胰岛素对昆虫病原线虫卡森斯氏线虫0657L生长发育及其共生细菌伯氏致病杆菌0657L的影响
2024-02-26李茜童王兴铎钱秀娟
李茜童, 陈 红, 王兴铎, 钱秀娟
(甘肃农业大学植物保护学院, 甘肃省农作物病虫害生物防治工程实验室, 兰州 730070)
近年来,全球气候变暖、产业结构调整、耕作制度变革、外来生物入侵等因素导致我国农业害虫发生危害依然严重,害虫绿色防控仍然是现代农业发展中的重要环节(萧玉涛等, 2019; 陆宴辉等, 2023)。生物防治的应用和推广是践行绿色环保理念,维持生态平衡,促进可持续发展的重要手段,其中昆虫病原线虫(etomopathogenic nematodes, EPNs)是备受重视的一种生防因子,在农林牧草隐蔽性害虫的生防领域占据关键地位(杨丽元, 2020)。昆虫病原线虫是一种寄生性线虫,能够感染并杀死多种寄主昆虫,据统计对350种以上的害虫具有控制效能,以斯氏属Steinernema和异小杆属Heterorhabditis为主(王杰等, 2021;Zhangetal., 2021)。昆虫病原线虫能够自主搜寻并进入合适的宿主昆虫体内,破坏寄主的免疫反应,并有效地繁殖子代,子代继续感染新的寄主昆虫(Heryantoetal., 2022)。且由于昆虫病原线虫生命周期短,易于低成本大量人工培养繁殖,已经被广泛用于害虫生物防治,可与低毒农药结合使用控制破坏性的农业害虫,主要用于难以防治的钻蛀性害虫和地下害虫幼虫的防治(谢钦铭和张选辉, 2002; 曹翠玲等, 2009; 朱小芳等, 2021)。
昆虫病原线虫共生细菌隶属于肠杆菌科(Enterobacteriaceae)革兰氏阴性菌(钱秀娟, 2014),与昆虫病原线虫互惠互利,二者共同起到致死寄主昆虫的效果,斯氏属和异小杆属线虫分别与致病杆菌属Xenorhabdus和发光杆菌属Photorhabdus细菌共生(王杰等, 2021)。昆虫病原线虫致死寄主昆虫时,侵染期幼虫(infective juveniles, IJs)经口、肛门及体壁进入寄主昆虫体内,向寄主昆虫血腔释放肠道内携带的共生细菌,共生细菌大量繁殖并与昆虫病原线虫形成一个复合体打破寄主的生理平衡,在24~48 h内致死寄主(丛斌等, 2000; 方香玲等, 2008; 王立婷等, 2013; 詹发强等, 2015; 颜珣和韩日畴, 2016)。在整个侵染过程中,线虫为共生细菌提供生长繁殖的环境,共生细菌为昆虫病原线虫提供营养成分并产生毒素抑菌物,且为昆虫病原线虫的繁殖提供良好的环境条件。但线虫与细菌的共生关系也有选择性,每种昆虫病原线虫只能以与它共生的细菌培养,而特定的共生细菌也只能培养产生它的线虫(颜珣和韩日畴, 2016; 张潘杰等, 2021)。近期研究表明,嗜菌异小杆线虫Heterorhabditisbacteriophora对共生细菌的专一性较斯氏线虫更严格,不同种斯氏线虫可以被同种共生细菌培养,而嗜菌异小杆线虫则不可以(李茜童等, 2022)。
