扁桃耐旱砧木木质部解剖结构与栓塞特性的关系分析
2024-02-22于秋红许盼云郭春苗迪利夏提·哈斯木木巴热克·阿尤普



摘"要:【目的】""基于茎木质部解剖结构与栓塞特性关系分析扁桃耐旱砧木资源对干旱胁迫环境的适应机理。
【方法】""以大巴旦(Amygdalus communis var.fragilis)(C1)、苦巴旦(A.communis var.amara)(C2)、甜仁桃巴旦[A.communis var.persicoides (West.) Rehd.](C3)和苦仁桃巴旦[A.communis var.persicoides (West.) Rehd.](C4)以及新疆毛桃(A.persica L.)(C5)等耐旱性程度不同5个扁桃砧木资源的当年生实生苗为材料,采用“外加压力”法,配套使用“XYLEM木质部导水率及栓塞测量系统”和“PMS空穴压力室”,测定在正常栽培条件下的茎自然栓塞程度以及栓塞脆弱性值(P50);测量和观察已栓塞枝条不同时段(30,80,130,180 min)的栓塞修复程度以及木质部解剖结构特征,探究耐旱性不同扁桃砧木资源的茎木质部解剖结构、栓塞抗性以及栓塞修复能力间的关系。
【结果】""(1) 扁桃不同砧木资源中大巴旦、甜仁桃巴旦、苦巴旦和普通桃的栓塞脆弱曲线为“s”形,苦仁桃巴旦的栓塞脆弱曲线为“r”形;(2) 茎木质部栓塞抗性(P50)平均值由大到小依次为大巴旦gt;甜仁桃巴旦gt;苦巴旦gt;苦仁桃巴旦gt;普通桃,其中大巴旦的抗栓塞能力最强;(3) 茎木质部栓塞修复能力由大到小依次为普通桃gt;甜仁桃巴旦gt;苦仁桃巴旦gt;苦巴旦gt;大巴旦;(4) 较耐旱砧木大巴旦、苦巴旦和甜仁桃巴旦的木质部导管及微孔直径小,木材密度、导管壁厚度及导管壁理论机械强度大,抗栓塞能力强,然而栓塞修复能力弱;‘普通桃’和苦仁桃巴旦的导管及微孔直径大,木材密度、导管壁厚度以及导管壁理论机械强度小,抗栓塞能力弱,但栓塞修复能力强。
【结论】""扁桃砧木资源木质部栓塞修复能力与栓塞抗性呈负相关,与木质部解剖结构中的导管及微孔直径呈正相关,与木材密度、导管壁厚度以及导管壁理论机械强度呈负相关。
关键词:""扁桃;砧木资源;栓塞抗性;栓塞修复;木质部解剖结构
中图分类号:"S662.9""""文献标志码:"A""""文章编号:"1001-4330(2024)11-2693-12
0"引 言
【研究意义】木质部栓塞(Xylem embolism)是一种在干旱条件下木本植物的木质部导管产生水分运输功能受损的现象,是影响植物正常生长和发育的重要制约因素[1]。植物的木质部空穴化及栓塞会使其输水功能受到影响,进而影响其耐旱能力,削弱植物防御机制,导致植物死亡[2,3]。依据Domec理论[4],木本植物木质部抵抗栓塞的能力可以用木质部导水率损失最大值的临界水势和P50值来表示[5]。在干旱地区,植物体内缺乏水分,木质部栓塞现象较为普遍[6]。耐旱性是植物感知水分缺失信号并启动应对策略的能力,在一定程度上可以由栓塞特性所决定[7-9]。研究发现,对植物重新供水后,木质部水分压力增大,导管内的气泡胀破,溶解在周围的溶液中,木质部栓塞可以得到修复[10]。栓塞修复率可以通过复水前后的导水率差值来表示,以此来描述栓塞修复能力[11,12]。植物木质部的抗栓塞能力和栓塞修复能力与其耐旱性关系密切,研究植物栓塞特性与木质部解剖结构间的关系,对于深入分析其适应干旱环境的生理生态机制的认识具有重要意义。【前人研究进展】目前木质部解剖结构与栓塞特性间的关系越来越成为研究植物耐旱性的热点[13,14]。木质部结构中的导管直径、密度、内径跨度以及纹孔结构等因素均对水分运输有重要影响[15,16]。例如木质部导管的直径越大,其内部的水分压力也就越高,当水分压力高到一定程度时,可能会导致导管内部的气泡凝聚形成气栓,从而引发栓塞[16,17]。党维等[18]在6个树种1年生枝木质部水力特征和栓塞修复能力的关系研究中发现植物木质部水力特征与栓塞修复能力有较强的关联,特别是栓塞脆弱性(P50)和水力直径是影响栓塞形成和修复的重要因素,其与导管的结构和功能密切相关;李美琪等[19]对杨树(Populus spp.)