柳江不同河网位置大型底栖动物群落特征及其影响因子差异比较研究
2024-01-22周佳诚宋志斌苗芃谭路唐涛
周佳诚,宋志斌,苗芃,谭路,唐涛*
1. 中国科学院水生生物研究所,湖北 武汉 430072;2. 中国科学院大学,北京 100049
环境过滤和空间扩散过程如何影响生物群落的空间格局是淡水生态学近年来关注较多的问题之一(Gothe et al.,2013;Sarremejane et al.,2017;Mruzek et al.,2022)。大多数研究发现,虽然河流生物多样性格局主要受环境过滤作用驱动(Astorga et al.,2012;Grönroos et al.,2013;He et al.,2023),但是空间过程因为限制生物扩散的距离和方向,所以对河流生物多样性也有重要影响(Cottenie,2005,Karna et al.,2015,Lansac-Toha et al.,2021)。不过现有研究多将同一区域的所有调查样点视为一个整体,探究环境过滤和空间扩散过程对生物多样性的影响差异。
然而,河流独特的树枝状网络结构制约着水生生物的空间迁移和扩散(Chaput-Bardy et al.,2017),对区域水生生物多样性格局具有重要影响。通常来说,河网边缘河段之间相对孤立,生物在这些位置间的扩散往往较困难(Brown et al.,2010)。而河网中心位置对上游河流生物具有汇聚作用,使得物种扩散变得相对容易(Altermatt et al.,2013;Trigal et al.,2015)。因此,空间扩散过程对河网边缘和中心位置的水生生物多样性具有不同的贡献(Brown et al.,2011;Heino et al.,2015)。此外,不同河网位置的栖息地环境也有较大差异。河网边缘由于连通性低,不同河段间的水体理化环境异质性较高(Meyer et al.,2007;Clarke et al.,2008)。河网中心由于连通性高,水体中的物质随水流在此汇集、交换,该区域的水体理化状况差异变小。因此,环境因子对河网边缘和中心位置的生物多样性的影响也具有明显差异(Lansac-Toha et al.,2021)。虽有少数研究比较了不同河网位置生物多样性的差异,但多集中于比较物种数量的变化(Hitt et al.,2011;Mazor et al.,2016),而对群落组成的影响差异研究较少。
大型底栖动物(Benthic Macroinvertebrate)是指生命周期的全部或者至少一段时间生活在水体底部的大于500 μm 的水生无脊椎动物群(刘建康,1999)。河流大型底栖动物多样性高,时空异质性分布特征明显,且对环境变化响应敏感(Williams-Subiza et al.,2020;Masese et al.,2021;Quanz et al.,2021),作为水生生物的重要组成部分,一直是河流生态学关注的重要类群之一。本研究基于柳江水系大型底栖动物的调查数据,探究水系内的河网边缘和河网中心位置之间的大型底栖动物群落格局差异,并比较不同河网位置环境因子和空间因子对大型底栖动物群落组成的影响差异。拟验证以下科学问题:大型底栖动物群落组成差异在河网边缘受环境因子和空间因子的共同影响,在河网中心主要受环境因子的影响。
1 材料与方法
1.1 研究区域与样点设置
柳江位于107°27′—110°34′E,23°41′—26°30′N间,发源于贵州省独山县,在广西壮族自治区象州县汇入西江,是西江水系第二大支流(卢玉典等,2023)。柳江干流全长751 km,流域面积为58 520 km2,流域内水系发达,地势西北高东南低,大部分属喀斯特地貌(杨昆等,2016)。该区域属亚热带季风气候区,四季较分明,年均温18—20 ℃,年降水量1 400—1 800 mm,4—8 月为雨季,雨季降水量占全年72%(李佳静等,2020)。流域内森林植被丰富,生境复杂多样,有较高的生物多样性。
于2022 年3—4 月低水位期对柳江水系81 个样点开展调查。本研究根据Strahler 法(Strahler,1957),以柳江1∶1 250 000 水系图为依据,确定各调查样点的河流级别。没有任何支流汇入的源头河流为1 级河流,两个同级别的河流汇流后河流级别便升高1 级,两个不同级别的支流汇流后河流级别仍然为汇流前级别较高的那一级,依次类推,直到最下游样点。研究中将1、2 级河流样点视为河网边缘样点,大于2 级的河流样点作为河网中心样点(Henriques-Silva et al.,2019)。按此分类,共有39个样点位于河网边缘位置,42 个样点位于河网中心位置(图1)。

图1 柳江流域样点分布图Figure 1 Locations of the sampling sites in the Liujiang River network
1.2 大型底栖动物数据
使用网筛孔径为0.42 mm 采样面积为0.09 m2的索伯网(Surber net)采集大型底栖动物。可涉水样点在整个河段选择采样区域,不可涉水样点则在水深小于0.5 m 的近岸河段采样。每个样点在纵向100 m 范围内根据不同底质类型所占比例采集5 个重复。采样时将索伯网放置于河床上,先随水流仔细清洗网内的大型石块,使石块上的大型底栖动物流进索伯网内,然后搅动河床下的底质,搅动的深度大于10 cm,使底质中的大型底栖动物随水流进入索伯网内。采样结束后将采集到的大型底栖动物拣出,将每个样点的5 个重复样本装入同一个标本瓶,并用95%的乙醇溶液现场固定。标本带回实验室后,参照国内外相关分类文献(Morse et al.,1994;Merritt et al.,2019;周长发等,2003),在解剖镜下鉴定并计数。大部分动物鉴定到属级别,摇蚊等少数类群鉴定到亚科级别。根据鉴定数据计算大型底栖动物的分类单元数、密度及平均相对丰度,并将平均相对丰度大于1%的分类单元定义为优势分类单元。
