Effects of drought on non-structural carbohydrates and C,N,and P stoichiometric characteristics of Pinus yunnanensis seedlings
2024-01-26ZhijuanZhaoLinaWangYuanxiLiuJianliSunJiandongXiaoQiongDongLianfangLiWanjieZhangChaoWangJunwenWu
Zhijuan Zhao · Lina Wang · Yuanxi Liu · Jianli Sun · Jiandong Xiao ·Qiong Dong · Lianfang Li · Wanjie Zhang · Chao Wang · Junwen Wu
Abstract To study non-structural carbohydrate characteristics and nutrient utilization strategies of Pinus yunnanensis under continuous drought conditions,2-year-old seedlings were planted in pots with appropriate water,light and moderate and severe drought treatments [(80 ± 5),(65 ± 5),(50 ± 5),and (35 ± 5)% of field water-holding capacity].Non-structural carbohydrates,carbon (C),nitrogen (N),and phosphorus (P) concentrations were measured in each plant component.The results show that: (1) With increasing drought,non-structural carbohydrates gradually increased in leaves,stems,and coarse roots,while gradually decreased in fine roots;(2) C concentrations of all were relatively stable under different stress levels.Phosphorous utilization of each component increased under light and moderate drought conditions,while N and P utilization efficiency of each plant component decreased under severe drought.Growth was mainly restricted by N,first decreasing and then increasing with increased drought;(3) There was a correlation between the levels of non-structural carbohydrates and C,N,and P in each component.Changes in N concentration affected the interconversion between soluble sugar and starch,which play a regulatory role in the fluctuation of the concentration of non-structural carbohydrates;and,(4) Plasticity analysis showed that P. yunnanensis seedlings responded to drought mainly by altering starch concentration,the ratio of soluble sugar to starch in leaves and stems,and further by altering N and P utilization efficiencies.Overall,these results suggest that the physiological activities of all organs of P.yunnanensis seedlings are restricted under drought and that trade-offs exist between different physiological indicators and organs.Our findings are helpful in understanding nonstructural carbohydrate and nutrient adaptation mechanisms under drought in P. yunnanensis seedlings.
Keywords Pinus yunnanensis seedlings · Drought ·Non-structural carbohydrates · C,N,P stoichiometric characteristics
Introduction
Drought is the main factor restricting plant growth,as it hinders photosynthesis,respiration,stomatal movement,and it affects plant development and physiological metabolic activities.In addition,it changes the distribution of substances and nutrients within the plant (Hartmann and Trumbore 2016;Sainju et al.2017).With global climate changes,the survival and growth of the world’s forests are under rising threats of drought due to the increasing intensity,frequency,and duration of drought events,coupled with rising temperatures.Tree drought mortality in the background of global warming is already a hot issue,and an increase in the number of dead trees in a forest will eventually lead to changes in the structure and function of the forests (Allen et al.2015).Winter–spring drought conditions are a major climate threat in Yunnan Province,and their impact on forest growth is increasing,in addition,regeneration and survival are severely affected (Shen et al.2020).
Non-structural carbohydrates (NSC) are important products of photosynthesis that not only participate in structure building but also support metabolic processes as important energy substances (Körner 2003).Non-structural carbohydrates storage in trees is closely related to their growth mechanisms and plays an important role in their adaptation to environmental changes (Dietze et al.2014).Nonstructural carbohydrates are composed of soluble sugar and starch,and its concentration characterizes the status of carbon acquisition,utilization,and carbon balance in plants,as well as their resistance to stress (Raessler et al.2010).When plants are exposed to long-term stress,the balance between carbon uptake and expenditure is disrupted.When carbon synthesis is insufficient to meet the requirements of metabolism,NSC concentrations decrease to a threshold level,“carbon starvation”occurs,which eventually leads to plant death (McDowell 2011).Soluble sugar and starch transform each other due to changes in environmental factors.For example,in a long-term water shortage environment,starch transforms into soluble sugar,increasing the osmotic adjustment ability of cells to resist drought.Therefore,variations of NSC concentrations are an important feature of trees for coping with drought (Yang et al.2016).Studies on the effects of drought on NSCs in Japanese cypress (Chamaecyparis obtusa(Siebold &Zucc.) Endl.) showed that starch levels in photosynthetic organs decreased at the beginning of drought while soluble sugar increased.With longer periods and intensities of drought,starch and soluble sugar levels are reduced (Tsuji et al.2022).However,under drought,the concentration of soluble sugar and NSCs in each organ ofBetula ermaniiseedlings increased significantly,but starch levels decreased,which is a response to the water deficit by increasing the transformation between starch and soluble sugar (Ma et al.2021).InRobinia pseudoacaciaL.seedlings under severe drought,starch and NSC concentrations were low,while soluble sugar and the proportion of soluble sugar to starch were high (Zhang et al.2015).In addition,drought affected NSCs transportation and distribution in different organs (Hartmann et al.2013).Current research on the trade-off between tree NSCs storage and growth are inconsistent.In addition to depending on tree species,it is also related to ontogeny (Myers and Kitajima 2007) and type of stress (Sara et al.2014).At present,there is no consensus on the impact of drought on the NSCs of seedlings.