进行水分胁迫盆栽试验发现,水分胁迫程度增强从而引起木质部的水势的下降,导致木质部导水率减少,因此栓塞脆弱性降低,提高了植物抵抗栓塞的能力。【本研究切入点】扁桃(Amygdalus communis Linn.)属于蔷薇科桃属植物[20]。目前新疆扁桃最常用的砧木是普通桃,该砧木具有抗旱性较差等缺陷,对扁桃产业的可持续性影响较大,评价和筛选耐旱扁桃砧木资源一直是扁桃砧木育种的重要研究方向之一[21-23]。【拟解决的关键问题】参考以往的扁桃砧木耐旱性评价研究结果[24],选择大巴旦、苦巴旦、甜仁桃巴旦、苦仁桃巴旦和普通桃等耐旱性差异较明显的代表性扁桃砧木资源为材料,比较扁桃不同砧木资源木质部栓塞抗性、栓塞修复程度以及木质部解剖结构之间的差异,分析其与耐旱性间的关系,为扁桃耐旱砧木育种以及深入了解扁桃耐旱砧木的干旱适应策略提供理论参考。
1"材料与方法
1.1"材 料
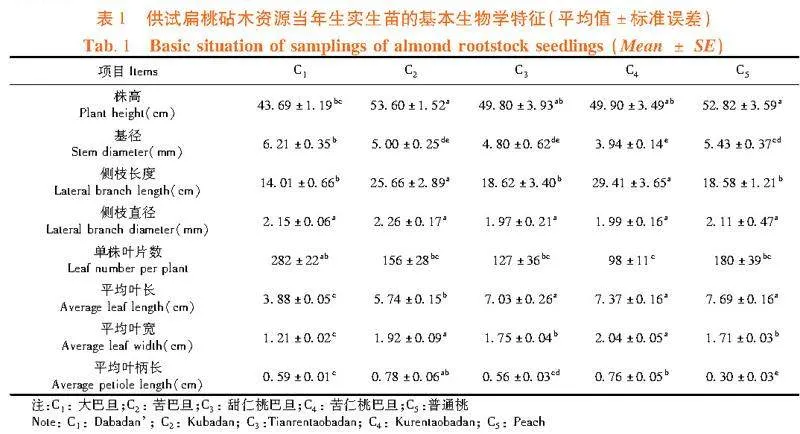
试验地选取在新疆乌鲁木齐市新疆农业科学院园艺作物研究所试验基地(43°49′13″ N, 87°34′47″ E)。该地区气候类型为中温带大陆性干旱气候,年均温4~13℃,最热月(7月)平均温度30℃,极端高温37℃,极端低温9℃;最冷月(1月)平均温度-15.2℃;年均降水量在194 mm左右。大巴旦、苦巴旦、甜仁桃巴旦、苦仁桃巴旦和普通桃种子均来自新疆喀什地区莎车县国有二林厂扁桃种质资源圃。各砧木资源分别选择100粒种子,在2022年2~4月期间室内育苗,4月初均移栽在直径25 cm、高32 cm的圆柱形花盆中,每盆1株。花盆土壤以沙土和沙壤土为主,土壤有机质含量为207.74 "g/kg,全氮含量为6.62 "g/kg,全磷含量为0.80 "g/kg,全钾含量为15.45 "g/kg,pH值为7.55。从各砧木资源选择生长健壮、长势一致的实生苗50盆,所有苗木放入四周通风通光的试验地,一直到9月中旬进行正常栽培管理。栽培期间,使所有盆栽土壤含水量达到饱和状态(达到田间持水量的75%~80%),每隔5~7 d浇1次水,每2 d测一次土壤含水量(使用SM 150T便携式土壤水分速测仪,英国,Delta公司,测定土壤电导率,参照仪器使用说明书计算土壤含水量)),使土壤含水量保持在田间最大持水量的75%~80%,保持样株生长旺盛。表1
1.2"方 法
1.2.1"栓塞脆弱性曲线的建立
试验始于2022年9月中旬,此时所有样株木质部已硬化,基本停止生长。试验当日06:00,在每个砧木资源中选取长势基本一致且主茎直径为6~8 mm的样株5株;剪取样株主茎的地上部分,在主茎上部套上黑色塑料袋,底部置于水中;在水中切除枝条基部2~3 cm的茎段,切取主茎中部长约15 cm的茎段,用“外加压力”法,配套使用“XYLEM木质部导水率及栓塞测量系统”和“PMS空穴压力室”建立栓塞脆弱性曲线[25]。
1.2.2"栓塞修复能力的测定
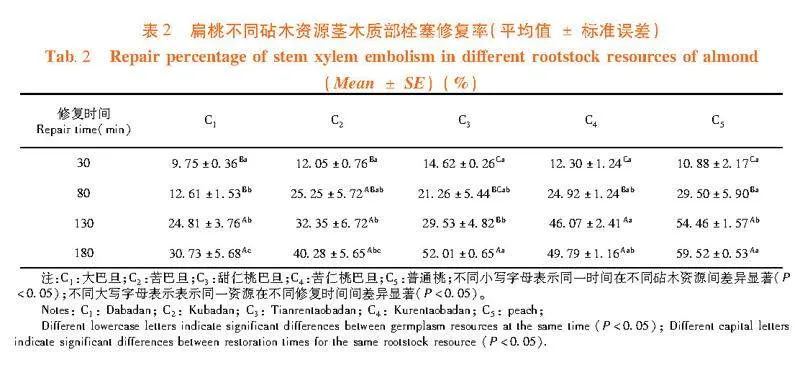
从每个砧木资源中选取长势基本一致且主茎直径为6~8 mm的样株4株,从每个样株上切取长20 cm的主茎,并对其进行人为诱导木质部栓塞,使其导水率损失值(PLC)达到95%以上。之后将茎水平放置于水中,经过30、80、130和180 min的恢复后,在靠近基部的一侧从中部截取5个连续的2 cm茎段(需在水下完成枝条的截取),随后使用“XYLEM木质部导水率及栓塞测量系统”测定其导水率的损失百分比(PLC)[6,18]。
木质部栓塞的修复能力可以通过导水率修复百分比来量化表示。
导水率修复百分比(%)=1-PLC=1-100(Kmax-Kh)/Kmax.
式中, Kh代表截段初始的导水率值,而Kmax是指截段在0.18 MPa压力下经过反复冲洗后达到的最大导水率值。对于每种扁桃砧木资源,在每个时间段内5次重复测量,取测量结果平均值表示该资源木质部栓塞的修复程度。
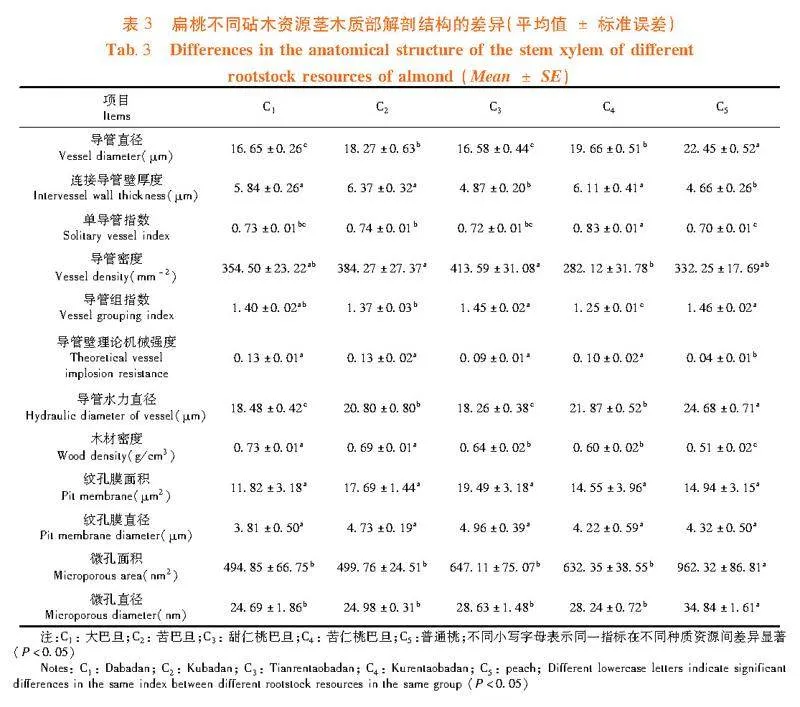
1.2.3"茎木质部解剖结构的测定
