FBXO22表达对HPV阳性宫颈癌细胞生物学行为的影响
2024-02-24白雪高福贤王春晓黄新瑞
白雪, 高福贤, 王春晓, 黄新瑞
沧州市人民医院妇科一区,河北沧州061000
宫颈癌(cervical cancer,CC)是女性常见妇科恶性肿瘤,人乳头瘤病毒(human papilloma virus,HPV)尤其是HPV-16和HPV-18的持续感染在宫颈癌发生中起着重要作用[1]。尽管随着宫颈癌筛查和HPV疫苗普及,宫颈癌发病率略有下降,但近年宫颈癌发病有年轻化趋势,尤其对于无手术机会的晚期宫颈癌尚需要更有效的治疗方案。近年来,免疫治疗的兴起给晚期宫颈癌患者带来了新的希望。F-box蛋白22(FBXO22)是F-box蛋白家族中的一员,FBXO22在多种肿瘤的发生发展中起重要的作用,FBXO22可通过泛素化降解抑癌基因及其产物从而促进肿瘤恶性进程[2],如FBXO22通过泛素化降解核PTEN促结肠癌进程[3];FBXO22通过多聚泛素化降解P21促进肝癌细胞增殖,并引起肝癌患者不良预后[4];FBXO22还可促进抑癌基因LKB1降解,下调其表达水平,促进肺癌恶性进程[5];FBXO22通过调控不同底物,促进乳腺癌生长,增强[6]或抑制[7]其迁移和侵袭能力等。本研究初步通过TCGA数据库分析发现,FBXO22在宫颈癌患者中表达上调,提示FBXO22可能与宫颈癌密切相关,因此本研究旨在探讨FBXO22表达对HPV阳性宫颈癌细胞生物学行为的影响,进一步挖掘宫颈癌免疫治疗的新靶点,为临床研究提供依据。
1 材料和方法
1.1 材料与试剂
H8细胞(人宫颈上皮永生化细胞)、宫颈癌细胞株Hela(HPV-18+)、CaSki(HPV-16+)、SiHa(HPV-)(购自中国科学院细胞库);FBXO22抗体(购自美国Proteintech Group);辣根过氧化物酶标记的鼠源或兔源免疫球蛋白抗体(美国CST公司);Lipo2000(Life Technologies);DNA聚合酶(北京NEB生物技术有限公司);T4连接酶(Takara);Transwell小室(美国Corning);电泳仪(美国Bio-rad公司);PCR仪(瑞士Roche公司);流式细胞仪(美国BD公司)。
1.2 细胞培养及分组
H8细胞和CaSki细胞使用含10%胎牛血清的RPMI-1640培养基,Hela细胞和SiHa细胞使用含10%胎牛血清的DMEM培养基,置于37 ℃ 5%CO2恒湿培养箱中传代培养。Hela细胞和CaSki细胞均被分为空载体组(sh-NC组)和FBXO22敲低组(敲低组),或对照组和FBXO22过表达组(过表达组)。
1.3 真核过表达质粒和shRNA质粒的构建
真核过表达质粒构建:设计FBXO22引物序列(F:5′-CGGAGCACCTTCGTGTTGA-3′;R:5′-CACACACTCCCTCCATAAGCG-3′)并合成,进行PCR扩增。将FBXO22序列和过表达真核PBABE-Flag载体用T4 DNA ligase连接,转化入感受态细胞DH5α,加入抗性LB培养基,均匀涂布平板,倒置后37 ℃过夜。从每个转化平板上分别挑取4个克隆,摇菌并提取质粒,37 ℃酶切2 h。电泳并回收酶切的FBXO22片段,同时进行质粒测序验证正确性。FBXO22敲除shRNA质粒构建:将特异靶向FBXO22的shRNA片段与线性化pSIREN Retro-Q载体连接,构建特定基因的敲除质粒。靶向shRNA片段的序列为shFBXO22-1(sh-1,GTGTGGTCCTTGTCTTTGGTT)和shFBXO22-2(sh-2,CGCATCTTACCACATACAGTT)。同时,使用无靶向效应的序列插入到pSIREN Retro-Q载体构建空载体sh-NC组。