Carbon (C),nitrogen (N),and phosphorus (P) are important elements involved in various activities of plants and play important roles in growth and metabolism (Wang et al.2018).The ecological stoichiometric ratio is one of the most effective indicators for determining the nutrient status of the environment for growth,and the C,N,and P concentrations in different plant organs and their stoichiometric ratios reflect their ecological adaptations in response to environmental changes (Moe et al.2005).Water is the basis of a series of physiological and biochemical reactions in plants.When water supply is insufficient,C,N,and P concentrations and their stoichiometric characteristics change in trees(Sardans and Penuelas 2012).In addition,C,N,and P levels and their ratios differ among tree components,and nutrient uptake responds differently to water supply (Brant and Chen 2015).Less soil moisture reduces the availability of nutrients and limits their absorption by plants (Ali et al.2016).N uptake efficiency usually increases with decreasing water supply (Vergutz et al.2012),while P levels in senescing leaves significantly decreases (Tang et al.2013).N/P increased in leaves of loess plateau plants (Zheng and Shangguan 2007) and N concentration in leaves of plants in sandy,semi-arid areas increased (Wei et al.2011).Wang et al.(2019) studiedCaragana microphyllaLam.under drought and found that C concentrations in fine and coarse roots gradually increased,while P levels in fine roots first increased and then decreased.This indicates that the absorption capacity of roots of different diameters varies,depending on water and nutrient responses to drought.Therefore,it will be beneficial to study changes in C,N,and P concentrations and their distribution within plants under different degrees of drought.
Pinus yunnanensisFranch.is a deep-rooted species that is sun-loving,highly adaptable and tolerant of dry climates and poor soils.As an endemic species in southwest China,it is a priority species for the reforestation of barren sites on the Yunnan–Guizhou Plateau and also a major forest ecosystem species in Yunnan Province.It has an irreplaceable role in the sustainable economic and environmental development of Yunnan forestry.With global warming,drought has become an important constraint on the growth and community development of forests,especially in the seedling or juvenile stage,where plants have a shallow root systems compared to mature trees and suffer more from water shortages under drought.Seedling mortality increases exponentially with decreasing precipitation or water availability (Cavender-Bares and Bazzaz 2000;Marod et al.2004).Under drought conditions,the basal diameters ofP.yunnanensisseedlings were smaller significantly and the biomass of each organ and the total biomass were less compared to controls (Gao et al.2020).Recently,researchers have carried out numerous studies on the effects of drought on NSCs and C,N,and P stoichiometric characteristics,typically including the effects of drought on the NSCs ofRobinia pseudoacaciaL.(Zhang et al.2015),Acer rubrum,Betula papyrifera,Fraxinus americana,Quercus rubra,Quercus velutina(Maguire and Kobe 2015),Ulmus minorMill.,Quercus ilexL.(Rodrguez-Calcerrada et al.2016),Pinus massoniana(Lin et al.2018),Pinus sylvestris,andPicea abies(Ivanov et al.2018),and on the stoichiometric characteristics ofMedicago sativa(Lu et al.2019),Pinus massoniana(Lin et al.2020),Fagus sylvaticasaplings (Zang et al.2021),andQuercus ilex(Sardans et al.2020).However,there have been no studies on NSCs and ecological stoichiometry forP.yunnanensisseedlings.The objectives of this study were to: (1) determine any changes in NSCs and C,N,and P stoichiometric characteristics at the organ level ofP.yunnanensisseedlings under drought and (2) analyze the phenotypic plasticity of physiological indicators ofP.yunnanensisseedlings under drought conditions.This study aimed to enhance the understanding of the physiological mechanisms of drought resistance and to provide a basis for nutrient cycling studies and water management forP.yunnanensisseedlings.