用于建立栓塞脆弱性曲线的茎段测定完成后,切取茎段中部相同位置的长度1~2 cm小段,采用番红-固绿染色法制成石蜡切片,切片厚度8~10 μm。每个扁桃砧木资源随机选取5个切片,使用Olympus BX51光学显微镜进行观察和拍照,采用Image-J图像处理软件测量木质部导管直径(Vessel diameter, D)、连接导管壁厚度(Intervessel wall thickness, TVW)、单导管指数(Solitary vessel index, VS)、导管密度(Vessel density, VD)、导管组指数(Vessel grouping index, VG)、导管壁理论机械强度(Theoretical vessel implosion resistance, (TVW/D连接导管)2)、导管水力直径(Hydraulic diameter of vessel, Dh)和木材密度(Wood density, WD)[26]。在剩余茎段中部切取长度5-10 mm的小段,参考Jansen等[27]方法在不同浓度梯度的乙醇溶液中脱水,于室内自然风干,将样品纵切后固定在样品架上,喷白金3 min;采用Zeiss Supar 55 (VP)扫描电子显微镜(德国,Carl Zeiss公司)在2 kV电压下观察纹孔膜结构并拍照,采用Image-J图像处理软件测量纹孔膜面积(Pit membrane area, Pa)、纹孔膜直径(Pit membrane diameter, Pd)、微孔面积(Microporous area, Ma)和微孔直径(Microporous diameter, Md)。
1.3"数据处理
利用SPSS 19.0 (SPSS Inc., Chicago, IL, USA)和Origin 2022 (OriginLab Corporation, Northampton, MA, USA)分析数据和绘图,分析前需满足正态分布和方差齐性。采用单因素方差分析自然栓塞度(PLC)、栓塞脆弱性(P50)、栓塞修复能力以及木质部解剖结构在扁桃不同砧木资源间的差异,采用LSD法进行差异显著性分析,Plt;0.05表示差异性显著。
2"结果与分析
2.1"扁桃砧木资源茎木质部栓塞抗性的差异
2.1.1"茎栓塞脆弱性曲线的建立
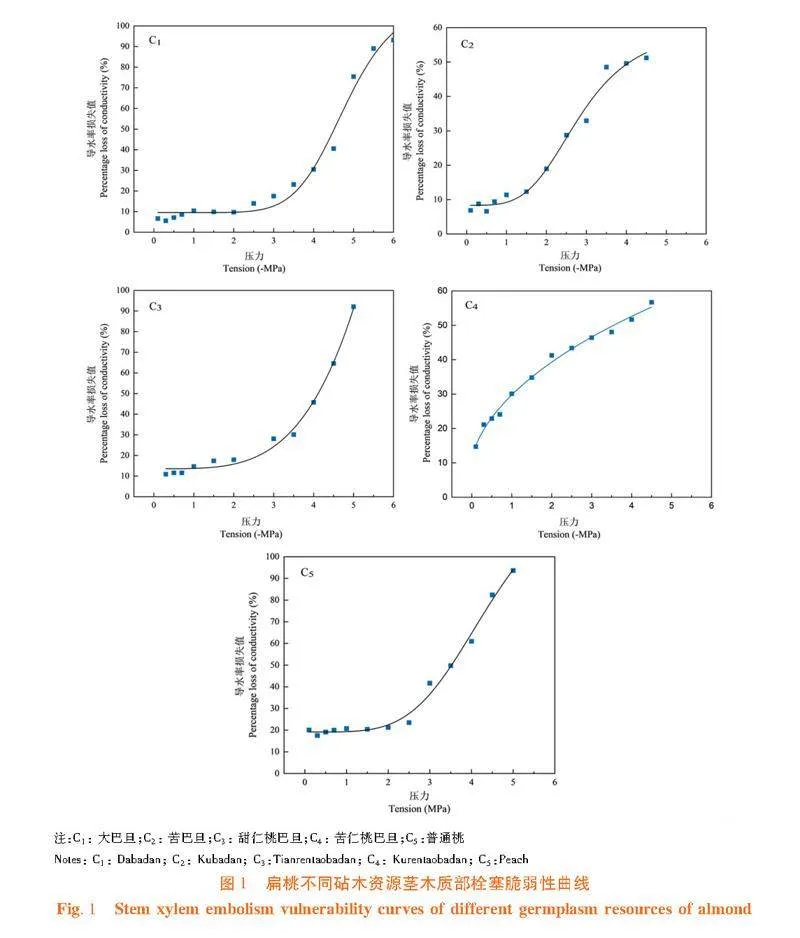
研究表明,苦仁桃巴旦的茎对木质部栓塞较敏感,在木质部压力出现负值的初始阶段,枝条木质部导水率损失值迅速增大,且随着水势的不断降低,木质部导水率损失百分数(PLC%)即栓塞化程度迅速增大,呈典型的“"r"”形。在木质部负压为-3.55 MPa时,其导水率损失值约为50%,达到最大导水率损失值时的木质部压力约为-6 MPa。大巴旦和苦巴旦茎木质部栓塞脆弱曲线呈典型的“S”形,当木质部压力大于-2.0 MPa时,其导水率损失值很小,木质部负压达到-2.0 MPa时,导水率损失值迅速增大,当导水率损失值达到最大时,木质部压力约为-5~-6 MPa。苦巴旦茎木质部压力大于-1.5 MPa时,导水率损失值几乎不变,但当压力处于-4.0~-1.5 MPa时,木质部导水率损失值迅速增大,木质部发生较大程度的栓塞。甜仁桃巴旦和普通桃的茎木质部压力大于-2 MPa时,导水率损失值几乎不变,但当压力处于-5.0~-2.0 MPa时,木质部导水率损失值迅速增大,木质部发生较大程度的栓塞。大巴旦的茎木质部压力大于-3.0 MPa时,导水率损失值几乎不变,仅发生较小程度的栓塞,当压力处于-5.5~-3.0 MPa时,木质部导水率损失值迅速增大,木质部发生较大程度栓塞。图1"
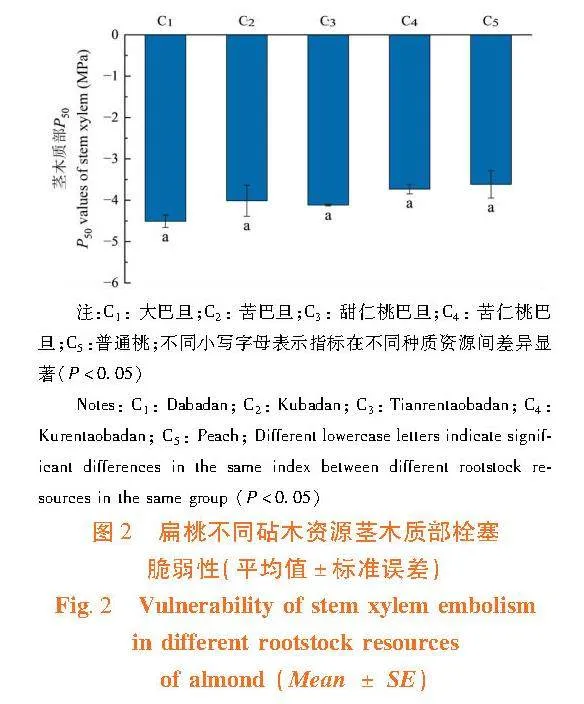
2.1.2"茎木质部栓塞抗性的差异
研究表明,扁桃不同砧木资源茎栓塞脆弱性之间差异不显著(Pgt;0.05)。各资源茎木质部栓塞脆弱性程度高到低为:普通桃(3.62 ± 0.33)gt;苦仁桃巴旦(3.73 ± 0.11)gt;苦巴旦(4.01 ± 0.38)gt;甜仁桃巴旦(4.12 ± 0.02)gt;大巴旦(4.51 ± 0.15) 。植物木质部的栓塞脆弱性大,栓塞抗性小,在水分运输过程中植物木质部易发生栓塞,对干旱胁迫的抵御能力差。因此,所有供试资源中普通桃的抗栓塞能力最弱,大巴旦的抗栓塞能力最强。图2