昆虫病原线虫作为生物防治因子,存贮和运输是其商品制剂开发中最大的障碍,在生产实际应用时其防效主要局限于线虫制剂的货架期(Georgisetal., 2006)。因此,为使昆虫病原线虫适应环境中复杂的生物和非生物因素影响并延长其制剂货架期,利用内源或外源化合物的补充延长寿命、提高存活率及增强致病力尤为重要(钱秀娟等, 2019)。胰岛素是由胰岛β细胞受内源性或外源性物质如葡萄糖、乳糖、核糖和精氨酸等的诱发而分泌的蛋白质类激素(彭竹清和郝友进, 2019),研究表明,控制胰岛素信号通路对昆虫及昆虫病原线虫同科生物秀丽隐杆线虫Caenorhabditiselegans的抵抗衰老,增强线虫的抗应激能力,延长线虫寿命,改善健康水平等方面具有重要的调控作用(李慧萍和韩日畴, 2007; 谢鹏飞等, 2018; 况琪斐等, 2021)。线虫与共生细菌在致死寄主昆虫的过程中均起到重要作用,胰岛素对共生细菌生长发育及线虫-共生细菌复合体的作用如何,目前报道尚少,因此研究胰岛素对昆虫病原线虫及其共生细菌的影响具有理论意义,同时对延长昆虫病原线虫制剂货架期及昆虫病原线虫在农业生产中充分发挥更广泛的应用潜力具有实践意义(Luetal., 2016)。
本研究以从甘肃地区筛选的抗旱性品系卡森斯氏线虫Steinernemakraussei0657L为材料,将不同浓度胰岛素滴加到共生细菌伯氏致病杆菌Xenorhabdusbovenii0657L的单菌落,观测菌落生长情况,测定其培养线虫寿命及运动和吞咽能力的方法,明确胰岛素的补充对昆虫病原线虫及其共生细菌生长发育的影响,以阐明胰岛素与昆虫病原线虫及其共生细菌的关系,为昆虫病原线虫的寿命延长研究、制剂化生产及防效的提高提供科学依据。
1 材料与方法
1.1 供试昆虫与线虫
大蜡螟Galleriamellonella由甘肃农业大学植物保护学院甘肃省农作物病虫害生物防治工程实验室提供,实验用大蜡螟3龄幼虫经滞育处理,以防止吐丝、化蛹。采用对蛴螬、韭蛆及模式昆虫大蜡螟致病力及生物学特性较好的昆虫病原线虫卡森斯氏线虫0657L,均由大蜡螟老熟幼虫培养、繁殖,悬浮液贮存于4 ℃备用。
1.2 试剂与器材
胰岛素母液(1 mL pH 2盐酸溶液加入1 mg胰岛素干粉,胰岛素干粉购于生工生物工程上海股份有限公司,纯度>95%);蒸馏水;75%酒精;林格氏液(蒸馏水1 000 mL,氯化钠8.6 g,氯化0.3 g,氯化钙0.28 g)。NBTB培养基:无菌水1 000 mL,琼脂15 g,LB琼脂预混干粉40 g,溴百里酚蓝0.01 g,摇匀,121 ℃,1 240 Pa灭菌20 min。LB培养基:营养肉汤8 g,琼脂15 g,酵母粉5 g,六水合氯化镁(MgCl2·6H2O) 2 g,蒸馏水900 mL,摇匀,121 ℃,1 240 Pa灭菌20 min。玉米糖浆7 mL,玉米胚芽油4 mL,蒸馏水89 mL,混合均匀,加入无菌的培养液中。TSY液体培养基:胰蛋白胨大豆肉汤40 g/L,酵母提取物5 g/L。