Materials and methods
Overview of the study area
The study was carried out in the greenhouse of the Arboretum of Southwest Forestry University (Kunming,Yunnan,102° 46′ E,25° 03′ N),located in the monsoon climate zone of the subtropical plateau,with an altitude of 1964 m,short frost period,and mild climate.The annual average temperature is 16.5 °C,annual average precipitation 1035 mm,and annual average relative humidity 67%.Temperatures in the greenhouse were (18.5–34.0 °C),relative air humidity(22.3–48.0%),and light transmittance 80%.
Test materials and treatments
Two-year-oldP.yunnanensisseedlings were cultivated on a Yiliang garden forest farm,and 120 seedlings with robust growth and uniform height were selected and transplanted into 9.23 L flowerpots.The soil used was a local red loam mixed with humus at a volume ratio of 3:2,and field waterholding capacity 25.9%.Its bulk weight was 1.14 g cm-3,and its total C,total N,and total P concentration were 3.26,5.98,and 0.62 g kg-1,respectively;pH was 6.6,as determined using the ring-knife method.The seedlings were placed at the experimental site,covered with mulch to prevent underground moisture from affecting the potted plants,and a shelter built to ensure good ventilation and no shade.The shelter was set up on rainy days to prevent rain from affecting the potted plants.The shelter was made of transparent material that protected the plants from the rain without affecting light and ventilation.
The moisture level of the seedlings was maintained at field capacity after planting to ensure healthy growth.Potting simulated natural drought conditions,with four moisture gradients,i.e.,suitable moisture (CK),light drought(LD),moderate drought (MD),and severe drought (SD).Soil moisture concentration of each gradient was set at(80 ± 5%),(65 ± 5%),(50 ± 5%) and (35 ± 5%) of field waterholding capacity,i.e.,the actual moisture concentration was maintained at 19.4–22.0%,15.6–18.2%,11.7–17.3%,and 7.8–10.4%,respectively.Different treatments ensured a uniform experimental environment throughout;the soil moisture levels of the potted plants was controlled manually.Seedlings that survived were numbered and divided into the four soil moisture conditions.Differences in heights and basal diameters indicated that the groups were evenly formed.When there were no differences in heights and basal diameters of the four groups,a tag was hung to mark the corresponding number.Heights and basal diameters of all surviving seedlings were recorded,and the water supply was then limited to the soil water concentration falling within the predetermined range.Seedling weights were recorded and samples taken as background values.The experiment started March 14,2021 and ended May 12,2021,a total of 60 days.Actual soil water concentrations were measured with a soil moisture meter and controlled using the weighing method,where all pots were weighed daily at 17:00,the weight was noted,and water control or watering performed according to the target weight.
Sampling and measurement
Sampling was carried out at the end of the experiment with four plants taken from each treatment,a total of 16 plants.Plants were divided into four parts: leaves,stems,coarse roots >2-mm diameter,and fine roots Excel 2013 and SPSS19.0 were used to process the data.The data were tested for normality and homogeneity of variance prior to analysis of variance.Two-way analysis of variance(ANOVA) tested the effects of drought intensity on plant organs,and interactions on NSCs levels and their components,and on C,N,and P concentrations and their ratios.One-way ANOVA and Duncan’s multiple comparisons tested the significance of the different leaf,stem,coarse root,and fine root components under different drought gradients(α=0.05).Graphpad Prism 8.0 and Origin 8.0 were used to plot the graphs.Pearson’s method analyzed the correlation of various indexes among different organs. whereXmaxandXminrepresent the maximum and minimum values of each index. The effects of different drought intensities and their interactions on NSCs concentrations and their components are shown in Table 1.There were significant differences in soluble sugar,starch,NSCs,and ratio of soluble sugar to starch among organs (P<0.01).Drought intensity highly affected soluble starch (SS) (P<0.01) and significantly non-structural carbohydrates concentrations (P<0.05).The effects on starch and ratio were not significant (P>0.05).The interaction of drought and organs had a significant effect on soluble sugar and starch concentration (P<0.05) and a highly significant effect on ratio (P<0.01),while there was no significant effect on NSCs concentration (P>0.05). Table 1 Two-factor analysis of variance (ANOVA) on the effect of drought on non-structural carbohydrates in different organs of Pinus yunnanensis seedlings As shown in Fig.1,with the increase in drought intensity,soluble sugar and NSCs concentration in leaves,stems,and coarse roots ofP.yunnanensisseedlings showed an increasing trend,but gradually decreased in fine roots,and starch concentration in fine roots showed an increasing trend.The soluble sugar concentration