2.2"扁桃砧木资源茎木质部栓塞修复能力差异
研究表明,在栓塞修复初期30 min时,甜仁桃巴旦的栓塞修复程度相对较好,而大巴旦的栓塞修复程度相对较慢。在栓塞修复中期80 min时,普通桃的栓塞修复程度相对较好,大巴旦的栓塞修复程度相对较慢;在栓塞修复进行130 min时,普通桃和苦仁桃巴旦的栓塞修复程度均较好。除普通桃和苦仁桃巴旦外,其余3个砧木资源在修复130 min时的栓塞修复程度差异不显著;在栓塞修复后期180 min时,栓塞修复程度最好的是普通桃,而大巴旦的修复程度最弱。在栓塞修复180 min时,“s”形曲线的大巴旦、苦巴旦、甜仁桃巴旦、普通桃和“r”形脆弱性曲线的苦仁桃巴旦之间栓塞修复程度存在显著性差异(Plt;0.05)。表2
木质部栓塞的修复程度逐渐提高的趋势。在修复30 min时,甜仁桃巴旦的修复程度大于"苦仁桃巴旦、苦巴旦和普通桃,但在修复"80~130 min时甜仁桃巴旦的修复程度要低于苦仁桃巴旦、苦巴旦和普通桃,甜仁桃巴旦在短期内修复程度较优,而苦仁桃巴旦、苦巴旦和普通桃在中期修复程度较甜仁桃巴旦优;在修复130~180 min时甜仁桃巴旦的修复程度要高于苦仁桃巴旦和苦巴旦,甜仁桃巴旦在长时间内修复程度较苦仁桃巴旦和苦巴旦要优;大巴旦在各个时间段其栓塞修复程度均较弱,显著低于其它砧木资源。
扁桃各砧木资源在180 min时的栓塞修复程度间差异显著(Plt;0.05),其木质部栓塞修复能力强到弱为普通桃gt;甜仁桃巴旦gt;苦仁桃巴旦gt;苦巴旦gt;大巴旦。即植物重新供水后,木质部栓塞修复能力大的砧木资源,其栓塞导管能迅速恢复其输水功能。图3
2.3"扁桃砧木资源茎木质部栓塞特性与解剖结构间的关系"
2.3.1"茎木质部解剖结构的差异
研究表明,茎木质部解剖结构的各项指标在扁桃砧木资源间均呈显著性差异,扁桃种质资源植株的茎木质部栓塞脆弱性曲线可以用单威布尔函数很好地拟合,并且苦仁桃巴旦的栓塞脆弱性"曲线呈典型的“"r"”形,大巴旦和苦巴旦为典型的“S”形曲线,这与前人研究提出的两种栓塞脆弱性曲线类型相符lt;0.05) 。其中,大巴旦和苦巴旦的(TVW/D连接导管)2均显著高于其它砧木资源;甜仁桃巴旦的VD显著高于其它砧木资源(Plt;0.05),但其D和Dh均显著低于其它扁桃种质资源;苦仁桃巴旦的VD和VG均显著低于其它扁桃种质资源,而VS较高;普通桃除了D、VG和Dh均显著高于其它砧木资源外,其它各项茎木质部解剖结构指标均显著低于其它砧木资源(Plt;0.05)。表3,图4
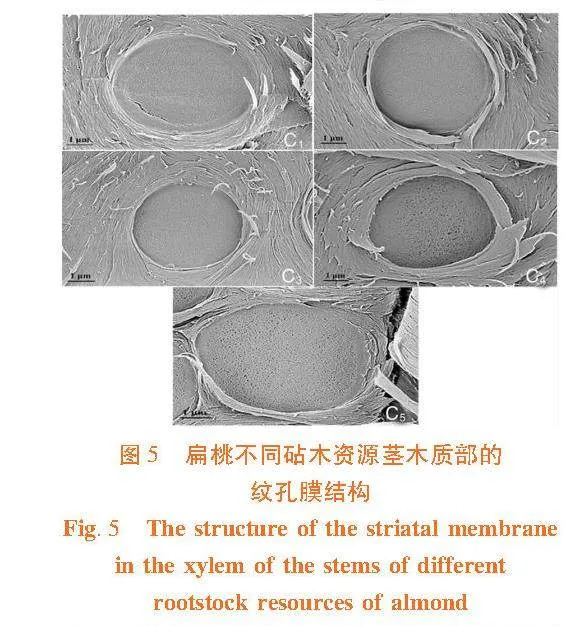
各扁桃砧木资源的茎纹孔膜上均有微孔,微孔的大小在各扁桃砧木资源间具有一定差异。普通桃的Ma和Md显著高于其它砧木资源,大巴旦显著低于其它资源(Plt;0.05)。而Pa和Pd在各砧木资源间差异不显著。图5
2.3.2"茎木质部栓塞抗性、栓塞修复与解剖结构间的内在关系"
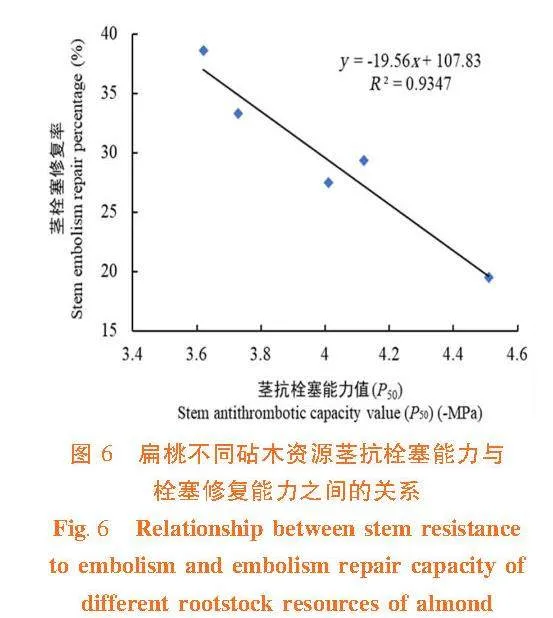
研究表明,扁桃不同砧木资源茎木质部抗栓塞能力(P50)、栓塞修复能力、木材密度和木质部解剖结构之间有较好的相关性。不同砧木茎木质部抗栓塞能力与栓塞修复能力之间具有负相关关系(R2 = 0.934 7),表现为抗栓塞能力强,不易发生栓塞,则栓塞修复能力弱;大巴旦的抗栓塞能力强,却栓塞修复能力弱,普通桃的抗栓塞能力弱,但栓塞修复能力强。图6
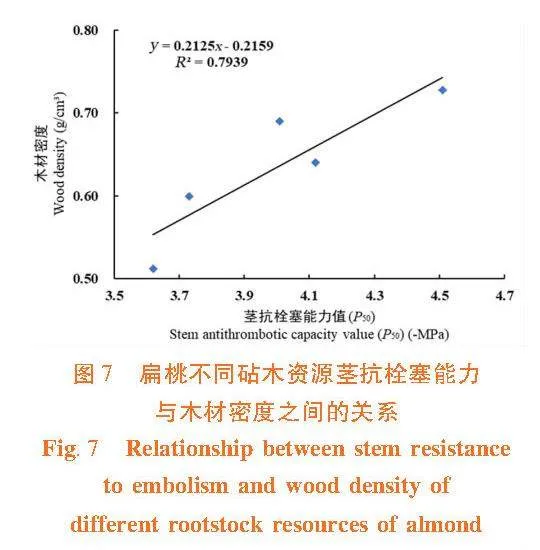
茎木质部抗栓塞能力与木材密度之间呈显著正相关关系(R2 = 0.793 9) 。木材密度大,栓塞抗性强,植物不易发生栓塞。栓塞抗性强的大巴旦和苦巴旦木材密度明显高于栓塞抗性弱的甜仁桃巴旦、苦仁桃巴旦和普通桃。并且,茎木质部栓塞修复能力与木材密度间呈负相关关系(R2 = 0.929 7)。由此可见,木材密度是木质部抗栓塞能力和栓塞修复能力权衡关系的调节因子。大巴旦和苦巴旦的木材密度大,抗栓塞能力强,栓塞修复能力弱;普通桃的抗栓塞能力最弱,但它的木材密度小,具有较高的储水能力,有利于栓塞的木质部恢复输水功能。图7,图8"