体视显微镜(Stemi 305,德国Zeiss公司),高压灭菌锅(GI54DWS,致微仪器有限公司),超净工作台(HCB-1300V,青岛海尔生物医疗股份有限公司),二氧化碳光照培养箱(SPX-250-GB-CO2,上海跃进器械医疗有限公司),冰箱(BCD-656 WDPT,青岛海尔股份有限公司),超纯水仪(GWB-2,北京普析通用仪器有限责任公司),电子天平(ER-180,AND),玻璃棒,酒精灯,9 cm培养皿,3.5 cm培养皿,移液枪,圆形滤纸,剪刀,接种环,封口膜,脱脂棉等。
1.3 卡森斯氏线虫0657L侵染大蜡螟后共生细菌的分离、纯化和鉴定
将经过75%酒精体表消毒的大蜡螟3龄幼虫(生理活性基本一致)各5头置于垫有两层无菌滤纸的培养皿中,吸取1.5 mL 3 000头/mL的卡森斯氏线虫0657L悬浮液侵染大蜡螟,25 ℃培养24 h后将大蜡螟第1对胸足末端剪破,从虫体尾部向前按压至被共生细菌侵染的大蜡螟的血液从伤口处滴落在NBTB培养基上。用接菌环蘸取大蜡螟体液划Z字型后密封培养基,置于25 ℃培养箱中,待菌体长出,吸收培养基中溴百里酚蓝染料且菌体呈绿色的则确定为昆虫病原线虫共生细菌(杨亚贤等, 2023)。将单菌落转移至新的NBTB平板培养基上,继续划线进行共生细菌的二次纯化,以此重复,直到培养纯化出均匀的单菌落。挑取其中的初生型单菌落转移至LB平板培养基,在25 ℃培养箱中培养72 h后将共生细菌单菌落挑取至TSY培养基中,在28 ℃ 180 r/min摇床中培养48 h,得到共生细菌发酵液,4 ℃保存备用。经课题组鉴定,卡森斯氏线虫0657L的共生细菌为伯氏致病杆菌Xenorhabdusbovenii0657L(张文德等, 2023)。在LB培养基上接种上述纯化的共生细菌伯氏致病杆菌0657L作为卡森斯氏线虫0657L的食物, 25 ℃恒温培养。
1.4 胰岛素对卡森斯氏线虫0657L寿命、运动能力和吞咽能力的影响的测定
在光学显微镜下将卡森斯氏线虫0657L怀卵大母虫挑至新的用划线法培养有伯氏致病杆菌0657L的LB培养基中,子代产出后将大母虫挑走,子代继续置于25 ℃中培养。为了抑制卡森斯氏线虫0657L产卵,子代幼虫期的第0-5天LB中含5 μmol/L五氟尿嘧啶,待卡森斯氏线虫0657L生长至第6天时转至普通的LB上,此时的线虫生长至同一生长时期J3。挑取NBTB培养基上菌落大小一致(挑取的菌量也基本一致)的伯氏致病杆菌0657L单菌落至直径3.5 cm的LB培养基上,随机分为8组,随后每个单菌落立即分别滴加10 μL胰岛素母液与林格氏液体积比为1∶500和1∶5 000及林格氏液(现配现用),另设1组不加胰岛素和林格氏液的空白对照,处理后每24 h在体视显微镜下用十字交叉法测量菌落直径1次并记录,计算1~6 d时伯氏致病杆菌0657L的生长速率,生长速率(%)=(Dt-Dt-1)/t×100(D为共生细菌单菌落直径;t为时间,单位d)。将同期化J3卡森斯氏线虫0657L转移到上述胰岛素处理48 h时的伯氏致病杆菌0657L的LB上培养,此时记为实验第0天。