of leaves,stems,and coarse roots under severe drought was significantly higher than that of the CK (P<0.05),and concentrations increased by 27.1%,30.9%,and 35.0%,respectively,while the soluble sugar levels in fine roots was reduced by 22.6% under the SD treatment compared to the controls.Non-structural carbohydrates concentrations of leaves,coarse roots,and fine roots was not significantly different among treatments (P>0.05),while the NSCs concentration of stems was higher than CK in moderate and severe drought conditions,rising by 34.1% and 40.0%,respectively.Under LD,the starch concentration of the leaves increased significantly (P<0.05)by 40.3% compared to CK.The starch concentration of stems under MD and SD also increased (P<0.05) by 65.3% and 68.0%,respectively,while the starch concentration in the coarse roots showed no significant difference among treatments (P>0.05).Under LD,the soluble sugar and starch concentration of leaves decreased significantly (P<0.05) by 27.4%.Under MD,the ratio of fine roots decreased by 36.7%.Under SD,the ratio in leaves increased by 41.9%,while it in stems and fine roots decreased significantly (P<0.05) by 37.8% and 31.3%,respectively. Fig.1 Accumulation and distribution of non-structural carbohydrates in Pinus yunnanensis seedlings under different drought treatments;CK (suitable moisture),LD(light drought),MD (moderate drought),SD (severe drought).NSC: non-structural carbohydrate,SS: soluble sugar,ST: starch.Different capitals indicate the same degree of drought with significant differences among different organs(P <0.05),and lowercase letters indicate the same organ with significant differences in degrees of drought (P <0.05) Under different degrees of drought,soluble sugar levels were highest in the stem and least in the fine roots;the soluble sugar concentration in fine roots was lower than that of other organs under SD treatment (P<0.05).Starch concentrations in stems was significantly higher than in fine roots under all treatments (P<0.05).The NSCs concentration of fine roots was significantly lower than that of other organs under each treatment (P<0.05),and the ratio of leaves was higher than that of stems and fine roots under SD treatment (P<0.05). The two-way ANOVA (Table 2) show highly significant differences (P<0.01) in C and N concentrations and C:N,C:P,and N:P ratios among the different organs,while the difference in P was insignificant.Drought had no effect on C concentration but was highly significant on N and P levels and C:N,C:P,and N:P ratios (P<0.01).The interaction between organs and drought only had a significant effect on C:P (P<0.05). Table 2 ANOVA on the effect of drought on C,N,and P concentrations and their stoichiometric ratios in different organs of Pinus yunnanensis seedlings There were no significant differences in C concentrations in leaves,stems,and coarse roots among treatments (Fig.2).The C concentration of fine roots under LD was significantly higher than that of the controls (P<0.05) and increased by 26.4%.Nitrogen concentrations increased with the greater drought,N of leaves and fine roots was higher than that of stems and coarse roots under all treatments.Leaf nitrogen levels were not significantly different from the controls under LD and SD (P>0.05),while they increased significantly by 42.4% under MD.Nitrogen levels of stems,coarse roots,and fine roots did not change significantly under LD and MD (P>0.05),while they increased significantly under SD(P<0.05).Nitrogen levels of coarse and fine roots increased 82.6% and 69.2%,respectively,compared to the controls.The N concentration of stems was 2.7-times higher that of CK.Compared with the controls,phosphorous levels significantly decreased in leaves and stems under LD and MD(P<0.05),by 54.1% and 35.4%,respectively,in leaves and 27.0% and 16.1%,respectively,in stems.Although stem P levels under SD increased significantly by 15.6% compared to the controls,leaf phosphorous did not change (P>0.05).Under LD,P increased in the coarse roots by 51.0% compared to CK (P<0.05),while it was insignificant for the fine roots.Under MD,P of both the coarse and fine roots did not change compared to the controls (P>0.05),while under SD,both increased (P<0.05),with the coarse root P increasing to more than twice that of the controls and fine root P increasing by 66.6%. Fig.2 Changes in C,N,and P and their stoichiometric ratios in Pinus yunnanensis seedlings under different drought treatments;capital letters indicate the same degree of drought with significant differences among various organs (P <0.05);different lowercase letters indicate the same organ with significant differences in degrees of drought (P <0.05) The pattern of variation in the C:N ratio of stems and coarse roots was consistent,dropping with increasing drought,with significant decreases of 74.3% and 55.6% under SD,respectively.Compared with the controls,leaf and fine root C:N ratios were not different under LD but decreased significantly under MD and SD.Leaf C:N decreased by 31.1% and 23.5%,and the fine root C:N ratio decreased by 33.1% and 47.0%,respectively.Leaf,stem,coarse