HLA-DM 基因多态性与脊髓灰质炎疫苗诱导抗体应答的相关性
2024-02-24齐汝楠刘舒媛
齐汝楠,史 磊,刘舒媛,李 菁,史 荔
(中国医学科学院 &北京协和医学院医学生物学研究所,云南 昆明 650118)
脊髓灰质炎是严重危害儿童健康的急性传染病,接种脊髓灰质炎减毒活疫苗(oral poliomyelitis vaccine,OPV)和脊髓灰质炎灭活疫苗(inactivated poliomyelitis vaccine,IPV)可以有效控制脊灰的传播,但现有脊灰疫苗的保护效果很难达到100%[1]。受种者遗传背景是影响疫苗免疫效果的重要因素,研究表明脊灰疫苗诱导抗体应答的遗传度为60%[2]。接种脊灰疫苗后,外源性抗原在内体溶酶体中被水解,HLA-DM 分子催化II 类分子相关的恒定链多肽(class II-associated invariant chain peptide,CLIP)从MHC II-CLIP 复合体中解 离,抗原肽与MHC II 类分子结合后表达在专职性抗原呈递细胞(antigen presenting cells,APC)表面,被CD4+T 细胞的TCR 识 别[3−5]。DM 分子是抗原被有效呈递的关键因素,可稳定空载MHC II 类分子构象阻止其发生变性失活,促进高亲和力抗原肽的装载并保证MHC II-抗原肽复合体在APC表面可较长时间的稳定表达[6−8]。