为保证食物充足和药物浓度,每3 d将卡森斯氏线虫0657L转移至新的接有胰岛素处理的伯氏致病杆菌0657L的LB上,每天监测卡森斯氏线虫0657L的存活情况,直至所有卡森斯氏线虫0657L死亡(卡森斯氏线虫0657L对铂丝的轻微机械触碰无反应即视为死亡),记录存活时间,计算寿命和存活率。每组至少3个重复,每个重复30条线虫。
随机选取胰岛素处理3 d(3龄幼虫), 7 d(4龄幼虫)和11 d(成虫)时的卡森斯氏线虫0657L挑取至未培养伯氏致病杆菌0657L的空白LB培养基上,使其在自由运动过程中除掉虫身黏附的共生细菌后,记录30 s内卡森斯氏线虫0657L头部来回摆动的次数(即为衡量卡森斯氏线虫0657L运动能力的标准),从一侧摆动到另一侧再摆回来视为1次。每组至少5条线虫,每条线虫统计至少统计3次,取其平均值作为最终结果。随机选取胰岛素处理3 d(3龄幼虫), 7 d(4龄幼虫)和11 d(成虫)时的卡森斯氏线虫0657L,直接在卡森斯氏线虫0657L生长培养基上对其30 s内吞咽次数进行统计,用铂丝轻轻触碰卡森斯氏线虫0657L,观察并记录卡森斯氏线虫0657L咽部中食道球前后抽动次数。每组至少5条卡森斯氏线虫0657L。每条卡森斯氏线虫0657L至少统计3次,取其平均值作为最终结果。
1.5 数据分析
卡森斯氏线虫0657L寿命数据采用long rank test统计方法,伯氏致病杆菌0657L菌落直径和生长速率以及卡森斯氏线虫0657L运动和吞咽数据采用LSD多重比较法进行差异显著性分析,所用分析及作图软件包括Excle 2016, SPSS 19.0和Origin 2021等。
2 结果
2.1 胰岛素对病原线虫卡森斯氏线虫0657L共生细菌伯氏致病杆菌0657L菌落直径的影响
1∶500和1∶5 000浓度胰岛素处理0~1 d时伯氏致病杆菌0657L生长较慢,所有单菌落均未长出; 2~6 d时,与空白对照组(CK)相比,1∶500和1∶5 000浓度胰岛素处理组伯氏致病杆菌0657L菌落直径显著增大(P<0.05),且低浓度(1∶5 000)胰岛素处理组菌落直径与高浓度(1∶500)浓度胰岛素处理组相比促进作用更明显,但二者之间没有显著差异(P>0.05)。在整个观察时间段内,除1 d时各处理组未生长外, 2~6 d时, 1∶5 000浓度胰岛素下, 菌落直径分别为2.89, 3.68, 4.06, 4.52和4.85 mm, 较空白对照组分别增大77.2%, 27.6%, 19.8%, 19.5%和17.6%; 与空白对照组相比, 1∶500浓度胰岛素处理组菌落直径分别增大54.0%, 20.1%, 14.2%, 15.1%和14.9%(图1, 2)。

图1 1∶500和1∶5 000浓度胰岛素下伯氏致病杆菌0657L培养3 d时的单菌落形态
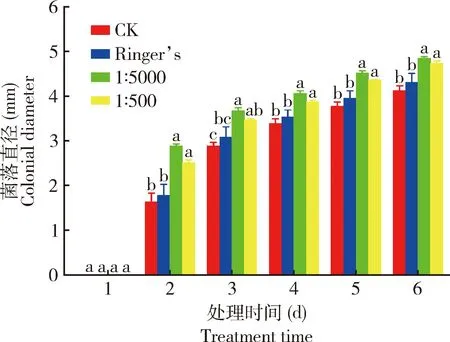