root,and fine root C:P ratios increased and then decreased with an increase in drought.Compared with CK,under LD,the leaf and fine root C:P ratios increased significantly by 55.7% and 71.8%,respectively,and the coarse root C:P ratio increased almost 2.5-fold compared to CK.As the degree of drought intensified,the N:P ratio of leaves,coarse roots,and fine roots first rose and then fell,and all rose significantly under MD,with the leaf and coarse root N:P ratio rising to greater than 2 times and greater than 5 times that of controls,respectively.The fine root N:P ratio increased by 81.4% compared with the controls.The stem N:P ratio gradually increased with greater drought,increasing to 2.8 times that of the controls under SD.Under different drought treatments,there was no significant difference in carbon levels of leaves,stems,and coarse roots,while nitrogen concentration of leaves and fine roots was significantly higher than that of stems and coarse roots.Under the controls,P in coarse roots was the lowest,while under LD and SD treatments,it was higher than in other organs.The C:N ratio of leaves was smaller than that of other organs under different treatments.The C:P ratio of fine roots and the N:P ratio of leaves were higher than those of other organs under CK,LD,and MD treatments,while they were highest under SD treatment. Analysis of the correlation between NSCs composition and C,N,and P stoichiometric ratios among organs revealed that stem N was significantly positively correlated with soluble sugar and NSCs concentration (P<0.05) and significantly negatively correlated with ratio of soluble sugar to starch (P<0.01) (Fig.3).C:N ratios were negatively correlated with soluble sugar and non-structural carbohydrates levels (P<0.05),while N:P correlated positively with soluble sugar and non-structural carbohydrates concentrations.N levels of coarse roots were positively correlated with starch concentration (P<0.05),and C:N ratios were negatively correlated with starch and non-structural carbohydrates concentrations (P<0.05).Non-structural carbohydrates of fine roots were negatively correlated with C and P levels (P<0.05). Fig.3 Correlation between NSC composition,C,N,and P concentrations,and their stoichiometric ratios of Pinus yunnanensis seedlings under drought;A,B,C,and D represent leaves,stems,coarse roots,and fine roots,respectively Fig.4 Phenotypic plasticity index of non-structural carbohydrates,C,N,and P concentrations,and their stoichiometric characteristics in Pinus yunnanensis seedlings Fig.5 Variation coefficients of non-structural carbohydrates,C,N,and P concentrations,and their stoichiometric characteristics in Pinus yunnanensis seedlings As shown in Figs.4 and 5,the top five items with greater plasticity indices of NSC characteristics under drought were leaf ratio of soluble sugar to starch (0.49),stem starch (0.40),leaf starch (0.39),stem ratio (0.38),and fine root ratio (0.37).The top five items with large coeffi-cients of variation were fine root ratio (38.6%),stem starch(31.9%),coarse root ratio (30.0%),coarse root starch(28.4%),and leaf ratio (27.4%).The plasticity index of leaf ratio of soluble sugar to starch was the largest,while its NSC plasticity index (0.13) and coefficient of variation(10.0%) were the smallest.The plasticity index of C,N,and P concentrations and their stoichiometric characteristics were more than 0.7 in three components: from large to small,coarse root N:P (0.80),stem C:N (0.74),and coarse root C:P (0.72).The first three with large coefficients of variation were coarse root N:P (62.9%),stem N:P (62.5%),and stem C:N (61.1%).The plasticity index of C,N,and P concentrations and their stoichiometric characteristics were larger than that of NSCs,and those that were more sensitive to plasticity changes had greater variability.The correlation between the degree of physiological response ofP.yunnanensisseedlings to drought and their variability (Fig.6) was in accordance with a linear equation fit:y=74.118x+2.0803,R2=0.8416,P<0.0001. Fig.6 Correlation between the coefficient of variation of non-structural carbohydrates,C,N,and P concentrations,their stoichiometric characteristics,and the plasticity index of Pinus yunnanensis seedlings;solid line represents correlations;dotted lines represent 95% confidence intervals As a product of photosynthesis,NSCs are a major storage material in the synthesis and utilization of energy in trees and serve as a carbon source buffer material when drought occurs,temporarily supplying plants with the energy required for growth and metabolism and playing an important role in maintaining the plant’s functional stability (Deng et al.2019).In this study,the intensity of drought had a significant effect (P<0.05) on NSCs levels(Table 1).Non-structural carbohydrates concentration of fine roots decreased with increasing drought (Fig.1),possibly because NSCs transport followed the top–down proximity principle (Dietze et al.2014).The increase in drought accelerated seedling respiration,which led to Non-structural carbohydrates depletion and inhibited its synthesis,making the already