DM 分子由DMA 和DMB 基因编码组成,研究表明DM 与多种自身免疫性疾病和感染性疾病的发病风险有关[9−11],不同DMA 和DMB 等位基因组合编码的DM 分子在功能活性上存在差异,继而影响抗原肽在APC 表面的呈现和对CD4+T 细胞的激活[12]。本研究将探讨HLA-DM 基因多态性对脊灰疫苗诱导抗体应答的可能作用。
1 资料与方法
1.1 研究对象
以355 名2~3 月龄广西壮族自治区的健康婴幼儿为研究对象,所有个体均无脊髓灰质炎疫苗接种史及其他可能影响试验评估的情况,由监护人签署知情同意书并通过广西伦理审查委员会的批准(批准文号GXIRB2017-0009-6)。受试者在2 月龄和3 月龄时分别接种1 剂IPV,4 月龄接种1 剂bOPV,免疫前和第3 针免疫后第28 天采集受试者静脉血,提取外周血基因组DNA,检测血清中Ⅰ、Ⅱ、Ⅲ型脊灰病毒中和抗体滴度。免疫前脊灰病毒中和抗体效价 < 1:8 且免疫后中和抗体效价≥1:8 或免疫前中和抗体效价≥1:8 且免疫后中和抗体效价≥4 倍增长的个体为抗体阳转组,不符合抗体阳转组标准的个体为抗体非阳转组。
1.2 HLA-DMA、HLA-DMB、HLA-DOA 和HLADOB 基因分型