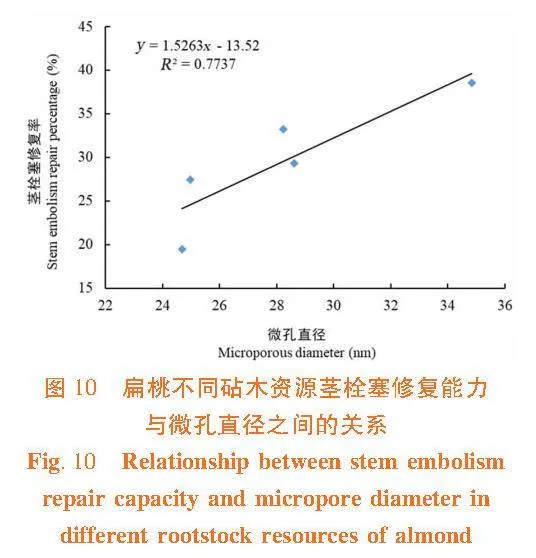
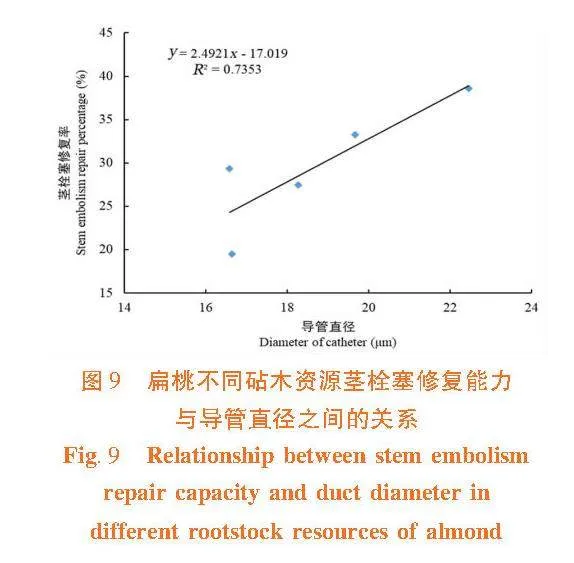
茎木质部栓塞修复能力与木质部导管直径和纹孔膜上的微孔直径间呈正相关关系(R2 = 0.735 3,R2 = 0.773 7),茎木质部导管直径和纹孔膜上的微孔直径越大,在栓塞修复过程中空气容易排出或溶解于周围溶液中,则栓塞越容易修复。 大巴旦的抗栓塞能力在5个砧木资源中最强,导管直径和纹孔膜上的微孔直径最小,栓塞修复能力最弱,在缺水环境下大巴旦茎木质部不容易发生栓塞,保持一定的导水能力,但一旦发生栓塞,其不易修复。普通桃的抗栓塞能力最弱,导管直径和纹孔膜上的微孔直径最大,栓塞修复能力最强。图9,图10
3"讨 论
3.1"栓塞抗性与木质部解剖结构间的关系
扁桃种质资源植株的茎木质部栓塞脆弱性曲线可以用单威布尔函数很好地拟合,并且苦仁桃巴旦的栓塞脆弱性曲线呈典型的“"r"”形,大巴旦和苦巴旦为典型的“S”形曲线,这与前人研究提出的2种栓塞脆弱性曲线类型相符[28,29]。建立栓塞脆弱性曲线可以更好地了解植物栓塞抵抗力的特征和变化,并进一步研究树木耐旱性和抵御干旱等环境压力的能力[30,31]。扁桃砧木资源的木质部解剖结构与其栓塞抗性间存在一定的相关性。首先,普通桃和苦仁桃巴旦的直径D较其它扁桃砧木资源大,且其与P50值相关性较高,D越大,易发生栓塞,栓塞抗性越小,与前人研究中大径导管植物比小径导管植物更容易发生栓塞的结论相似[32,33]。普通桃和苦仁桃巴旦之所以容易发生栓塞也可能与前人研究提出的气种假说有关,导管的纹孔膜透性以及纹孔膜上微孔的大小,尤其是在干旱等胁迫条件下,对植物的耐旱性具有至关重要的影响[34,35]。随着D的增加,纹孔占据的面积相应增大,纹孔膜上出现较大微孔的概率也随之提高,降低了植物对栓塞的抵抗力。在变化的水分环境和生长条件下,树木通过调整自身形态和结构来协调导水效率与栓塞抗性,从而确保其生长发育的成功,特别是在干旱或不稳定环境中尤为关键[36]。
除导管直径外,栓塞抗性还与(TVW/D连接导管)2有关。有研究表明,(TVW/D连接导管)2与抗栓塞能力有一定的关系,(TVW/D连接导管)2较大的植物可以减小水势负压对木质部导管壁的破坏[37]。Cochard等[30]对蔷薇科李属植物研究得出相同结论,导管壁理论机械强度越大,P50值与其相关性越好。纹孔膜厚度和导管壁厚度较大的植物具有较高的木材密度和力学性质,这是因为较厚的导管壁可以增加木材的强度和抗压能力,而纹孔膜的厚度则可以减少木材的孔隙率,从而增加木材的密度[14,27]。木巴热克等[26]对扁桃引进品种和新疆本地品种的茎木质部解剖结构和栓塞抗性关系进行研究,发现其木材密度和栓塞抗性间有较强的相关性,木材密度越大,栓塞抗性越强,植物不易发生栓塞。在研究中,栓塞抗性较强的大巴旦和苦巴旦的WD和(TVW/D连接导管)2大,栓塞抗性强,可以更好的承受水势负压对其导管壁的破坏。"
3.2"栓塞修复与木质部解剖结构间的关系
植物木质部内部相邻导管间水势是否存在一定的差值是决定栓塞可否消除的前提,水势差高时栓塞易修复[6]。邻近部位或区域的水势越高,栓塞修复越好。在修复过程中,茎木质部输水结构尤其是纹孔膜结构特征,会影响所需水势梯度的大小[11]。研究中,栓塞修复能力与导管直径以及纹孔膜上微孔直径呈正相关,导管直径和微孔直径小,木质部抗栓塞能力强,栓塞修复能力弱。
Ogasa等[8]的研究结果表明,木材密度和栓塞抗性与修复能力之间存在负相关关系。这表明,木材密度越高,植物的抗空穴化能力强,但栓塞的修复能力可能会降低。因此,木材密度低的植物可能会在栓塞修复方面表现更加出色。当受到干旱胁迫时,高密度的木材可能会为植物提供更好的保护,可以更好地防止空穴的发生。然而,当植物需要频繁修复栓塞时,低密度的木材可能更有利于栓塞的修复。研究发现,栓塞修复能力与木材密度呈负相关,木材密度较小的普通桃,栓塞抗性较弱,但其栓塞修复能力强,以此适应长期干旱的环境。
3.3"栓塞抗性与栓塞修复间的关系