图2 1∶500和1∶5 000浓度胰岛素下伯氏致病杆菌 0657L菌落直径
2.2 胰岛素对伯氏致病杆菌0657L生长速率的影响
根据伯氏致病杆菌0657L的生长速率绘制曲线图(图3),伯氏致病杆菌0657L的生长速率在整个实验周期内呈现由快至慢的趋势,符合逻辑斯蒂生长曲线。1~2 d时,由于菌落生长缓慢,显微镜下观察不到明显菌落,即为延缓期;2~3 d时,菌落快速生长,即为对数生长期,共生细菌生长速度最快;1∶500和1∶5 000胰岛素处理组共生细菌生长速率较空白对照组(CK)和林格氏液组显著(P<0.05)加快,分别由空白对照组的68.0 μm/d提高至104.7和120.4 μm/d。1∶500胰岛素处理组较空白对照组和林格氏液组分别加快36.7和30.6 μm/d,1∶5 000浓度下,较空白对照组和林格氏液组分别加快52.5和46.4 μm/d,但高浓度(1∶500)处理组与低浓度(1∶5 000)处理组之间无显著差异(P>0.05)。3~7 d时,菌落的生长速率明显变慢, 后逐渐趋于平稳, 即为稳定期。

图3 1∶500和1∶5 000浓度胰岛素下伯氏致病杆菌 0657L生长速率
2.3 胰岛素对卡森斯氏线虫 0657L寿命的影响
结果表明(图4),取食不同浓度胰岛素处理的伯氏致病杆菌0657L可以延长自然状态下卡森斯氏线虫0657L寿命。与空白对照组(CK)相比,取食1∶5 000 胰岛素处理的伯氏致病杆菌0657L后卡森斯氏线虫 0657L的平均寿命由11.0 d延长至14.0 d, 延长了27.3%, 1∶500浓度胰岛素处理组较1∶5 000处理组的效果更好,卡森斯氏线虫0657L平均寿命为16.0 d,延长了45.5%。综上所述,不同浓度胰岛素均可以延长卡森斯氏线虫0657L寿命,但两处理浓度胰岛素处理组之间差异不显著(P>0.05)。

图4 取食1∶500和1∶5 000浓度胰岛素处理 48 h时的伯氏致病杆菌0657L后卡森斯氏线虫0657L的寿命
2.4 胰岛素对卡森斯氏线虫0657L运动能力的影响
结果如图5所示,与空白对照组(CK)相比,卡森斯氏线虫0657L在3龄幼虫期(胰岛素处理3 d时),胰岛素的补充可显著加快卡森斯氏线虫0657L的每30 s头部摆动次数,取食1∶5 000和1∶500胰岛素处理的伯氏致病杆菌0657L后卡森斯氏线虫0657L头部摆动频率由28.4次/30 s分别加快到44.4次/30 s和49.6次/30 s,分别显著提高了56.3%和74.7%(P<0.05)。卡森斯氏线虫0657L在4龄幼虫期(胰岛素处理7 d时),与空白对照组相比,取食高浓度(1∶500)胰岛素处理的伯氏致病杆菌0657L后卡森斯氏线虫0657L头部摆动频率为57.2次/30 s,显著提高了47.4%(P<0.05);而取食低浓度(1∶5 000)胰岛素处理的伯氏致病杆菌0657L后卡森斯氏线虫0657L头部摆动频率无显著差异(P>0.05)。而在卡森斯氏线虫0657L生命的后期成虫期(胰岛素处理11 d时),运动能力较3龄幼虫略强,较4龄幼虫期略弱,但与空白对照组相比,不同浓度的胰岛素加入对线虫的运动能力均有所提高,但并无显著差异(P>0.05)。综上所述,胰岛素的补充有助于卡森斯氏线虫0657L抵御运动能力的衰退,且不同浓度处理组的效果也不同,高浓度对线虫运动能力衰退的延缓作用更强。