根据世界卫生组织HLA 命名委员会的命名原则,采用Sanger 测序法对HLA-DMA 外显子3 和HLA-DMB 外显子2/3 中的10 个SNPs 进行基因分型,见表1~2。扩增引物信息见表3。扩增体系为12.5 µL Takara PrimerSTAR Max DNA Polymerase(2×),0.5 µL 正向引物(10 pmol/µL),0.5 µL 反向引物(10 pmol/µL),1.5 µL DNA 样本(20 ng/µL),加入ddH2O 至总体系为25 µL,反应条件为98 ℃预变性2 min,98 ℃变性10 s、58 ℃退火5 s,共30 个循环,最后72 ℃延伸5 s。

表1 HLA-DMA 基因分型SNPs 位点信息Tab.1 HLA-DMA genotyping SNPs locus information
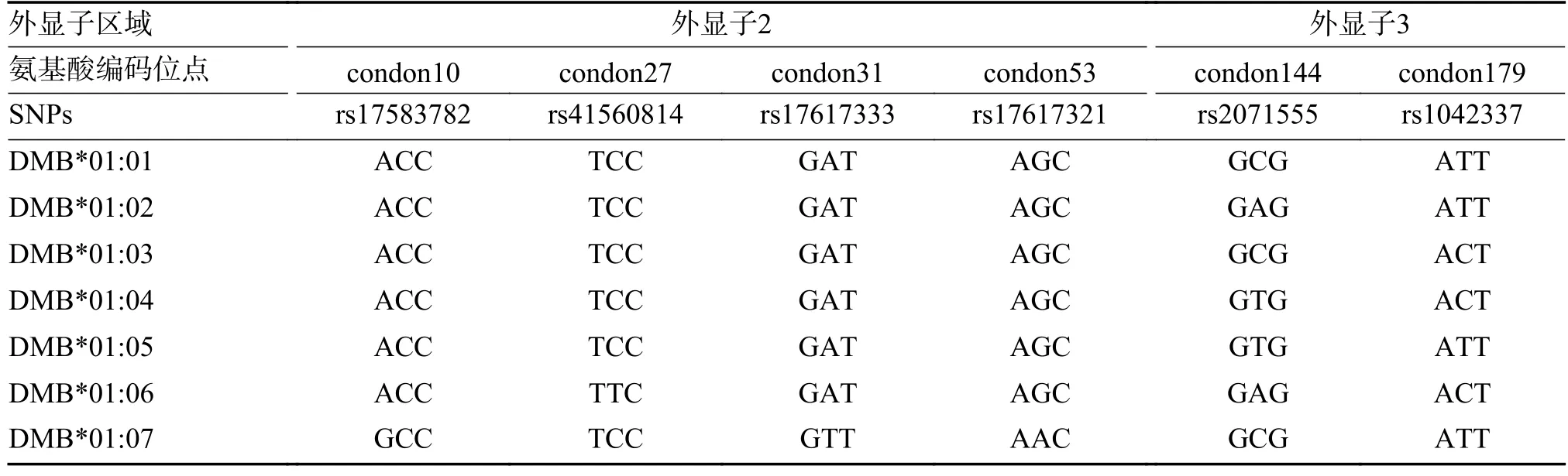
表2 HLA-DMB 基因分型SNPs 位点信息Tab.2 HLA-DMB genotyping SNPs locus information
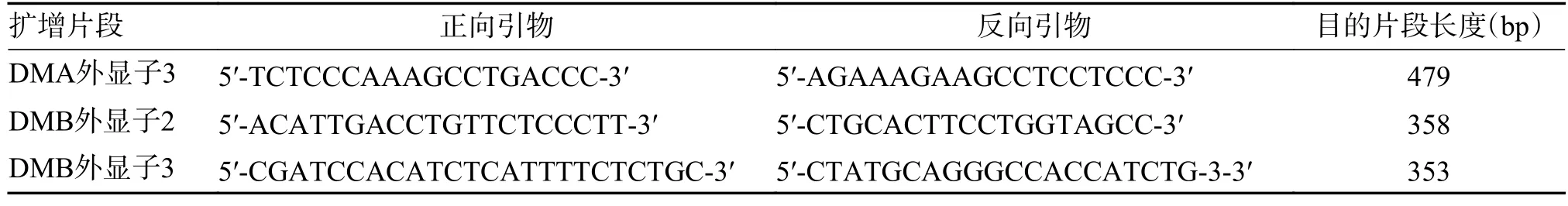
表3 引物序列及片段大小Tab.3 Primers sequence and fragment size
1.3 统计学处理
在Pypop 0.7.0 软件中对分型结果进行哈迪-温伯格平衡(hardy-weinberg equilibrium,HWE)检验。采用SHEsis 软件分析基因位点间的连锁不平衡,构建HLA-DMA-DMB 单倍型。应用SPSS 23.0 中的卡方检验(χ2)比较不同性别、民族、DM各等位基因与基因型在抗体阳转组和非阳转组之间的分布,P< 0.05 为差异有统计学意义。
