基于大脑海马认知机理的主从式AUV 协同定位方法
2024-02-16黄鸿殿孔庆宇奔粤阳
李 倩,聂 简,黄鸿殿,孔庆宇,奔粤阳
(哈尔滨工程大学 智能科学与工程学院,哈尔滨 150001)
在海洋科学研究与军事打击任务中,单体自主水下航行器(Autonomous Underwater Vehicle,AUV)受限于搭载传感器的种类、感知范围以及信息处理能力,往往难以满足复杂任务需求。与单体AUV 不同,AUV 集群协同作业能够以更高的效率执行更加复杂的分布式任务,而且更适应复杂动态环境,因而受到广泛关注[1]。针对AUV 集群编队,基于艇间信息交换与共享实现AUV 协同定位是提高编队整体导航定位性能的主要技术途径。在主从式协同定位系统中,部分主AUV 搭载高精度惯性导航系统或惯性基组合导航系统,其余从AUV 装备低精度航位推算系统。主从AUV 通过水声通信设备共享定位信息并获取艇间相对距离量测信息,从而利用数据融合技术估计并校正从AUV 定位误差。通常情况下,水下复杂环境会导致艇间水声测距量测信息含有大量野值[2],进而导致AUV 协同定位系统量测噪声一般不满足高斯分布假设,从而对AUV 协同定位算法的鲁棒性提出了更高需求。
目前,AUV 协同定位主流核心算法大都基于贝叶斯估计框架,而这其中卡尔曼滤波(Kalman Filter,KF)是协同定位算法中最常见也是工程上最易实现的方法之一。由于AUV 协同定位系统量测模型具有非线性,所以协同定位算法中KF 通常以其非线性扩展形式出现,即扩展卡尔曼滤波(Extended Kalman Filter,EKF)[3]。除此之外,无迹卡尔曼滤波(Unscented Kalman Filter,UKF)、容积卡尔曼滤波(Cubature Kalman Filter,CKF)也被应用于协同定位从而达到减小非线性量测模型线性化误差的目的[4,5]。但是,上述协同定位算法在面对非高斯噪声时,存在导航状态估计精度下降、甚至滤波失效等问题[6]。粒子滤波(Particle Filter,PF)虽然能够解决非高斯噪声估计问题,但需要大量粒子才能较好地近似系统后验概率密度,因此其巨大的计算量限制了该算法在实际工程中的应用[7-9]。针对AUV 协同定位非高斯噪声问题,文献[10]提出一种基于泰勒级数展开的扩展Student'st鲁棒卡尔曼滤波算法(Student’s t Extended Kalman Filter,SEKF),解决AUV 协同定位过程中水声测距噪声非高斯分布问题。文献[11]将基于改进广义最大似然估计的卡尔曼滤波与交互式多模型算法结合,解决AUV 协同导航过程中量测噪声非高斯分布与时变问题。文献[12]采用统计方法对磁罗盘、GPS、水声定位与通信设备的输出进行野值判别与剔除。当输出被判定为野值时,EKF只进行时间更新而不进行量测更新。值得注意的是,上述方法都是在传统卡尔曼滤波框架下进行算法鲁棒优化设计。