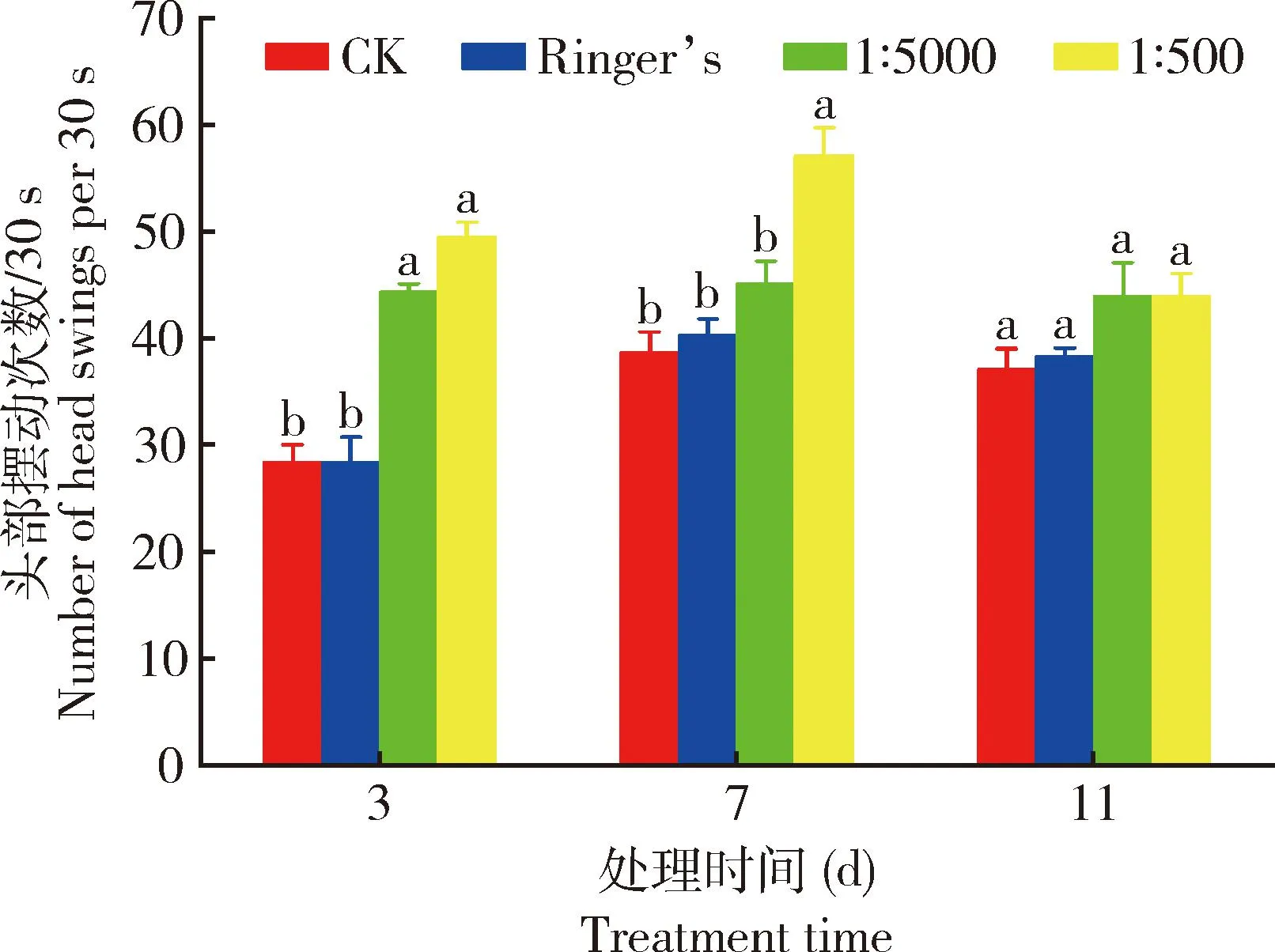
图5 取食1∶500和1∶5 000浓度胰岛素处理48 h时的伯氏致病杆菌0657L后卡森斯氏线虫 0657L的每30 s头部摆动次数
2.5 胰岛素对卡森斯氏线虫0657L吞咽能力的影响
结果显示(图6),胰岛素的补充在卡森斯氏线虫0657L 3龄幼虫期(胰岛素处理3 d时)、4龄幼虫期(胰岛素处理7 d时)和成虫期(胰岛素处理11 d时)均可显著提高卡森斯氏线虫0657L的吞咽能力。与空白对照组(CK)相比,3龄幼虫期时,取食1∶5 000和1∶500胰岛素处理的伯氏致病杆菌0657L后,卡森斯氏线虫0657L吞咽频率由11.0次/30 s分别提高到16.6次/30 s和18.2次/30 s,分别显著提高了50.9%和65.5%(P<0.05);4龄幼虫期时吞咽频率由14.2 次/30 s分别提高到18.0 次/30 s和19.2次/30 s,分别显著提高了26.8%和35.2%(P<0.05);11 d时吞咽频率由11.8次/30 s分别提高到14.6次/30 s和15.0次/30 s,分别显著提高了23.7%和21.1%(P<0.05)。而且同运动能力一样,成虫期较3龄幼虫期略强,较4龄幼虫期略弱。综上所述, 胰岛素能够提高卡森斯氏线虫0657L吞咽频率, 且不同浓度处理组促进作用略有差异, 高浓度较低浓度对吞咽能力的促进作用更明显。

图6 取食1∶500和1∶5 000浓度胰岛素处理48 h时的伯氏致病杆菌0657L后卡森斯氏线虫 0657L的每30 s吞咽次数
3 讨论与结论
昆虫病原线虫进入昆虫体内后,将其携带的共生细菌释放于寄主昆虫的血淋巴中,共生细菌迅速繁殖产生杀虫毒素,昆虫病原线虫自身分泌的毒素结合共生细菌产生的活性物质分解寄主昆虫组织,使寄主昆虫在短时间内患败血症死亡(王大业, 2020)。目前已经有对昆虫病原线虫共生细菌的杀虫蛋白和共生菌代谢产物开发的研究,结果表明,共生细菌具有杀虫活性主要是其代谢产物中的血腔活性物质起作用(彭刘亮, 2017),包括抑菌活性物质(黄武仁等, 2005; Fangetal., 2011; Binda-Rossettietal., 2016)、杀虫毒素(Bowenetal., 1998; Sheetsetal., 2011; Yangetal., 2012)、抗癌活性物质(Paiketal., 2001)等,在目前的相关研究中,昆虫病原线虫共生细菌常被用作生物制剂应用于害虫生物防治及生态环保领域。本研究结果表明,共生细菌作为昆虫病原线虫在寄主昆虫体内生长繁殖的重要介体,其生长发育可以受胰岛素的调控,共生细菌可以直接吸收利用胰岛素。本研究结果将在利用昆虫病原线虫共生细菌及其代谢产物提高对害虫的致病力方面提供新思路(吴文丹等, 2014; 王杰等, 2021),而对于共生细菌的具体杀虫机制以及能否利用胰岛素提高共生细菌代谢产物活性,尤其是血腔活性物质的活性,进而改善昆虫病原线虫共生细菌的杀虫效果等问题还尚不明确,这也将成为未来新的研究方向。