2 结果
2.1 样本基本情况
纳入研究对象的355 名婴幼儿由196 名男性和159 女性组成,其中114 名是汉族,241 名是壮族。各型抗体阳转组和非阳转组间的性别和民族分布,差异无统计学意义(P> 0.05),见表4。
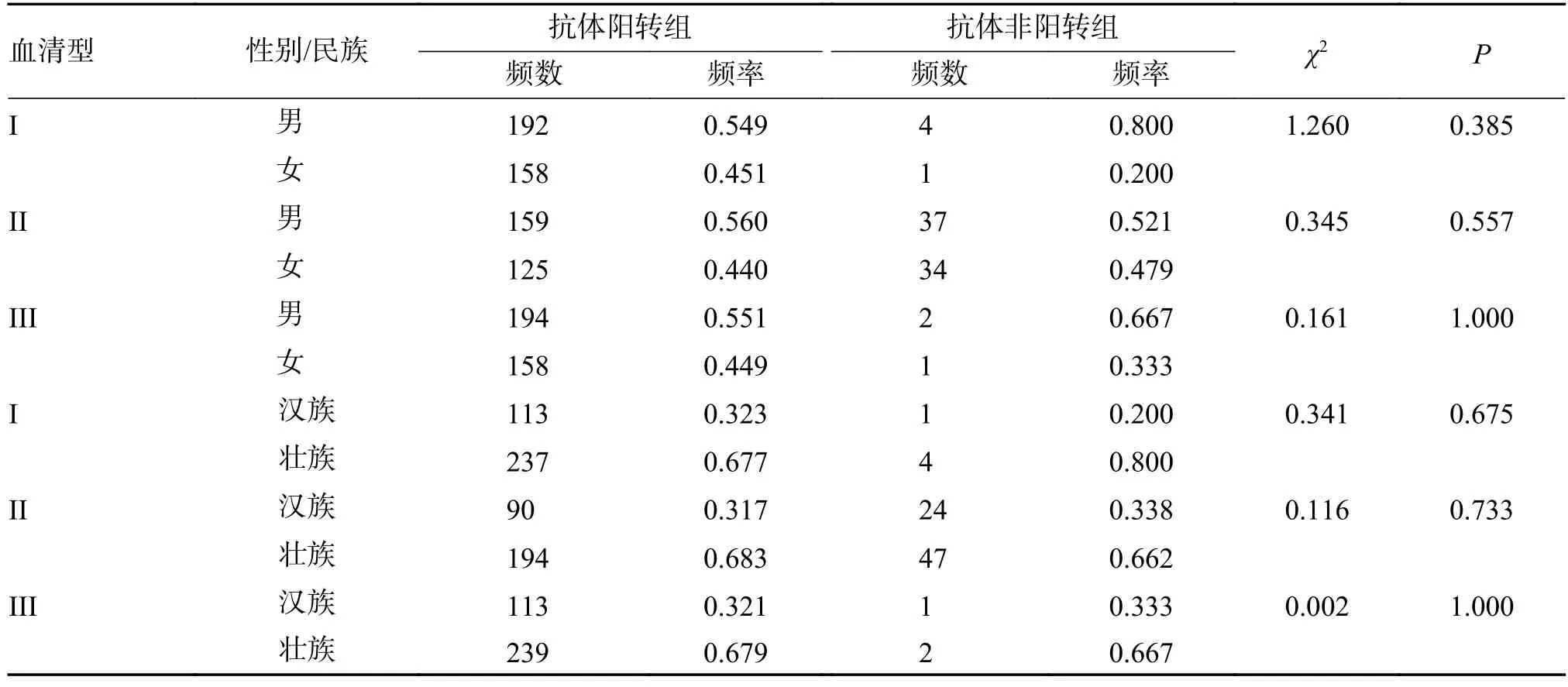
表4 研究对象基本情况Tab.4 Basic information on research subjects
2.2 等位基因分析
DMA 和DMB 基因频率分布均符合HWE(P>0.05)。脊灰I 型中DMA*01:02 在抗体非阳转组中的频率高于阳转组(P=0.022),DMB*01:01 在抗体非阳转组中的频率高于阳转组(P=0.021)。脊灰II 型中DMA*01:02 在抗体非阳转组中的频率高于阳转组(P=0.014),见表5。
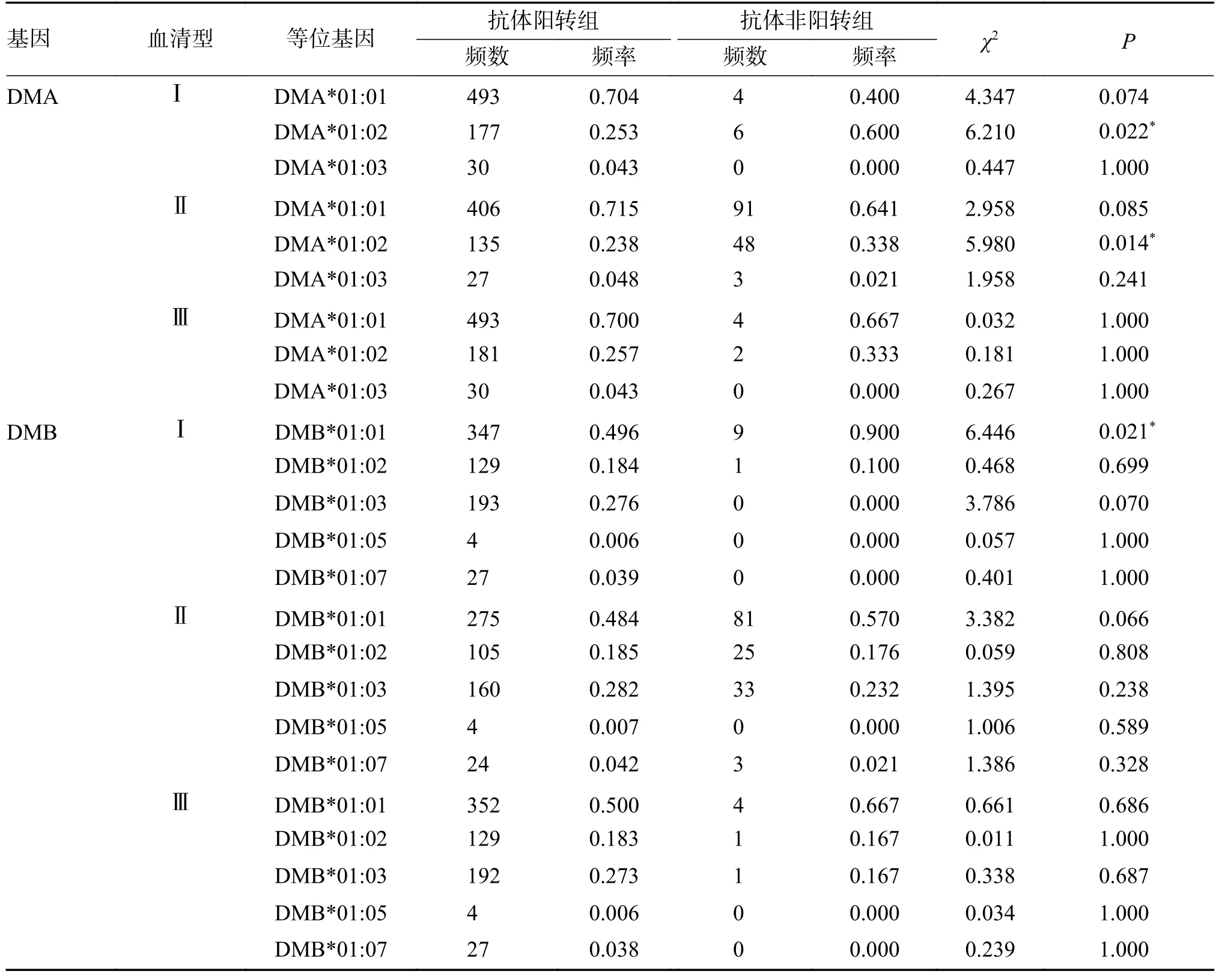
表5 DMA 和DMB 各等位基因在抗体阳转组和非阳转组中的频率分布Tab.5 Comparison of frequencies of alleles of DMA and DMB between two groups