limited amount unavailable to fine roots.It is also possible that transport channels were blocked,and non-structural carbon from other organs could not be transported to the fine roots;thus,the fine roots had to rely on their own stores to cope with drought (Pinheiro and Chaves 2011;Hartmann et al.2013).Non-structural carbohydrates concentrations increased in all other organs (Fig.1),and the increase was mainly caused by the increase in soluble sugar in each organ.This may be because plants that reduce their rate of growth have less demand for carbon under drought.When diminishing C assimilation is slower than the reduction in growth,photosynthetic products are stored in the form of NSCs (Li et al.2020).Non-structural carbohydrates concentrations of the coarse roots increased(Fig.1),which may be due to the continuous transport of NSCs in the fine roots to the coarse roots to promote the growth and water uptake of the coarse roots.This is also to maintain the osmotic potential of the coarse roots,resulting in an increase in NSCs concentration of coarse roots but a decrease in NSCs concentration of fine roots.In addition,drought restricted seedling growth and impeded respiration,and NSCs were reduced in all organs,resulting in its accumulation in each organ as a response to withstand stress. The changes in starch in various seedling components under drought differed from changes in soluble sugar.In this study,the intensity of drought had a significant effect on levels of soluble sugar,while the effect on starch was insignificant (P>0.05) (Table 1).Starch is the main energy storage substance in plants.In the face of various stresses,starch and soluble sugar can alternate each other and be transferred to various organs.When soluble sugar concentrations are high,the remaining part can be converted into starch for storage in addition to its consumption for growth.In contrast,when soluble sugar levels are low,starch can be broken down into soluble sugar to maintain regular plant physiological activities.In studies onPinus ponderosaDouglas ex C.Lawson (Sala and Hoch 2009),Robinia pseudoacaciaL.(Maguire and Kobe 2015),andPopulusspp.L.(Wang et al.2015),when water deficits reached a specific level,starch was converted to soluble sugar which increased,and cellular osmoregulation was enhanced to maintain metabolism and promote water uptake,thus resisting the gradually increasing stress by drought.The soluble sugar:starch ratio is an index of the transformation efficiency of soluble sugar and starch;the higher the value,the greater the plant demand for soluble sugar.This study found that the range of soluble sugar:starch in each organ ofP.yunnanensisseedlings under drought was 1.7–3.9,obviously greater than 1,indicating that the proportion of soluble sugar in each plant component was significantly higher than starch,suggesting that soluble sugar were the main energy supply for the growth ofP.yunnanensisseedlings and played an important role in resisting drought.In this study,as the intensity of drought increased,soluble sugar increased in all organs except in the fine roots(Fig.1),thereby increasing the osmoregulatory capacity of cells and improving the drought tolerance of seedlings.Starch increased in stems and fine roots,increased in leaves under light drought and moderate drought stress levels and decreased under severe drought.Starch levels in coarse roots increased under severe drought treatment (Fig.1).This may be because,with the intensification of drought,photosynthesis of seedlings is blocked and photosynthetic products are reduced.To maintain physiological metabolic activity,needles converted stored starch to soluble sugar.At the same time,under drought,the utilization efficiency of soluble sugar was reduced,resulting in the accumulation of starch in stems,and in coarse and fine roots.In addition,to resist long-term drought,starch stores,mainly stored in stems and roots,and helps promote root growth to maintain the osmotic potential to absorb more water.If the future environments are suitable,seedlings may rapidly recover the morphological reconstruction of tissues and organs,similar to the results onCaryopteris mongolicaBunge seedlings by Shen et al.(2019). The distribution of NSCs in different plant organs reflects the response patterns and adaptation strategies to drought.In this study,stem soluble sugar concentrations were higher than that of the leaves and roots under all stress treatments.Starch and NSC concentrations increased with greater drought and was higher than that of other organs under moderate drought and severe drought,a process in which the increase in stem soluble sugar may be derived from photosynthetic products rather than from starch conversion.Starch levels of coarse roots were higher than in other organs under the controls and then decreased.With the onset of drought,starch levels of stems and coarse roots was higher than in other organs under severe drought,indicating that stems and coarse roots were storage organs in drought environments. Water deficits affect physiological and biochemical processes such as C,N,and P