本研究结果表明,一定浓度胰岛素处理会促进共生细菌伯氏致病杆菌0657L菌落的生长(图1, 2),不同浓度的胰岛素对伯氏致病杆菌0657L生长速率的提高程度不同,低浓度胰岛素(1∶5 000)较高浓度胰岛素处理(1∶500)生长快(图3)。而有趣的是,高浓度胰岛素(1∶500)处理的伯氏致病杆菌0657L饲喂的卡森斯氏线虫0657L运动和吞咽能力均强于低浓度胰岛素(1∶5 000)处理的伯氏致病杆菌0657L饲喂的卡森斯氏线虫0657L(图5, 6)。在实验数据上表现为促进共生细菌生长速率加快的最佳浓度为1∶5 000,而促进线虫行为学能力增强的最佳浓度是1∶500。这表明,胰岛素对卡森斯氏线虫0657L生长发育及其体内分离出的共生细菌伯氏致病杆菌0657L的生长都有影响,但可能因为细菌和线虫的生长繁殖方式不同,对胰岛素的吸收和利用的方式也不同,因此共生细菌在形态学上表现出低浓度胰岛素处理下直径更大,而被取食后,仍然为高浓度更有利于线虫抵抗运动和吞咽能力衰退,这就与营养成分利用有关,具体机理还有待进一步考究。
胰岛素作为一种内分泌激素,与细胞生长、寿命、营养、生殖有关,还可促进蛋白质合成及脂肪储存,防止脂肪分解等。胰岛素在机体中的功能表现在多个方面,涉及受体结合,信号传导系统和缺陷的纠正等,进食和禁食期间能量的储存和释放,基本由胰岛素调控,是生物体生存的重要方面(Mayeretal., 2007; 顾世红和陈建国, 2009; Thevisetal., 2010)。胰岛素、胰岛素样生长因子(insulin growth factor, IGF)和胰岛素样肽(insulin-like peptide, ILP)通常与维持葡萄糖稳态有关,当机体内胰岛素分泌不足时,来自食物的糖分就只流动在血液中,无法进入细胞为机体所用,此时胰岛素的补充能够帮助食物中的糖分顺利进入机体内各组织细胞以提供能量(顾世红和陈建国, 2009),这种功能在昆虫(果蝇)和秀丽隐杆线虫中是保守的(Thevisetal., 2010; Graham and Pick, 2017),此外,胰岛素在代谢、生长、繁殖、成虫健康和衰老方面也发挥着重要作用(Hafen, 2004)。胰岛素信号通路是高度保守的衰老调控通路,包括Ins R/IRS-1通路和RAS/MAPK通路(Beale, 2013)。其中IRS-1可通过PI3K/Akt通路起到促进胰岛素影响葡萄糖代谢与脂质代谢的作用,该通路中胰岛素受体同源蛋白daf-16受基因daf-2负调控(张思雨等, 2022)。