栓塞抗性较高的植物能够有效地防止栓塞的发生,从而减少了栓塞修复的需求[6]。刘丽等[38]研究发现,84K杨(P.abla × P.glandulousa ‘84K’)的木质部导管直径越大,其抗栓塞能力越弱,自然栓塞度越高,栓塞修复能力越强;党维等[18]在6个树种的1年生枝条中,木质部的水力特性与栓塞修复能力有着密切的联系。尤其是栓塞脆弱性(P50)和水力直径,是影响栓塞形成与修复的重要因素,与导管的结构和功能紧密相关;安锋等[39]对8种木本植物木质部栓塞抗性、脆弱性和修复能力进行了测试和比较,结果表明,栓塞脆弱性越大,植物组织的破坏程度越大,反映出该植物对于环境的适应性较低,重新补水后,栓塞修复速率越快,意味着在干旱环境中具备更好的适应性和修复能力。在扁桃不同砧木资源中,普通桃的栓塞抗性最弱,P50最大,更容易发生栓塞;而大巴旦较耐旱,抗栓塞能力强,但栓塞后的修复能力弱。
4"结 论
扁桃5个砧木资源中,耐旱性较强大巴旦资源的茎木质部导管直径以及微孔直径小、木材密度大,栓塞抗性强,但栓塞修复能力弱;而缺水较敏感砧木普通桃的茎木质部导管直径以及微孔直径大、木材密度小,栓塞抗性弱,但栓塞修复能力强。
参考文献"(References)
[1]"木巴热克·阿尤普, 伊丽米努尔, 荆卫民.不同水分处理下几种柽柳属植物幼株木质部栓塞及其解剖结构特征[J].北京林业大学学报, 2017, 39(10): 42-52.Mubareke Ayoupu, Yiliminuer, JING Weimin.Xylem anatomical features and native xylem embolism of several Tamarix spp.species seedlings under different water treatments[J].Journal of Beijing Forestry University, 2017, 39(10): 42-52.
[2] McDowell N G, Beerling D J, Breshears D D, et al.The interdependence of mechanisms underlying climate-driven vegetation mortality[J].Trends in Ecology amp; Evolution, 2011, 26(10): 523-532.
[3] Nardini A, Battistuzzo M, Savi T.Shoot desiccation and hydraulic failure in temperate woody angiosperms during an extreme summer drought[J].New Phytologist, 2013, 200(2): 322-329.
[4] Domec J C, Gartner B L.Cavitation and water storage capacity in Bole xylem segments of mature and young Douglas-fir trees[J].Trees, 2001, 15(4): 204-214.
[5] Choat B, Jansen S, Brodribb T J, et al.Global convergence in the vulnerability of forests to drought[J].Nature, 2012, 491(7426): 752-755.
[6] 党维.耐旱树种木质部栓塞恢复状况的研究[D].杨凌: 西北农林科技大学, 2016.DANG Wei.The Refilling of Embolized Xylem of Drought-tolerant Tree Species[D].Yangling: Northwest A amp; F University, 2016.
[7] Awad H, Barigah T, Badel E, et al.Poplar vulnerability to xylem cavitation acclimates to drier soil conditions[J].Physiologia Plantarum, 2010, 139(3): 280-288.
