳态。严重内质网应激时,ATF4激活CHOP凋亡通路,级联启动caspase-12表达活化,细胞凋亡程序启动[11,12]。
二、未折叠蛋白反应与胰腺癌耐药
1.未折叠蛋白反应与药物外排:ATP结合转运蛋白(ATP-binding cassette transporter,ABC)家族作为药物外排泵广泛参与多种肿瘤耐药,48种ABC转运蛋白中有19种可以外排抗癌药物,其中ABC亚家族B成员1(ABCB1)、ABC亚家族C成员1(ABCC1)、ABC亚家族G成员2(ABCG2)与多药耐药相关,能非特异性外排蒽环类、紫杉烷类、长春花生物碱类及酪氨酸激酶抑制剂等化疗药物,因此又被称为多药耐药相关蛋白[13]。
Dauer等[14]研究发现,与单用吉西他滨或紫杉醇处理胰腺癌细胞比较,联用siGRP78(GRP78小干扰RNA)后两种药物外排百分比分别由27.1%和2.9%下降至0.56%和0.68%,即干扰GRP78表达能够减少化疗药外排,此过程由ABC转运蛋白介导,并由转录因子NRF2调控。NRF2为PERK下游靶基因,其激活后上调应激反应蛋白、药物代谢酶和ABCB1编码的多药耐药蛋白1(multidrug resistance 1,MDR1)表达,提升对化疗药物抵抗力[15]。因此PERK/NRF2/MDR1轴可能参与胰腺癌化疗药物外排,抑制该代谢通路有望提高胞内药物浓度,强化其杀伤作用。
2.未折叠蛋白反应与自噬:化疗药物在诱导肿瘤细胞凋亡的同时也能诱导细胞自噬,与凋亡必定导致肿瘤细胞死亡不同,自噬对肿瘤细胞有一定程度保护性作用[16]。一方面,其能够帮助肿瘤细胞清除失活的细胞器和蛋白质, 减轻化疗药物的毒性损伤;另一方面,其会在应激时触发自噬依赖性细胞死亡, 从而逆转肿瘤细胞的耐药性。因此, 自噬在肿瘤化疗耐药中所起作用是双向的[17,18]。
Thakur等[19]研究显示,单用自噬抑制剂氯喹或STF-083010(IRE1通路抑制剂)处理胰腺癌细胞可抑制其自噬过程,但不足以延缓肿瘤生长,然而当与吉西他滨联合使用时,氯喹和STF-083010具有附加的抗肿瘤疗效。联合用药荷瘤小鼠肿瘤体积明显小于单独用药小鼠,双药联用毒性不良反应没有明显增加,但动物存活率更高。此外有研究表明,抑制乳腺癌细胞IRE1/JUK-beclin-1自噬通路能提高化疗药物疗效。因而未折叠蛋白反应IRE1/TRAF2/JUNK自噬通路抑制剂能在不增加毒性不良反应情况下增强化疗药物杀伤作用,自噬抑制剂与化疗药物联用有望改善胰腺癌耐药[20]。
3.未折叠蛋白反应与上皮-间充质转化:研究表明,上皮-间充质转化(epithelial-to-mesenchymal transition, EMT)与肿瘤侵袭、转移和耐药性相关,EMT肿瘤细胞丧失极性,与周围细胞及基质细胞黏附力降低,胰腺癌出现早期转移是其预后不良的重要原因,而上皮-间充质转化在胰腺癌早期转移中作用不可忽视,其同样在肿瘤耐药中发挥重要作用,上皮-间充质转化作为胰腺癌最显著和关键的特征之一,最终导致胰腺癌不良预后[21]。
吖啶黄(acriflavine,ACF)早期被当作染料及抗菌药使用,近年来研究显示,其可作为抗癌剂和缺氧诱导因子(hypoxia Inducible Factor,HIF)抑制剂。Dekervel等[22]研究发现,用TGF-β1(转化生长因子β1)或氯化钴模拟严重缺氧环境可诱导胰腺导管腺癌细胞发生上皮-间充质转化,再用吖啶黄处理可抑制此转化过程。基因富集分析显示,吖啶黄阻断了eIF2α磷酸化并减少ATF4翻译,从而抑制未折叠蛋白反应PERK/eIF2α/ATF4通路,即吖啶黄恢复了获得耐药胰腺癌细胞株的药物敏感度。因此,靶向PERK/eIF2α/ATF4通路可以抑制胰腺癌细胞上皮-间充质转化,有望提高胰腺癌化疗疗效。