1.4 Hela、CaSki细胞过表达和沉默FBXO22基因的转染
取对数生长期细胞经胰酶消化后,调整细胞为1×105个/mL,分别将Hela和CaSki细胞接种于6孔板24 h后取出,分别用无血清培养基洗涤3次,放回培养箱培养至60%~70%汇合时开始转染。取200 μL无血清培养基,加入12 μL质粒和12 μL转染试剂,涡旋10 s后于室温静置15 min,缓慢滴入6孔板,轻柔摇晃混匀;6 h后细胞换液,继续正常培养备用。
1.5 qRT-PCR检测
用TRIzol试剂提取总RNA,反转录合成cDNA。PCR引物序列GAPDH:上游5′-GGAAGCTTGTCATCAATGGAAATC-3′,下游5′-TGATGACCCTTTTGGCTCCC-3′;FBXO22:上游5′-CGGAGCACCTTCGTGTTGA-3′,下游5′-CACACACTCCCTCCATAAGCG-3′。PCR反应条件:94 ℃5 min,94 ℃40 s,53 ℃45 s,72 ℃80 s,28个循环;72 ℃ 10 min。采用2-ΔΔCT公式计算表达量。
1.6 Western blotting检测
收集细胞后提取总蛋白,进行BCA法蛋白定量,凝胶电泳并转膜3 h,封闭1 h。加入一抗,4 ℃过夜孵育,TBST洗膜3次,每次10 min。加入二抗孵育1 h,TBST洗膜3次,每次10 min。使用ECL发光试剂,进行X片曝光、显影、定影,检测FBXO22表达。
1.7 MTT检测
收集细胞悬液,每组设置6个复孔,混匀后的细胞悬液每孔加入100 μL到96孔板。细胞贴壁5 h后更换为150 μL完全培养基,再加入20 μL 5 g/L MTT溶液,培养4 h;弃去上清,每孔加入150 μL DMSO溶液;常温摇床摇晃10~15 min。选择490 nm波长,在酶联免疫监测仪上测定各孔光密度值。重复检测10天,绘制细胞生长曲线。
1.8 细胞克隆形成实验
收集细胞,使用含10%胎牛血清的完全培养基重悬并计数。各实验组接种细胞700个/孔于6孔板中,每3天换液并观察细胞状态,培养14天。镜下拍照,每孔用PBS洗涤1次,加入1 mL 4%多聚甲醛固定40 min,再用PBS洗涤1次,加入结晶紫染液1 mL,染色15 min。PBS洗涤细胞,晾干后对整板及每孔拍照,观察克隆形成情况。
1.9 流式细胞术
取对数生长期的细胞经胰酶消化后,用4 ℃预冷PBS洗涤并重悬,1 500 r/min离心5 min,加入4 ℃预冷的70%乙醇吹打混匀,用封口膜封口后4 ℃冰箱过夜固定。检测前将固定的细胞2 000 r/min离心4 min,用4 ℃预冷的PBS洗涤2次,100 μL 100 mg/L RNase A和0.2% Triton X-100重悬细胞,37 ℃水浴30 min,加入1 g/L碘化丙啶50 μL混匀,室温避光孵育30 min。在流式细胞仪检测前用300目尼龙网过滤,上机结束后用FlowJo软件分析细胞周期时相分布。
1.10 划痕实验
在6孔板中用间距0.5 cm横线作标记,取对数生长期的细胞经胰酶消化后接种入6孔板培养过夜,直至单层细胞覆盖6孔板。用移液枪头在6孔板上垂直画一条直线,用PBS洗涤3次,使用无血清培养基在培养箱中培养;在显微镜下定期观察0、24 h,随机采集3张以上视野图像进行观察并测量划痕宽度。
1.11 Transwell侵袭实验
在Transwell小室涂布稀释后的基质胶,静置成膜。每组细胞设置3个复孔,在小室上腔中接种100 μL细胞稀释液,下腔中加入500 μL含有10%胎牛血清的培养液。Transwell小室置于培养箱中36 h。擦去小室中上层细胞,加入4%多聚甲醛固定10 min后用0.1%结晶紫溶液染色。镜下随机选取4个高倍视野计数,取4个视野均值作为穿膜细胞数。