本研究发现,胰岛素处理在明显促进卡森斯氏线虫0657L共生细菌的生长的同时,有延长昆虫病原线虫寿命、抵御运动和吞咽能力衰退作用。这可以说明昆虫病原线虫作为共生细菌的携带者,能够通过胰岛素处理的特异性共生细菌间接地影响昆虫病原线虫体内新陈代谢信号的传导(共生细菌吸收胰岛素后长势更佳,为昆虫病原线虫提供稳定的生活环境和丰富的营养物质,其中蛋白质、脂类及少量糖类参与其体内细胞膜的组成,细胞膜完整保证了细胞的正常能量和物质交流),从而起到抗衰老的作用,初步揭示了胰岛素在昆虫病原线虫及其共生细菌生长发育中的作用。
研究表明,寿命的延长通常是与饮食限制(dietary restriction, DR)和热量限制(caloric restriction, CR)有关的,且饮食限制诱导的寿命延长的其中一个关键通路是IIS(胰岛素/胰岛素样生长因子1)信号传导(Nässel and Vanden, 2016)。在本研究中,吞咽试验为直接在线虫生长培养基上统计中食道球抽动次数,即线虫正常取食,说明并未造成饮食限制(即饥饿),而已有报道线虫(秀丽隐杆线虫)可在多种不同饮食限制方案下主要通过激活LET-363/mTOR和IIS两个营养物传感途径延长寿命(Greer and Brunet, 2009),结合上述证据,可推断胰岛素能够延长昆虫病原线虫的寿命,且排除胰岛素对卡森斯氏线虫0657L寿命的延长作用是由饮食限制引起的可能性,这与人类医学模式生物秀丽隐杆线虫C.elegans的寿命研究结果并不完全吻合。鉴于本研究中线虫摄食正常但最终得到寿命延长的结论,初步猜想是某些酶通过IIS通路中的靶标基因(如UTX-1)调控寿命(Greer and Brunet, 2009),共生细菌吸收胰岛素后为卡森斯氏线虫0657L提供丰富的营养成分,线虫充分利用营养物质至体内蛋白(酶)合成达到平衡时,相应靶标基因所在的信号通路关闭,基因表达量降低,衰老相关细胞功能维持稳态,寿命得以延长。这也恰好说明,寿命不仅在遗传水平上被调控,而且与新陈代谢调节和营养物质也密切相关。
研究使用胰岛素处理的共生细菌培养卡森斯氏线虫0657L,发现线虫生长早期,头部摆动次数和吞咽次数均较慢,生命中期,其头部摆动频率和咽泵吞咽频率均提高,直到生命后期又有一定程度的下降,说明线虫的衰老随着生命过程而发生,主要体现在运动和吞咽能力上。运动和吞咽与肌肉功能息息相关(Dall and Færgeman, 2019; 刘嘉榆, 2022),3个不同生长阶段中的胰岛素处理组运动和吞咽能力均较对照组强,表明胰岛素可以提高线虫的肌肉功能,起到有效抵抗衰老的作用。
综上所述,胰岛素的补充可以促进昆虫病原线虫共生细菌的生长,能够延长昆虫病原线虫的平均寿命并提高运动和吞咽能力。本研究仅证实了胰岛素在线虫生物学特征方面的作用,为昆虫病原线虫的制剂化提供了新的思路,也为昆虫病原线虫在生物防治领域的进一步高效开发利用提供了理论基础。为明确其作为调控机体代谢的重要物质在表观遗传学上是否也具有类似的作用,具体的作用机制如何,还需通过分子生物学技术手段的进一步探究。