4.未折叠蛋白反应与肿瘤干细胞:在导致肿瘤耐药的诸多因素中,肿瘤干细胞(cancer stem cells,CSCs)的作用不容忽略。在胰腺癌中,CSCs仅占胰腺癌细胞总数的0.2%~0.8%,但目前认为其广泛参与肿瘤侵袭、转移和耐药[23]。目前有研究显示,胰腺导管腺癌中CSCs并非实体而是一种“状态”,其具有独特的侵袭、代谢、增殖特性,能够自我更新并逃避化疗,未针对CSCs池靶向杀伤可能是胰腺导管腺癌化疗失效的部分原因,因此如何识别并清除CSCs将直接关系到胰腺导管腺癌化疗效果[24]。
Nomura等[25]研究发现,胰腺CSCs高表达CD133和GRP78,CD133能调节细胞代谢以维持低氧化还原水平,使其处于药物持续性耐受状态;GRP78与胰腺CSCs关联机制尚不明确,但有研究表明,在胶质瘤干细胞(glioma stem cells,GSC)中,GRP78/micRNA-205轴在GSC维持和抗辐照性上起重要作用[26]。Dauer等[27]在敲低GRP78表达后观察到胰腺CSCs克隆原性和自我更新性降低,化疗敏感度增加,但具体机制尚不清楚。因而CD133有望成为胰腺导管腺癌CSCs标志物,靶向未折叠蛋白反应核心GRP78相关通路可为清除胰腺CSCs提供了新治疗思路。
5.未折叠蛋白反应与黏蛋白:黏蛋白(mucins,MUC)是一类高相对分子质量糖蛋白,主要分为分泌型黏蛋白和跨膜黏蛋白两大类。胰腺导管腺癌的一个重要特征是分泌型黏蛋白产生异常,过量表达的黏蛋白形成物理屏障,阻止药物到达预定部位;以MUC-4为代表的跨膜黏蛋白相较正常组织,在卵巢癌、胰腺癌等肿瘤中明显差异表达,广泛参与肿瘤细胞代谢重排、免疫逃避、侵袭及耐药等过程,因此黏蛋白不仅在功能上参与肿瘤进展,同时有望成为新型肿瘤标志物[28]。
硼酸衍生物硼替佐米作为蛋白酶体抑制剂已用作血液系统恶性肿瘤和实体肿瘤的治疗药物,其本身能够削弱肿瘤细胞内泛素化标记异常蛋白降解能力,诱导细胞凋亡[29]。Wissniowski等[30]研究表明,沉默胰腺癌细胞MUC-4基因表达,硼替佐米诱导的细胞凋亡率上升,此凋亡过程由XBP-1剪切激活的CHOP凋亡通路介导。沉默MUC基因在减少黏蛋白合成量的同时强化未折叠蛋白反应介导的细胞凋亡水平,因而针对MUC-4/XBP-1/CHOP凋亡通路研究具有重要意义,相关研究有望削弱胰腺癌、结直肠癌等黏蛋白高表达肿瘤对化疗药物的抵抗作用。
三、展 望
未折叠蛋白反应的时长和强度直接决定了细胞最终结局。短期、轻度应激状态下,细胞激活适应性促生存通路,慢性或强烈应激会诱导细胞凋亡。目前针对肿瘤耐药提出的方案包括两方面:一方面,抑制未折叠蛋白反应介导的逃避化疗药物杀伤途径;另一方面,持续应激诱发细胞凋亡。但持续应激状态会对正常细胞产生毒性不良反应,在未找到靶向凋亡诱导剂之前,目前解决胰腺癌耐药主要研究方向仍是阻断其促生存通路。
未折叠蛋白反应体现了内质网对蛋白代谢失衡过程的动态调节,其调控机制精密且复杂,3条主要通路研究较完善,但仍有部分问题尚未解决,如决定GRP78合成数量的上游机制,GRP78如何感应并衡量蛋白代谢压力大小,GRP78激活3条通路有无倾向性,GRP78同时调控蛋白质、脂质和钙离子代谢,它们之间是否相互影响,内质网分子伴侣是一群功能复杂的“装配机器“,共同参与内质网腔内蛋白质折叠、组装及降解,除已知的通路外,可能还有未知的通路或调控机制参与未折叠蛋白反应,要切断未折叠蛋白反应为胰腺癌化疗耐药提供的促生存通路,上述问题都亟待解决。
综上所述,未折叠蛋白反应在胰腺癌化疗耐药多个方面发挥着重要作用,肿瘤细胞通过强化促生存相关通路增加对化疗药物抵抗力,遗憾的是目前相关研究仅停留在细胞或动物实验水平,后续药物研究及临床试验仍有待于完善,如何靶向打破肿瘤细胞内质网稳态诱导其凋亡有待于进一步研究。因此,改善胰腺癌化疗耐药研究仍具有很大潜力,选择性阻断胰腺癌未折叠蛋白反应逃避化疗药物杀伤途径,有望改善其不良预后。