1.12 统计学分析
2 结 果
2.1 各组细胞FBXO22表达水平的比较
FBXO22 mRNA和蛋白表达水平Hela、CaSki、SiHa细胞高于H8细胞,HPV阳性的Hela和CaSki细胞高于HPV阴性的SiHa细胞(P<0.05;图1)。提示FBXO22在宫颈癌细胞中呈现过表达,且在HPV阳性的宫颈癌细胞中过表达更为显著,因此Hela和CaSki被选定用于后续实验。

图1 各组细胞FBXO22表达水平的比较A为Western blotting;B为qRT-PCR。a为P<0.05,与H8细胞比较;b为P<0.05,与SiHa细胞比较。
2.2 FBXO22稳定敲低和过表达细胞系的构建
与sh-NC组比较,敲除质粒(sh-1和sh-2)转染细胞后FBXO22 mRNA和蛋白表达均降低(P<0.05;图2A),提示成功构建稳定敲低FBXO22基因的Hela和CaSki细胞,其中sh-1转染效率较高,因此sh-1被选定为敲低组进行后续实验。与对照组比较,过表达质粒转染细胞后FBXO22 mRNA和蛋白表达均增加(P<0.05;图2B),提示成功构建稳定过表达FBXO22基因的Hela和CaSki细胞。
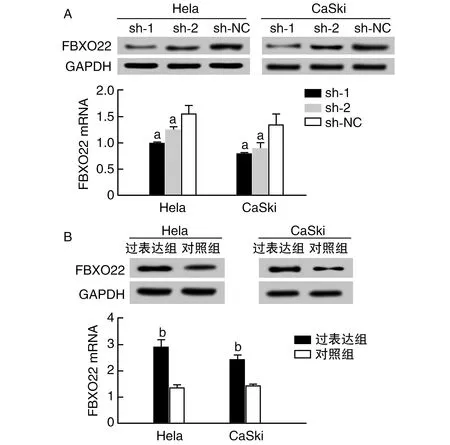
图2 FBXO22稳定敲低和过表达细胞系的构建A为敲低FBXO22时的Western blotting和qRT-PCR;B为过表达时的Western blotting和qRT-PCR。a为P<0.05,与sh-NC组比较;b为P<0.05,与对照组比较。
2.3 FBXO22稳定敲低和过表达对细胞增殖的影响
细胞生长曲线和细胞克隆形成实验发现,在Hela细胞和CaSki细胞中,敲低组细胞增殖能力低于sh-NC组,过表达组细胞增殖能力高于对照组(P<0.05;图3),提示FBXO22对Hela、CaSki细胞增殖具有促进作用。
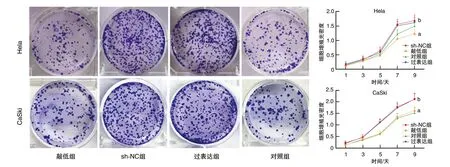
图3 FBXO22稳定敲低和过表达对细胞增殖的影响a为P<0.05,与sh-NC组比较;b为P<0.05,与对照组比较。
2.4 FBXO22稳定敲低和过表达对细胞周期的影响
流式细胞术分析发现,敲低组G2/M细胞百分率高于sh-NC组,过表达组G2/M细胞百分率低于对照组(P<0.05;图4),提示FBXO22能抑制Hela、CaSki细胞G2/M期阻滞,促进细胞异常增殖。
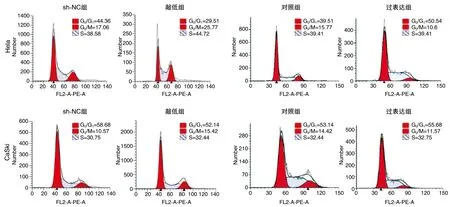
图4 FBXO22稳定敲低和过表达对细胞周期的影响
2.5 FBXO22稳定敲低和过表达对细胞迁移的影响
24 h后划痕宽度Hela细胞敲低组(59.2±0.68)μm大于sh-NC组(37.1±0.54)μm,CaSki细胞敲低组(57.6±0.64)μm大于sh-NC组(37.6±0.59)μm(P<0.05);Hela细胞过表达组(12.4±0.37)μm小于对照组(36.9±0.43)μm,CaSki细胞过表达组(14.5±0.67)μm小于对照组(35.2±0.68)μm(P<0.05;图5),表明FBXO22可增强Hela、CaSki细胞的迁移能力。

图5 FBXO22稳定敲低和过表达对细胞迁移的影响(100×)
2.6 FBXO22稳定敲低和过表达对细胞侵袭的影响