2.3 基因型分析
脊灰I 型中DMB*01:01/DMB*01:01 在抗体非阳转组中的频率高于阳转组(P=0.020)。脊灰II型中DMA*01:02/DMA*01:02 在抗体非阳转组中的频率高于阳转组(P=0.001),DMB*01:01/DMB*01:01 在抗体非阳转组中的频率高于阳转组(P=0.003),见表6。
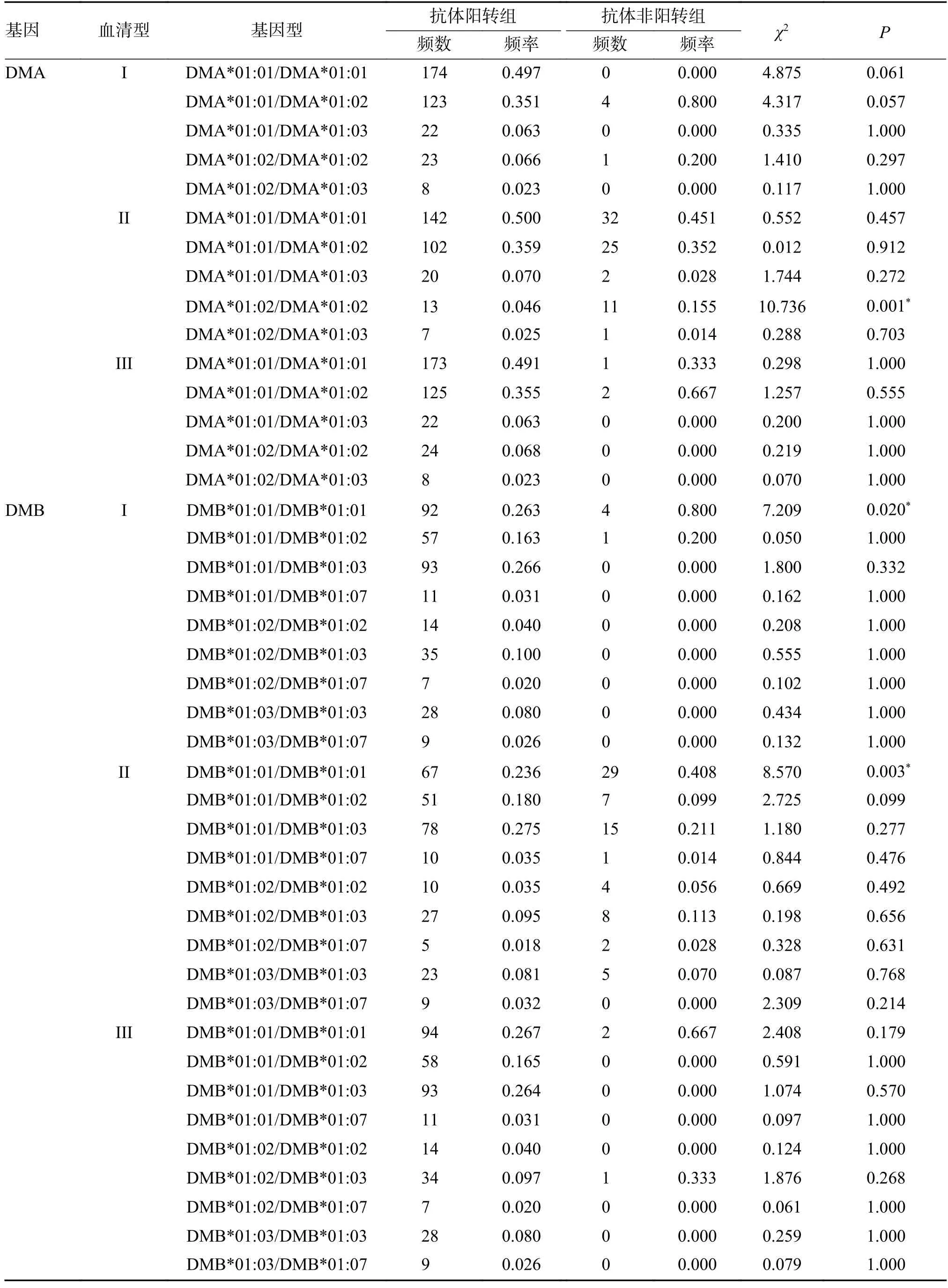
表6 DMA 和DMB 各基因型在抗体阳转组和非阳转组中的频率分布Tab.6 Comparison of frequencies of genotypes of DMA and DMB between two groups
2.4 单倍型分析
脊灰I 型中DMA*01:02-DMB*01:01 单倍型在抗体非阳转组中的频率高于阳转组(P=0.017)。脊灰II 型中DMA*01:02-DMB*01:01 单倍型在抗体非阳转组中的频率高于阳转组(P=0.008),见表7。
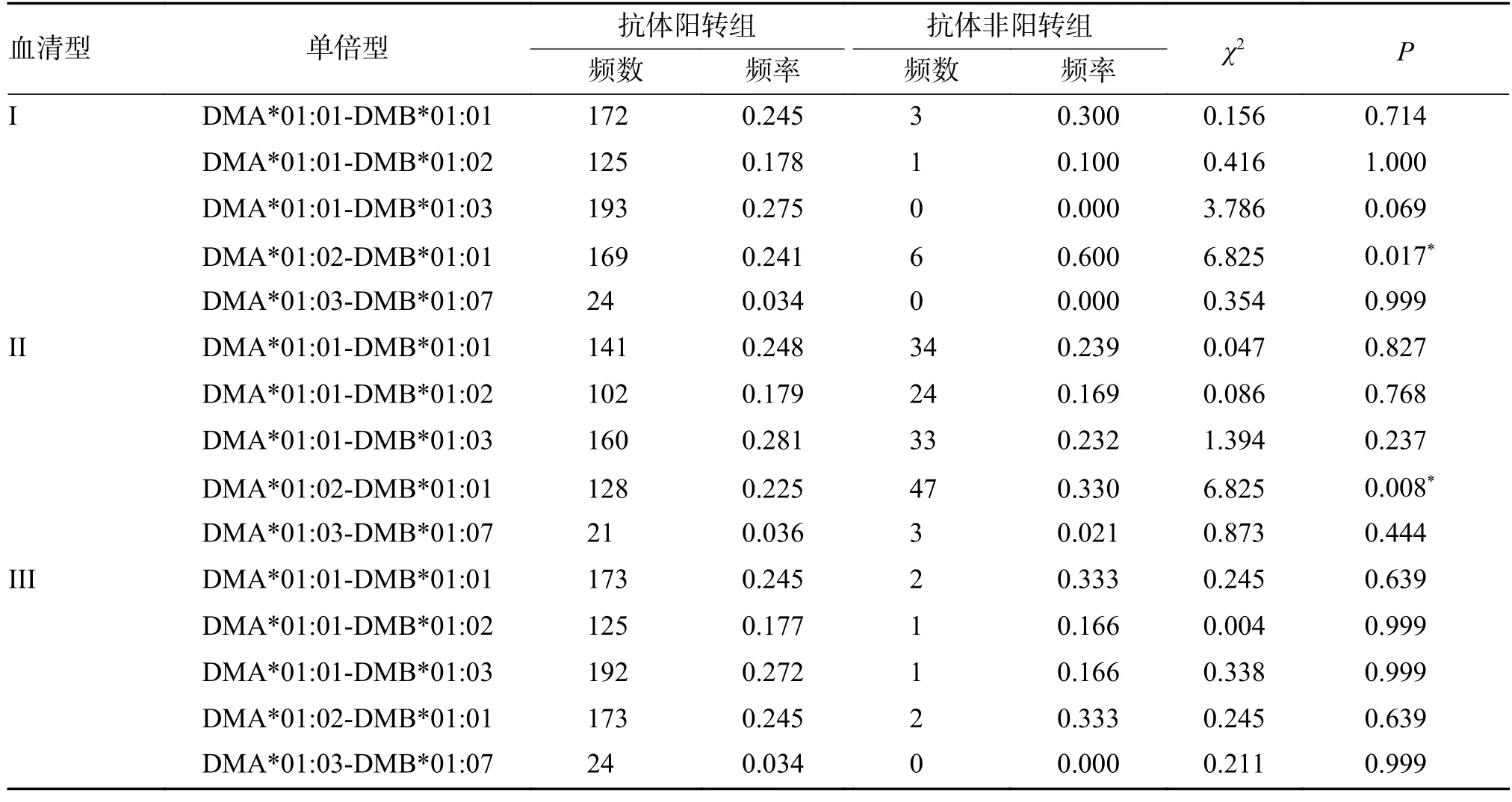
表7 DMA-DMB 单倍型在抗体阳转组和非阳转组中的频率分布Tab.7 Comparison of frequencies of genotypes of DMA and DMB between two groups
3 讨论