[8] Ogasa M, Miki N H, Murakami Y, et al.Recovery performance in xylem hydraulic conductivity is correlated with cavitation resistance for temperate deciduous tree species[J].Tree Physiology, 2013, 33(4): 335-344.
[9] Rolland V, Bergstrom D M, Lenné T, et al.Easy come, easy go: capillary forces enable rapid refilling of embolized primary xylem vessels[J].Plant Physiology, 2015, 168(4): 1636-1647.
[10] Love D M, Sperry J S.In situ embolism induction reveals vessel refilling in a natural aspen stand[J].Tree Physiology, 2018, 38(7): 1006-1015.
[11] 孟凤.七种槭树科植物木质部栓塞及其恢复与植物抗旱性的关系[D].杨凌: 西北农林科技大学, 2019.MENG Feng."The Relationship between Xylem Embolism and Embolism Recovery and Plant Drought Resistance in Seven Genus Acer[D].Yangling: Northwest A amp; F University, 2019.
[12] McCulloh K A, Meinzer F C.Further evidence that some plants can lose and regain hydraulic function daily[J].Tree Physiology, 2015, 35(7): 691-693.
[13] Hacke U G, Jansen S.Embolism resistance of three boreal conifer species varies with pit structure[J].The New Phytologist, 2009, 182(3): 675-686.
[14] Plavcová L, Hacke U G, Sperry J S.Linking irradiance-induced changes in pit membrane ultrastructure with xylem vulnerability to cavitation[J].Plant, Cell amp; Environment, 2011, 34(3): 501-513.
[15] Levionnois S, Kaack L, Heuret P, et al.Pit characters determine drought-induced embolism resistance of leaf xylem across 18 Neotropical tree species[J].Plant Physiology, 2022, 190(1): 371-386.
[16] Avila R T, Kane C N, Batz T A, et al.The relative area of vessels in xylem correlates with stem embolism resistance within and between Genera[J].Tree Physiology, 2023, 43(1): 75-87.
[17] Christensen-Dalsgaard K K, Tyree M T.Frost fatigue and spring recovery of xylem vessels in three diffuse-porous trees in situ[J].Plant, Cell amp; Environment, 2014, 37(5): 1074-1085.
[18] 党维, 姜在民, 李荣, 等.6个树种1年生枝木质部的水力特征及与栓塞修复能力的关系[J].林业科学, 2017, 53(3): 49-59.DANG Wei, JIANG Zaimin, LI Rong, et al.Relationship between hydraulic traits and refilling of embolism in the xylem of one-year-old twigs of six tree species[J].Scientia Silvae Sinicae, 2017, 53(3): 49-59.
[19] 李美琦, 姜在民, 赵涵, 等.加杨水力学与生理特性对不同土壤水分条件响应研究[J].植物生理学报, 2017, 53(4): 632-640.LI Meiqi, JIANG Zaimin, ZHAO Han, et al.Study on the adaptability of hydraulic and physiological characteristics to different soil moisture conditions in Populus x canadensis Moench[J].Plant Physiology Journal, 2017, 53(4): 632-640.
[20]梁艳霞, 张亚楠, 王占和.扁桃薄壳品种‘蒙特瑞’在山西中部地区的引种表现及栽培技术[J].果树资源学报, 2022, 3(6): 88-90.LIANG Yanxia, ZHANG Yanan, WANG Zhanhe.Introduction performance and cultivation technology of the thin-shelled variety 'Monterey' of lentil in central Shanxi[J].Journal of Fruit Tree Resource, 2022, 3(6): 88-90.
[21] 侯江涛, 张毅芳, 沈聪聪, 等.扁桃引种栽培技术研究综述[J].林业科技通讯, 2020,(2): 11-14.HOU Jiangtao, ZHANG Yifang, SHEN Congcong, et al.Review on introduction and cultivation techniques of Amygdalus communis[J].Forest Science and Technology, 2020,(2): 11-14.
[22] 梁艳霞, 王占和, 张亚楠, 等.扁桃特性及其丰产栽培技术[J].黑龙江粮食, 2021,(10): 108-109.LIANG Yanxia, WANG Zhanhe, ZHANG Yanan, et al.Characteristics of almond and its high-yield cultivation techniques[J].Heilongjiang Grain, 2021,(10): 108-109.
[23] 任哲, 贡翔, 张锐, 等.扁桃优株叶片解剖结构与其抗旱性关系研究[J].农业与技术, 2022, 42(3): 10-14.REN Zhe, GONG Xiang, ZHANG Rui, et al.扁桃优株叶片解剖结构与其抗旱性关系研究[J].Agriculture and Technology, 2022, 42(3): 10-14.
[24] Ayup M, Yang B, Gong P, et al.Evaluation of drought resistance of native almond-rootstock varieties in Xinjiang, China[J].New Zealand Journal of Crop and Horticultural Science, 2022, 50(1): 48-68.
[25] 李荣.耐旱树种木质部结构与耐旱性关系研究[D].杨凌: 西北农林科技大学, 2016.LI Rong.Relationships between Xylem Strucrure And Drought Tolerance of Drought Tolerant Tree Species[D].Yangling: Northwest A amp; F University, 2016.
[26] 木巴热克·阿尤普, 杨波, 艾沙江·买买提, 等.基于当年生枝木质部解剖结构的扁桃品种栓塞抗性分析[J].西北林学院学报, 2021, 36(5): 99-105.Rebareke Ayoupu, YANG Bo, Ashajiang Maimaiti, et al.Xylem embolism resistance of different AlmondCultivars based on the xylem anatomical characteristics of current-year shoot[J].Journal of Northwest Forestry University, 2021, 36(5): 99-105.
[27] Jansen S, Choat B, Pletsers A.Morphological variation of intervessel pit membranes and implications to xylem function in angiosperms[J].American Journal of Botany, 2009, 96(2): 409-419.
[28] Torres-Ruiz J M, Jansen S, Choat B, et al.Direct X-ray microtomography observation confirms the induction of embolism upon xylem cutting under tension[J].Plant Physiology, 2015, 167(1): 40-43.
[29] Sperry J S, Christman M A, Torres-Ruiz J M, et al.Vulnerability curves by centrifugation: is there an open vessel artefact, and are ‘r’ shaped curves necessarily invalid?[J].Plant, Cell amp; Environment, 2012, 35(3): 601-610.
[30] Cochard H, Barigah S T, Kleinhentz M, et al.Is xylem cavitation resistance a relevant criterion for screening drought resistance among Prunus species[J].Journal of Plant Physiology, 2008, 165(9): 976-982.