uptake,transport,and accumulation,which leads to changes in nutrient cycling and distribution among organs (Ventura et al.2008).The results of this study show that drought had no significant effect on stem or coarse root C concentrations,and that leaf C decreased under light drought and fine root C concentration increased(Fig.2).This may be because leaves under light drought transfer fixed C of photosynthesis to the fine roots to increase the vitality of the seedlings and promote water absorption.Carbon levels of fine roots were higher than that of other organs under all stress treatments (Fig.2),possibly because seedlings under drought need to absorb more water through their root systems.Fine roots are the main water-absorbing organ,leading to an increase in carbon levels of fine roots.Carbon is the basic element that constitutes the skeleton of organic matter,and stems and coarse roots play a supporting role of seedlings,which have a more stable C concentration.Carbon levels of needles under different drought treatments was not significantly different from that of the controls (Fig.2),which may be because seedlings under drought close stomata,reduce transpiration,and limit C fixation.Drought also reduces C consumption due to growth limitations,and carbon acquisition and consumption are in equilibrium,so there is no significant change in C in needles,which is similar to the results of Wang et al.(2019) forCaragana microphylla. Nitrogen levels in needles,coarse and fine roots increased under drought,except for the highest N concentration in fine roots under severe drought and the highest N concentration in leaves under all other treatments (Fig.2).Under drought,plants allocate more N to leaves to improve water utilization and obtain an adequate photosynthetic volume at a smaller stomatal conductance to maintain growth and C assimilation(Wright et al.2001).However,plant growth has a dilutive effect on nitrogen levels (Wang et al.2018),which is lower than the drought treatment due to the inhibition of growth and normal growth under suitable moisture.In addition,as drought intensified,plant needles were damaged and photosynthesis decreased,while seedlings under severe drought grew rapidly by allocating more nitrogen to the fine roots to obtain more water and nutrients. Phosphorous levels of all seedling components under the severe drought treatment were higher than other drought treatments (Fig.2),possibly because the seedlings reduced growth to store more P to ensure survival and to prepare for more stress.In addition,protein synthesis during growth requires large amounts of P (Elser et al.2010),and severe drought limits growth,consuming less P.Embolisms form in the xylem due to drought (Li et al.2016),which ultimately affected the P transportation and accumulation in various organs. N and P stoichiometric characteristics are important indicators of plant adaptation strategies to environmental changes,reflecting C assimilation capacity and N and P nutrient limitations.C:N and C:P ratios are key indicators for determining nutrient use efficiency,which ultimately reflect the ability of plants to absorb nutrients for carbon assimilation (Zhang et al.2016).In this study,C:N ratios in all organs were reduced (Fig.2),indicating that drought affected N use efficiency which in turn affected C assimilation capacity and nutrient use efficiencies (Fig.2).Under light drought and moderate drought,C:P ratios in each organ was higher than in the controls,while C:P under severe drought was lower (Fig.2).This indicated thatP.yunnanensisseedlings maintained normal water and nutrient transport to adapt to drought by improving the P utilization rate under light drought and moderate drought.While seedling growth under severe drought was restricted,P consumption was reduced,transport was blocked,and P accumulated in the organ,leading to a decrease in C:P ratios.Leaf N:P ratios can reflect the status of plant productivity as limited by nutrients (Yuan et al.2011),and usually it is considered that growth is mainly limited when N:P <14.When N:P >16,growth is mainly limited by P;when N:P is between 14 and 16,growth is limited by both nutrients (Han et al.2005).In this study,needle N:P ratios were <14 under different degrees of drought,indicating that seedling growth was generally limited by N. Tree species rely on phenotypic plasticity or the ability to adjust their acquisition,utilization,and consumption of environmental resources under stress.The magnitude of their capacity to adapt to stress is reflected by the phenotypic plasticity index (Savva et al.2007).The phenotypic plasticity indices ofP.yunnanensisseedlings in this study were high for leaf soluble sugar:starch (0.49),stem starch (0.40),leaf starch (0.39),stem soluble sugar:starch (0.38),coarse root N:P (0.80),stem C:N (0.74),and coarse root C:P (0.72).A comparison of various indicators of plasticity in different organs showed thatP.yunnanensisseedlings responded to drought mainly by altering starch and soluble sugar:starch ratios in leaves and stems and in nutrient regulation by altering N and P utilization