DMA 和DMB 基因的多态性可能会改变DM异二聚体结构,导致DM 分子催化肽交换功能变化,从而影响抗原肽的呈递[12]。本研究发现DMA*01:02 和DMB*01:01 在脊灰II 型抗体非阳转组中的等位基因频率高于阳转组。其他研究中,DMA*01:02 和DMB*01:01 在I 型糖尿病患者中的频率低于对照组[13],DMA*01:02 与银屑病发病风险相关[10],类风湿性关节炎患者中DMA*01:02的频率高于健康对照组[11],DMA*01:03、DMA*01:04 和DMB*01:02 在系统性红斑狼疮患病风险相 关[14],DMA*01:01 和DMB*01:01 在尖锐湿疣患者中的频率较高,而健康对照者中DMA*01:02在的频率较高[15]。相比于优势等位基因DMA*01:01,DMA*01:02 的rs1063478 位点碱基发生G→A 变异,导致DMA 第140 位氨基酸由谷氨酰胺突变为异亮氨酸,DMA*01:03 的rs6926628 位点碱基发生A→C 变异,导致DMA 第155 位氨基酸由甘氨酸突变为丙氨酸,在rs41555121 位点G→A变异影响下,DMA*01:03 等位基因的第184 位氨基酸由精氨酸突变为组氨酸,此位点直接影响DM 分子与DR 分子结合[16]。