在探索更加高效导航方法的过程中,研究人员发现哺乳动物不需要高精度的感官信息就能确定自身在复杂环境中所处的位置[13]。通过模仿哺乳动物大脑海马体空间细胞导航机理,研究人员提出了模仿生物大脑的类脑导航方法,并将类脑导航机理应用于视觉SLAM 领域[14]。通过建立空间表征细胞模型,编码多种定位信息模拟神经细胞放电表征环境位置变化,进而完成导航定位[15]。昆士兰科技大学Milford 教授团队建立的RatSLAM[16]系统利用局部视图细胞和位姿细胞,使用吸引子网络建立细胞模型,通过视觉模板信息对视觉里程计误差进行校正,从而准确表达机器人位置信息。但是,RatSLAM 主要基于纯视觉信息用于导航定位。近年来,研究人员逐渐开始利用多种大脑导航细胞模型实现多源导航信息融合。文献[17]提出的NeuroBayes-SLAM 使用头朝向细胞和网格细胞实现角度编码与位置编码,并基于贝叶斯理论将视觉信息与运动信息融合到细胞模型编码中。北京工业大学于乃功团队提出了一种包含头朝向细胞、条纹细胞、网格细胞、位置细胞的机器人建图方法[18,19]。利用视觉信息对路径积分误差进行校正,并验证了网格细胞积分对于不合法或者突变数据具有一定的滤波整合效果,相比于纯路径积分拥有更高的定位精度。
受大脑海马体不同种类细胞模型之间信息交互机制的启发,本文提出一种基于海马体细胞模型的主从式AUV 协同定位方法。首先,基于二维吸引子网络模型构建网格细胞模型,并利用其对航向信息与速度信息进行路径积分。在此基础上,利用竞争性Hebb学习使多尺度的网格细胞生成具有单个放电活动包的位置细胞。最后,将AUV 主从艇之间的水声测距量测信息转化为位置细胞放电变化,并通过位置细胞多次放电更新后的累积变化逐渐逼近真实位置,降低量测野值对AUV 位置更新的影响,从而实现对从艇航位推算累积误差的抑制。
1 基于水声测距的协同定位系统模型
本文采用双领航艇主从式AUV 协同定位配置方式。考虑到AUV 普遍配备高精度深度传感器,且其深度误差不随时间累积,因此将AUV 三维定位问题简化为二维定位问题。选取东-北-天地理坐标系作为导航坐标系,从AUV 在x-y水平面上的离散运动学方程如式(1)所示:
其中,Xk=[xk,yk]T表示k时刻AUV 东向与北向位置,控制输入可以通过AUV 所搭载的多普勒计程仪或电磁计程仪与罗经系统测量得到。实际工程中,传感器输出均受到噪声干扰,因此可以表示为:
其中,wk=[wv,k,wθ,k]T表示系统噪声。
系统量测信息Zk=[z1,k,z2,k]T是AUV主从艇之间的距离,可以表示为:
其中,z1,k,z2,k分别表示从AUV 与主AUV1、主AUV2 之间的距离量侧信息,可由水声设备获得。X1,k=[x1,k,y1,k]T、X2,k=[x2,k,y2,k]T分别表示主AUV1 与主AUV2 的位置信息,vk=[ν1,k,ν2,k]T表示水声测距量测噪声。
根据式(1)~式(3),基于水声测距量测信息的AUV 协同定位系统模型可以表示为:
其中,f(·) 表示非线性状态方程函数,h(·) 表示非线性量测方程函数。