efficiency.In this study,the ranking of phenotypic plasticity indices was not consistent with the ranking of coefficients of variation.The degree of phenotypic plasticity change in each organ varied considerably,while they varied synergistically.This asymmetry is prevalent in ecosystems (Peng et al.2013),suggesting an adaptive adjustment in trade-offs between leaf,stem,and coarse and fine root indices ofP.yunnanensisseedlings under drought. There were correlations between NSC levels and the composition of leaves,stems,and coarse and fine roots and their C,N,and P concentrations and stoichiometric characteristics.There was a positive correlation between N in stems and coarse roots and NSCs and its component concentrations(P<0.05) (Fig.3).Leaf N:P ratios can be used to determine whether plant growth is limited by N and/or P.In this study,leaf N:P was <14 under different drought levels,and therefore growth ofP.yunnanensisseedlings was limited by N.Nitrogen is the material basis of life,and it is also the main component of chlorophyll,proteins,nucleic acids,and phospholipids (Dawson and Curran 1998;Li et al.2012).It is crucial for the influence of N level variations among the organs ofP.yunnanensisseedlings on the concentration of NSCs and their components and to govern the interconversion of soluble sugar and starch.Changes in N concentrations during drought can be a strong indicator of the carbon metabolism and growth status of seedlings. The intensity of drought had a significant effect on both soluble sugar and NSC levels,but not on the starch concentrations and soluble sugar: starch ratios.With drought intensification,soluble sugar and NSC concentrations generally increased in leaves,stems,and coarse roots,while decreasing in fine roots.Under severe drought,except for the NSC concentration of leaves,the NSC of stems and coarse roots increased by 40.0% and 16.5%,respectively,while it decreased by 17.9% in fine roots.The effect of drought on C levels was insignificant;there was a significant effect on N and P and on C:N,C:P,and N:P ratios.With drought intensification,N levels increased in all organs but the utilization efficiency decreased.Phosphorous utilization efficiency in all organs increased under light drought and moderate drought,while the efficiency of N and P utilization decreased under severe drought.Growth under drought was mainly limited by nitrogen.Nitrogen limitation first weakened and then increased with an increase in drought intensity.There was a correlation between NSC,C,N,and P concentrations and their stoichiometric characteristics in each seedling organ.Changes in N levels play a crucial role in changes in the concentration of NSCs and their components and affect the interconversion between soluble sugar and starch.A comprehensive comparison of various indicators of plasticity in different organs showed that seedlings responded to drought mainly by altering starch and ratio of soluble sugar to starch in leaves and stems,and in nutrient regulation by altering efficiency of nitrogen and phosphorous utilization. AcknowledgementsWe thank LetPub (www.letpub.com) for its linguistic assistance during the preparation of this manuscript. Author contributionZZ wrote the manuscript,JW designed the experiments,provided critical revisions and final approval of the article.LW,YL,JS and JX carried out the experiments and ran the data.All authors helped to write and approve the final manuscript. Data availabilityThe datasets are available from the corresponding author on reasonable request. Declarations Conflict of interestThere are no conflicts of interest to declare.Data analysis
Results
Effects of drought on non-structural carbohydrate concentrations of P. yunnanensis


Effects of drought on C,N,P concentrations and their stoichiometric characteristics in seedling organs


Relationship between non-structural carbohydrates and C,N,and P stoichiometric ratios of P. yunnanensis seedlings
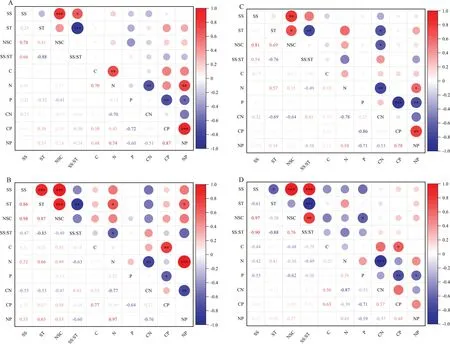


Plasticity of each index in different organs of P.yunnanensis seedlings under different drought levels

Discussion
Effect of drought on non-structural carbohydrates in Pinus yunnanensis seedlings
Effects of drought on C,N,and P stoichiometric characteristics
Phenotypic plasticity of non-structural carbohydrate,C,N,and P concentrations,and their stoichiometric characteristics
Relationship between non-structural carbohydrate and C,N,and P stoichiometry characteristics among organs
Conclusion
杂志排行

Journal of Forestry Research的其它文章
- New indices to balance α-diversity against tree size inequality
- Climate-growth relationships of Pinus tabuliformis along an altitudinal gradient on Baiyunshan Mountain,Central China
- Nitrogen mineralization in the oldest climax communities in the eastern Mediterranean region
- Physiological and psychological responses to tended plant communities with varying color characteristics
- Impact of cattle density on the structure and natural regeneration of a turkey oak stand on an agrosilvopastoral farm in central Italy
- Peat properties of a tropical forest reserve adjacent to a fire-break canal