HLA-DMA 多态性影响DM 分子的结构稳定性和肽交换催化活性,Miguel Álvaro-Benito 等[17]发现,与单倍型DMA*01:01-DMB*01:01 相比,DMA*01:03-DMB*01:01的热稳定性下降且肽交换催化速率降低。之后该研究团队比较了DMA*01:01-DMB*01:01 和DMA*01:03-DMB*01:07,发现HLA-DM 单倍型具有特定的细胞功能,并差异性地影响T 细胞的激活,与前者相比,DMA*01:03-DMB*01:07 单倍型存在时的T 细胞活性大幅增强,同时观察到在HLADR 和HLA-DM 表达水平相同的条件下,DM 同种异型影响着形成免疫肽组的特异性组成[12]。在本研究中,未发现到DM 基因多态性与抗体水平存在相关性,推测可能是单独的DM 基因突变对T 细胞活化和浆细胞分泌抗体的影响较为局限。其他疫苗诱导抗体反应的研究发现,DMA*01:02-DMB*01:01 和DMA*01:03-DMB*01:07 单倍型与乙脑疫苗抗体非阳转组有关[18]。
综上所述,本研究表明HLA-DM 基因的多态性与脊灰疫苗诱导抗体应答存在相关性,尤其是DMA*01:02 和DMB*01:01 在脊灰Ⅰ型和Ⅱ型抗体应答阳转组和非阳转组中的差异分布可作进一步的探究。接下来可纳入更多的研究群体进行讨论,进一步分析HLA Ⅱ类抗原呈递基因多态性与疫苗诱导抗体反应之间的关联,探讨在其他疫苗和群体中DM 基因多态性影响抗体的变化方向,为新兴疫苗的研发设计提供参考。