在基于海马体细胞模型的AUV 协同定位算法中,首先利用网格细胞完成路径积分,即实现如式(1)所示的Xk时间更新。进一步,将路径整合信息传递给位置细胞,通过位置细胞放电获得AUV 实际位置。为校正从AUV 航位推算误差,获取AUV 艇间测距信息Zk并将其转换为位置细胞放电输入,从而对位置细胞进行校正,即实现如式(3)所示的量测更新。同时,为维持位置细胞稳定放电过程,通过全局兴奋与全局抑制将放电活动包限制在有效范围内。最后,从AUV 经协同校正后的位置信息可以根据校正后的位置细胞放电活动计算得出。基于海马体细胞模型的AUV 协同定位算法具体原理框图如图1 所示。

图1 基于海马体的AUV 协同定位原理Fig.1 Principle of cooperative localization for multiple AUVs based on hippocampal formation
2 基于大脑海马细胞的协同定位算法
2.1 基于网格细胞的航位推算
2005 年,Hafting 等在大鼠脑海马体中发现网格细胞,研究表明其在生物活动时主要起路径积分作用[20]。通过构建合适的数学模型模拟动物大脑网格细胞作用机理,可以实现基于子艇航向与速度信息的路径积分。本文采用Bruak 提出的二维吸引子模型对网格细胞进行建模[21]。吸引子模型中的细胞相互连接,通过合理的激励、抑制与连接机制,可以驱动细胞网络状态向偏移方向连续移动,形成稳定的六边形放电活动包,从而实现路径积分。
采用二维吸引子网络模型进行建模,网格细胞在x-y水平面上排列形成Nx×Ny的二维细胞板。其中,Nx、Ny分别表示细胞板x、y轴向排列的细胞数量,如图2(a)所示。由于细胞板的长度与宽度有限,为解决吸引子边界问题,将细胞板上下两侧的细胞与左右两侧的细胞相连接,从而组成如图2(b)所示的二维环状模型结构。
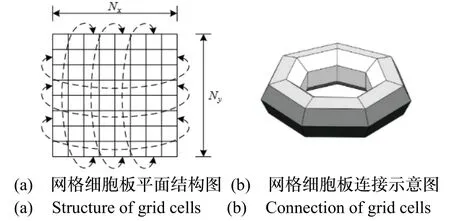
图2 二维连续吸引子网络模型Fig.2 2D continuous attractor network model
网格细胞的放电由吸引子动力学等式给出,即:
其中,τ表示网格细胞神经元相应的时间常量;si表示Nx×Ny个吸引子细胞中第i个网格细胞的放电率;神经元传递函数f[·] 是一个简单的非线性整流函数,即当x>0 时,f(x)=x;当x≤ 0时,f(x)=0。
Bi用于输入优先方向的速度信息,即:
其中,eθi表示指向网格细胞i优先朝向的单位向量,v表示速度向量,可由AUV 控制输入中的速度经由航向分解获得。实际应用中网格细胞的优先朝向是任意值,为方便程序编码,将网格细胞优先方向设定为北、南、东、西四个方向,分别对应0、π/2、π、3/2π,在网格细胞板中2×2 的区域内均匀排布,如图3 所示。
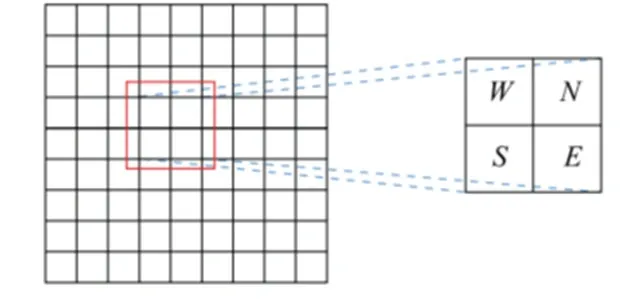
图3 网格细胞的优先朝向Fig.3 Preferred direction of grid cells
根据图3 可知,每个网格细胞都与周围其它网格细胞相互连接,从而接收其它网格细胞的信息输入。权值Wij表示网格细胞j到网格细胞i之间的连接权值,具体表达式为:
式(8)中,a、γ、β均为权值调节参数,由经验值设定,一般取表示网格周期,可以用来调节生成网格图案的大小。表示编号为i的网格细胞在细胞板上的横纵坐标位置向量。式(7)中的l与式(6)中的α决定了速度输入的驱动强度。根据式(7)可知,权值矩阵分布呈以为中心的墨西哥帽形状分布,如图4(a)所示,通过调节a可以调节细胞网络中单个网格细胞激励或抑制周围网格细胞放电。特别地,当a=1时,权值都为负,即网格细胞之间的连接都是抑制性的,如图4(b)所示。
通过上述网格细胞神经元网络的激励与抑制作用,可以促使网格细胞放电图形向真实方向移动,从而表达AUV 的位移信息,如图5 所示。
需要注意的是,单一网格细胞板放电对位置信息的表达具有歧义。因此,需要通过改变λnet生成具有不同尺度的网格细胞板,从而使用多个网格细胞板联合解码得到精确位置信息,不同尺度网格细胞如图6所示。
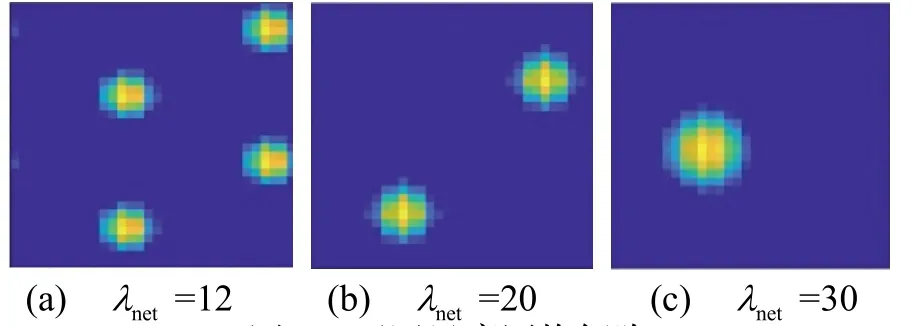
图6 不同尺度网格细胞Fig.6 Grid cells with different scales
在获得多尺度网格细胞的基础上,进一步通过由网格细胞到位置细胞的放电即可实现对真实位置的表达。位置细胞于1971 年被O’ Keefe 等人在对鼠脑海马体研究的过程中发现[13],该细胞主要起到环境认知的作用,一个位置细胞神经元和物理空间中某一具体位置有着对应关系。生物学上位置细胞的放电受到多种感官信息的影响,而网格细胞放电就是位置细胞的信息输入来源之一。多个不同尺度的网格细胞板放电作为前向输入,使位置细胞生成单一的放电活动包,从而表征AUV 位置信息。
为了处理不同尺度网格细胞的输入,使用竞争性Hebb 网络更新不同尺度网格细胞到位置细胞的权值,即:
其中,wn表示第n层网格细胞板到位置细胞板的连接权值,K表示学习率,pi表示位置细胞放电率,表示第n层网格细胞板中第i个网格细胞的放电率,Ig表示学习阈值,一般由的平均值确定。位置细胞的放电率可以进一步表示为:
其中,A表示增益系数,位置细胞放电抑制量Ip=B× max(pi)用来控制位置细胞放电活动包的个数,参数B越大,位置细胞放电范围越大,M表示网格细胞神经板的层数。
为了保证Hebb 竞争性网络稳定的学习与后续位置信息解码,每次时间更新都对位置细胞放电进行归一化处理,即:
通过上述Hebb 竞争性算法可以确保只有少量网格细胞族的子集被选择形成位置细胞,从而避免网格细胞位置表达的歧义性。由含有多个放电活动包的网格细胞生成具有单一放电活动包的位置细胞,如图7所示。

图7 位置细胞放电活动Fig.7 Firing activity of place cells
需要注意的是,此时得到的位置细胞板同网格细胞板一样,具有二维环状模型结构,即上下两边、左右两边分别连通。当位置细胞放电活动包跨过边界时,需要根据环状连接结构进行循环。利用网格细胞放电对位置细胞放电更新,再对位置细胞放电进行解码,进而得到位置细胞板放电中心位置,即:
其中,xi
p
,k是k时刻位置细胞i在位置细胞板上的位置向量,pi,k是k时刻位置细胞i的放电率。
进一步,经过边界修正与坐标系转换以后可以得到k时刻位置细胞所对应的物理空间具体位置,即:
其中,Xk表示AUV 位置,C表示位置细胞坐标板坐标系到导航坐标系的转换矩阵;Dk=[εxNx,εyNy]T+d,其中εx,εy表示位置细胞经过边界的次数,d用来校正初始位置。当位置细胞经过细胞板边界时,则重新分割二维环状细胞板模型,从而使位置细胞放电包处于细胞板中心,并基于分割位置重新计算AUV 位置。
2.2 基于位置细胞放电的协同校正
利用艇间测距信息作为量测信息,可以实现从AUV 航位推算累积误差的协同校正。在基于大脑海马认知机理的SLAM系统中,大多利用视觉模板匹配得到的位置信息对位置细胞放电直接进行校正,从而获得更加精确的位置信息。考虑到AUV 协同定位系统中的艇间测距量测信息并不是对从艇位置的直接观测,本文利用主从艇的位置信息与距离量测信息构建“新息”,进一步将该“新息”转化为位置细胞的放电活动,从而通过放电更新不断逼近位置细胞准确位置,获得更精确的AUV 位置信息。
首先,选择以位置细胞放电活动包为中心的位置细胞板周围区域组成一个新的Mx×My的校正位置细胞板。校正位置细胞板初始均匀放电,即放电率为:
其中,pm表示第m个细胞的放电率,m=1,2…Mx×My。
当从AUV 接收到与两个主AUV 之间的距离量测信息z1、z2以及主AUV 位置信息以后,更新位置细胞放电率为:
其中,δ表示控制放电范围的参数,通常设置为常数;Ic表示校正位置细胞板放电抑制常量,l1、l2表示新息,其表达式为:
其中,xm、ym表示第m个位置细胞对应的导航坐标系位置,可基于式(13)由位置细胞坐标系到导航坐标系转换得出;x1、y1与x2、y2分别表示主AUV1 与主AUV2 的位置。
每次获取量测信息以后,校正位置细胞板放电也随之更新,并逐渐汇聚到真实位置附近。因此,位置细胞的放电包含多次量测信息,从而可以有效降低单次量测野值对于位置细胞放电的负面影响。需要注意的是,随着多次放电更新,位置细胞的放电变化会逐渐减小甚至停止,如图8 所示为第1、4、7 次校正位置细胞板的放电活动。

图8 校正位置细胞板放电活动Fig.8 Firing activity of correction place cells
为了维持细胞板稳定放电,当细胞放电集中在小范围以后,本文采用一种全局兴奋与全局抑制放电规则将放电活动包限制在有效范围内。位置细胞间的兴奋性权值连接矩阵由一个二维高斯分布表示:
同理,位置细胞的抑制性权值连接矩阵可以表示为:
其中,kr2与ar2< 0为常量。
通过全局兴奋与全局抑制,校正位置细胞板上的放电活动范围在不断接收新量测信息的情况下仍能处于稳定状态。校正位置细胞板更新后,利用高斯分布重新建立位置细胞模型表示为:
其中,pi表示校正后的位置细胞放电,是第i个位置细胞在位置细胞板坐标系的横纵位置坐标,x'=[x',y']T是更新后的位置细胞中心,δp控制位置细胞放电范围的大小。
3 仿真实验验证
为验证本文所提出的基于大脑海马认知机理的AUV 协同定位算法性能,将其与基于扩展卡尔曼滤波的协同定位算法[3]进行对比。仿真实验中设置参数如下:网格细胞板与位置细胞板的横纵细胞数量Nx=Ny=32;使用40 个不同的网格细胞板,为了生成不同尺度的网格细胞,网格周期从12 到32 之间间隔为0.5 均匀分布采样;W0调节参数a=1.0315,学习率k为0.005,抑制性系数B=0.5;在更新的位置细胞板中,选择位置细胞放电活动包中心周围16 m×16 m 作为校正区域,划分成32×32 的细胞板,位置细胞放电更新的阈值设定为0.9。