穿膜细胞数Hela细胞敲低组(69±7.23)个低于sh-NC组(102±5.64)个,CaSki细胞敲低组(65±6.44)个低于sh-NC组(101±4.59)个(P<0.05);Hela细胞过表达组(183±6.63)个高于对照组(102±5.64)个,CaSki细胞过表达组(178±3.47)个高于对照组(101±4.59)个(P<0.05;图6),提示FBXO22可增强Hela、CaSki细胞的侵袭能力。
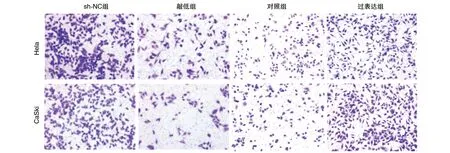
图6 FBXO22稳定敲低和过表达对细胞侵袭的影响(200×)
3 讨 论
世界卫生组织调查报告显示,2020年宫颈癌仍是女性常见的癌症之一[8],由于宫颈癌恶性程度高、侵袭性强、治疗方法局限、发病年轻化、预后较差、易复发[9-10]等特点,其发病率和病死率仍居高不下。HPV是宫颈癌主要致病因素,在癌病理组织中检出率达99%,HPV-16和HPV-18是高危人乳头瘤病毒,占宫颈癌发病的70%。为了探索宫颈癌的病因,开发更高效的治疗方法,进行宫颈癌细胞恶性生物学行为影响因素及机制的研究具有重要意义。
FBXO22第一次进入人们视野是作为p53的靶向基因[11],参与p53诱导的特异性蛋白的降解[12]。有研究表明,FBXO22是原发性乳腺癌的独立预后指标,FBXO22过表达患者的总生存期和无病生存期明显短于FBXO22低表达患者,敲除FBXO22抑制了裸鼠体外细胞生长和肿瘤形成,而过表达FBXO22增加了细胞活力[13]。FBXO22通过缺氧诱导因子-1α/血管内皮生长因子途径抑制乳腺癌细胞[14]、黑素瘤细胞[15]迁移、侵袭和血管生成。lncRNA 0006282通过miR-155上调FBXO22表达,促进胃癌的发展[16]。FBXO22通过抑制基质金属蛋白酶-9抑制人肾细胞癌迁移[17];lncRNA SNHG14通过miR-433-3p/FBXO22轴促进骨肉瘤进展[18]。FBXO22通过MAPK/ERK通路促进上皮性卵巢癌的生长和迁移并抑制自噬[19]。另一方面,FBXO22有时表现出肿瘤的抑制特性。在雌激素受体阳性和人表皮生长因子受体2型阴性乳腺癌患者中,FBXO22表达降低与预后不良紧密相关[20]。FBXO22低表达与乳腺癌患者低生存率有关[21]。同样有研究者在肾细胞癌组织的免疫组化分析中发现,与正常肾组织比较,肾细胞癌标本中FBXO22的表达水平降低,FBXO22在肾细胞癌患者中的低表达与肿瘤大小、TNM分期和生存率差异有关[17]。
尽管FBXO22与多种肿瘤的相关性已被证实,FBXO22对于宫颈癌发生、发展和迁移的作用却未见证实。因此,本研究设计并进行了相关实验,以期发现宫颈癌治疗的新靶点,为宫颈癌的个体化精准治疗提供理论基础。本研究首先通过TCGA数据库分析发现,FBXO22在宫颈癌患者中表达上调,随后通过qRT-PCR和Western blotting验证了HPV阳性Hela细胞和CaSki细胞的FBXO22表达水平较HPV阴性的SiHa细胞和H8细胞显著上调。为了进一步确定FBXO22在HPV阳性宫颈癌发生发展中的作用,本研究选择了HPV阳性的Hela和CaSki细胞,利用细胞功能缺失或获得的方法,分别构建了FBXO22敲除和过表达模型,检测构建模型后FBXO22的表达水平,进行了一系列细胞功能实验,结果表明,FBXO22低表达下调了FBXO22 mRNA及蛋白的表达水平,抑制了HPV阳性Hela和CaSki细胞的增殖、迁移及侵袭功能,而FBXO22过表达则可上调FBXO22 mRNA及蛋白的表达水平,促进HPV阳性Hela和CaSki细胞的增殖、迁移及侵袭功能。
综上,本研究结果显示,FBXO22对宫颈癌细胞的增殖、迁移和侵袭等恶性生物学行为均有明显的促进作用。但关于其调控宫颈癌细胞恶性行为的具体机制尚不明确,需要进一步的实验研究证实。