[31] Tyree M T, Engelbrecht B M J, Vargas G, et al.Desiccation tolerance of five tropical seedlings in Panama.relationship to a field assessment of drought performance[J].Plant Physiology, 2003, 132(3): 1439-1447.
[32] Cai J, Zhang S X, Tyree M T.A computational algorithm addressing how vessel length might depend on vessel diameter[J].Plant, Cell amp; Environment, 2010: 33(7): 1234-1238.
[33] Markesteijn L, Poorter L, Paz H, et al.Ecological differentiation in xylem cavitation resistance is associated with stem and leaf structural traits[J].Plant, Cell amp; Environment, 2011, 34(1): 137-148.
[34] Wheeler J K, Sperry J S, Hacke U G, et al.Inter-vessel pitting and cavitation in woody Rosaceae and other vesselled plants: a basis for a safety versus efficiency trade-off in xylem transport[J].Plant, Cell amp; Environment, 2005, 28(6): 800-812.
[35] Cochard H, Bréda N, Granier A, et al.Vulnerability to air embolism of three European oak species (Quercus petraea (Matt) Liebl, Q pubescens Willd, Q robur L)[J].Annales Des Sciences Forestières, 1992, 49(3): 225-233.
[36] Johnson D M, McCulloh K A, Woodruff D R, et al.Hydraulic safety margins and embolism reversal in stems and leaves: Why are conifers and angiosperms so different[J].Plant Science, 2012, 195: 48-53.
[37] Hacke U G, Sperry J S, Pockman W T, et al.Trends in wood density and structure are linked to prevention of xylem implosion by negative pressure[J].Oecologia, 2001, 126(4): 457-461.
[38] 刘丽, 张立, 蔡靖, 等.干旱胁迫及复水后84K杨栓塞修复及其他水力学特性的研究[J].北京林业大学学报, 2021, 43(7): 22-30.LIU Li, ZHANG Li, CAI Jing, et al.Hydraulic characteristics and embolism repair of Populus alba × P.glandulosa after drought stress and rehydration[J].Journal of Beijing Forestry University, 2021, 43(7): 22-30.
[39] 安锋, 蔡靖, 姜在民, 等.八种木本植物木质部栓塞恢复特性及其与PV曲线水分参数的关系[J].西北农林科技大学学报(自然科学版), 2006, 34(1): 38-44.AN Feng, CAI Jing, JIANG Zaimin, et al.Refilling of embolism in the xylem of eight tree species and its relationship with Pressure-Volume parameters[J].Journal of Northwest Sci-Tech University of Agriculture and Forestry (Natural Science Edition), 2006, 34(1): 38-44.
Relationship between the anatomical structure of xylem"""and embolization characteristics of drought tolerant rootstocks of almond
YU Qiuhong1, XU Panyun2, GUO Chunmiao2, Dilixiati Hasimu1, Mubareke Ayoupu1
(1. College of Life Sciences, Xinjiang Agricultural University, Urumqi 830052, China; 2. Institute of Horticultural Crops, Xinjiang Academy of Agricultural Sciences, Urumqi 830091, China)
Abstract:【Objective】 ""To analyze the adaptation mechanism of drought resistant rootstocks of almond to the drought stress environment from the relationship between the stem xylem anatomical structure and the embolism characteristics.
【Methods】 ""The one-year old seedlings of five different almond rootstocks with different drought tolerance levels, including Da Badan (Amygdalus communes var.fragilis)(C1), Ku Badan ( A.communs var.amara)(C2), Tianren Taobadan [A.communs var."persistides (West.) Rehd.](C3), Kuren Taobadan [A.communs var.persistides (West.) Rehd.](C4), and Xinjiang local peach (A.persista L.)(C5), were selected as research materials.Using the \"external pressure\" method, combined with the \"XYLEM xylem hydraulic conductivity and embolism measurement system\" and \"PMS pressure chamber\", the degree of stem natural embolism level and embolism vulnerability value (P50) were measured under normal cultivation conditions.The embolism repairment of the embolized branches at different time periods (30, 80, 130, 180 min) and the xylem anatomical characteristics were analyzed to explore the relationship between the anatomical structure of the stem xylem, the embolism resistance, and the embolism repairment of different drought resistant almond rootstocks.
【Results】 ""(1) The vulnerability curves(CV) of Da Badan, TianRen Taobadan, Ku Badan, and local peaches were in an \"S\" shape, while the CV curves of Kuren Taobadan were in an \"R\" shape; (2) The order of stem xylem embolism resistance (P50) was Da Badangt; TianRen Taobadangt; Ku Badangt; Kuren Taobadangt;Local Peach, with Da Badan having the most embolism resistant xylem; (3) The order of xylem embolism repair ability was as follows: Local peachgt; TianRen Taobadangt; Kuren Taobadangt; Ku Badan gt; Da Badan; (4) The drought resistant rootstocks as Da Badan, Ku Badan , and TianRen Taobadan all had small vessels and micropores, high wood density, thick conduit walls and theoretical mechanical strength of conduit walls.They had relative strong embolism resistant ability, but were weaker to embolism repairment.Local peaches and Kuren Taobadan had larger vessels and micropores, lower wood density, and its vessel wall thickness, and theoretical mechanical strength of the vessel wall were smaller than other rootstocks.
【Conclusion】 ""In summary, the xylem embolism resistance is negatively correlated with embolism repair ability in different almond rootstocks, and the drought resistant rootstocks have embolism resistant xylem.The xylem embolism repairment is positively correlated with the diameter of vessel and micropores, and negatively correlated with wood density, vessel wall thickness, and theoretical mechanical strength of the vessel wall in xylem.
Key words:""almond; rootstocks; embolism resistance; embolism repairment; xylem anatomical structure
Fund projects:""Key research and development program of the Xinjiang Uygur Autonomous Region (2024B02018-1);Basic Scientific R amp;D Program of Public Welfare Research Institutions of Xinjiang Uygur Autonomous Region(KY2020110)
Correspondence author:"""Mubareke Ayoupu (1981-), female, from Shache,Xinjiang, associate researcher, doctorate, research direction: the physiological ecology of fruit tree adversity, (E-mail) mubarek@ms.xjb.ac.cn
收稿日期(Received):
2024-04-19
基金项目:
新疆维吾尔自治区重点研发计划项目(2024B02018-1);新疆维吾尔自治区公益性科研院所基本科研业务经费项目(KY2020110)
作者简介:
于秋红(1998-),女,河北沧州人,硕士研究生,研究方向为植物逆境生理,(E-mail)2074276722@qq.com
通讯作者:
木巴热克·阿尤普(1981-),女,新疆莎车人,副研究员,博士,研究方向为果树逆境生理生态,(E-mail)mubarek@ms.xjb.ac.cn