仿真实验过程中,系统噪声满足高斯白噪声分布,即ωk~ N (0,Q)。为模拟水声测距数据中的异常野值,将量测噪声设置为Student’st分布,根据式(20)生成:
其中,Q=diag([0.1 m/s,1 °]2) 表示系统噪声协方差矩阵,R=diag([2 m,2 m]2)表示量测噪声协方差矩阵。根据式(20)产生的量测噪声如图9 所示。图9中,黑色菱形标记了测量噪声中存在的野值。

图9 主从AUV 距离测量噪声Fig.9 Range measurement noise between leader AUV and slave AUV
为提高系统可观测性,令从AUV 航向角按照模型θ=π/9 · sin(3π ·k/1000)+π/4变化,从而进行‘S’形机动航行,首尾向速度v=0.5m/s,仿真时间1000 s。考虑到量测噪声含有随机野值,同时与带有阈值判断的EKF 协同定位算法进行对比,主、从AUV 航行轨迹以及从AUV 估计轨迹如图10 所示。
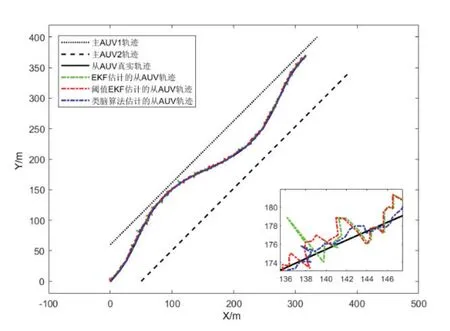
图10 AUV 轨迹示意图Fig.10 Trajectory of AUVs
为进一步定量评估算法性能,定义协同定位误差LE、平均定位误差ALE 与均方根误差RMSE 如下:
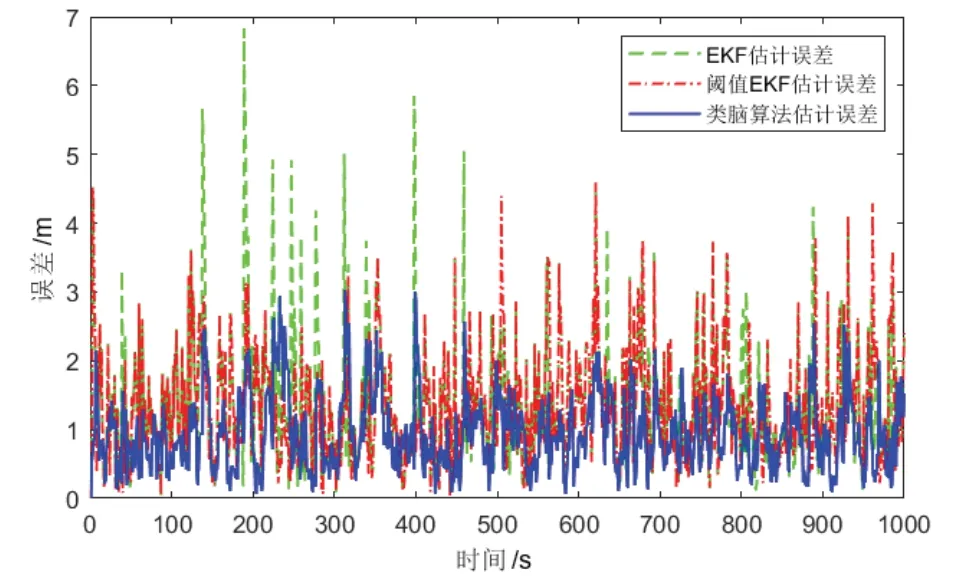
图11 从AUV 定位误差Fig.11 Localization errors of slave AUV
不同协同定位算法位置误差如表1 所示。
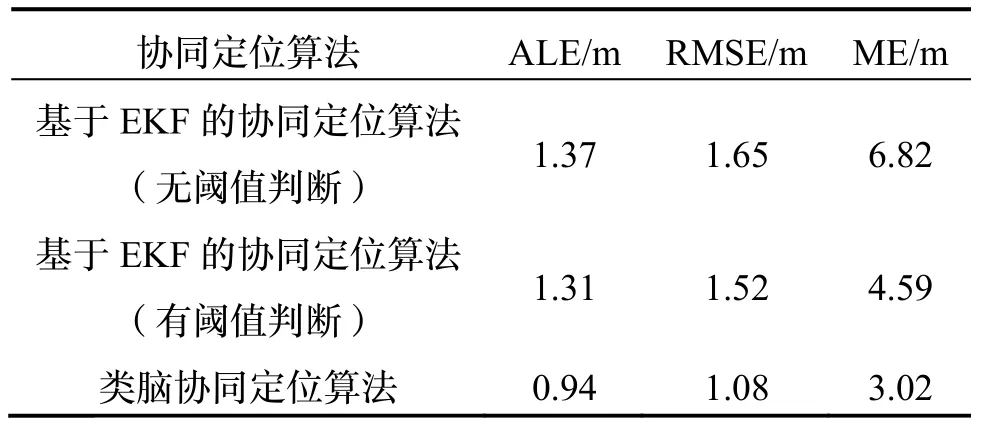
表1 不同协同定位算法位置误差Tab.1 Localization errors with different cooperative localization algorithms
根据表1 可以看出,在量测噪声含有野值的情况下,类脑协同定位算法的最大定位误差(ME)与均方根误差(RMSE)都远小于EKF 类协同定位算法。这是由于类脑协同定位算法量测更新并非通过对新息逐个加权直接实现,而是经过多次细胞放电更新逐渐逼近真实位置,从而在一定程度上避免单一量测野值对细胞放电产生影响。与带有阈值判断的EKF 协同定位算法相比,类脑协同定位算法定位误差最大值可以降低34%,平均定位误差可以降低28%。
4 结论
本文基于哺乳动物大脑海马体空间细胞导航机理,提出一种主从式AUV 协同定位算法。通过构建二维吸引子网格细胞模型实现从AUV 速度与航向信息的积分推算,同时,利用主从艇位置信息与距离量测信息构建“新息”,并将该“新息”转化为位置细胞的放电活动,从而通过放电更新不断逼近位置细胞准确位置,获得更为精确的AUV 位置信息。本文所提协同定位算法可以在非高斯量测噪声条件下有效降低艇间测距噪声中所含有的大量野值对于系统定位精度的影响,从而提高AUV 协同定位系统在水下复杂环境中的适应能力。