1.3 环境因子
大型底栖动物采样前,用水质分析仪(YSI ProPlus)测定水体溶解氧(Dissolved oxygen,DO)、电导率(Conductivity,Cond)和浊度(Turbidity,Turb)。此外,用预先清洁的聚乙烯瓶采集约100 mL水样,并用浓硫酸酸化至pH<2,低温保存运回实验室。然后,根据《水和废水监测分析方法》利用连续流动水质分析仪(SAN++,Skalar)测定总氮(Total nitrogen,TN)、总磷(Total phosphorus,TP)和化学需氧量(Chemical oxygen demand,COD)等水质指标(国家环境保护总局,2002)。采集底栖动物的同时,测量每个采集处的水深,将5 个重复采样处的平均水深作为样点的水深。用直读式流速仪(Global Water FP101)测定每个大型底栖动物采集处的流速,将5 个重复采样处的平均流速作为样点的流速。
1.4 空间因子
大部分河流大型底栖动物属水生昆虫,它们在幼虫时期主要随水流沿河道扩散,成虫阶段有翼,可以在陆上进行飞行扩散(Canedo-Argueelles et al.,2015;Arce et al.,2023)。为量化大型底栖动物的空间扩散过程,根据采样点的经纬度坐标计算2 个距离指标:陆上距离(Overland distances)和河道距离(Watercourse distances)。陆上距离是指样点间的直线(欧几里得)距离,河道距离表示沿河流网络样点间的成对最小路径距离。这两种距离都使用ArcGIS 10.5(ESRI,2017)软件计算,其中陆上距离用点距离分析工具计算,河道距离则用Network Analyst 工具计算。
1.5 数据分析
分析前对大型底栖动物丰度数据和除pH 外的环境因子数据进行log(x+1)转换,以降低极端值的影响,并根据Pearson 相关系数剔除相关性较高的环境因子(r>0.8)。
首先,用Mann-Whitney U 非参数检验分析河网边缘和河网中心的环境因子、大型底栖动物分类单元数和密度的差异。其次,基于每个样点每种大型底栖动物的密度数据计算样点间的 Bray-Curtis 相异度作为β 多样性指数,Bray-Curtis 相异度的取值范围由0(两个群落组成完全一致)到1(两个群落完全不同)。用基于Bray-Curtis 相异度的非度量多维尺度分析(Non-metric Multidimensional Scaling,NMDS)展示河网边缘和河网中心的大型底栖动物群落组成差异。该分析设定,当Stress 值小于0.2 时,表示NMDS 结果合理(Khalaf et al.,2002)。使用置换多元方差分析(Permutational multivariate analysis of variance,PERMANOVA)进一步检验河网边缘和河网中心大型底栖动物群落的差异显著程度。该方法是基于F 统计的方差分析,依据距离矩阵(本研究为Bray-Curtis 相异度)对总方差进行分解的非参数多元方差分析方法(Anderson,2001)。接着,用“bio-env”函数筛选出与大型底栖动物群落相关性最高的环境因子组合。“bio-env”函数能够对所有可能的环境因子组合进行检验,并计算出群落相异性矩阵和不同环境因子组合之间的相关性,从而得出与群落相关性最高的环境因子组合(Clarke et al.,1993)。最后,使用Mantel test 分析检验大型底栖动物Bray-Curtis 相异度(β 多样性)与环境距离矩阵(使用 “bio-env” 筛选出的环境因子计算欧氏距离)、空间距离矩阵(陆上距离矩阵、河道距离矩阵)之间的相关性(Astorga et al.,2012)。当Mantel 相关性指数r>0,表示大型底栖动物β 多样性随着环境距离、空间距离的增加而增加。此外,应用Partial Mantel test 分析来检验环境距离矩阵和空间距离矩阵对大型底栖动物β 多样性的单独影响。
以上分析均在R(Version4.1.3)中进行。Mann-Whitney U 检验使用“stats”包,置换多元方分析(PERMANOVA)、非度量多维尺度分析(NMDS)、bio-env 和Mantel 检验均使用“vegan”包。
2 结果
2.1 理化环境状况
根据国家《地表水环境质量标准》对柳江水质状况进行分析(国家环境保护总局等,2002)发现:总氮、总磷浓度在河网边缘多数样点达到Ⅲ类水质标准,在河网中心多数样点达到Ⅳ类标准;溶解氧和化学需氧量含量在河网边缘和河网中心大都达到I类水质标准。总体而言,河网边缘样点的水质优于河网中心样点。Mann-Whitney U 检验发现,大部分理化环境指标在河网边缘和中心位置间存在显著差异。电导率、总氮、化学需氧量和水深在河网边缘较低,而溶解氧和流速则在河网边缘较高(表1)。

表1 河网边缘和河网中心环境因子差异Table 1 Statistics and differences in environmental factors between the edge and center positions of the Liujiang River network
2.2 不同河网位置大型底栖动物群落格局差异分析
2.2.1 物种组成及优势种差异
本研究共采集到大型底栖动物5 门8 纲21 目75 科132 个分类单元。分别隶属于节肢动物门的昆虫纲和甲壳纲、环节动物门的蛭纲和寡毛纲、软体动物门的腹足纲、双壳纲、线形动物门的铁线虫纲,扁形动物门的涡虫纲。其中,昆虫纲共109 个分类单元,占总分类单元数的82.6%。
河网边缘、中心位置样点采集到的大型底栖动物分类单元数分别为129 和114 个,河网边缘、中心位置的平均分类单元数分别为32 和22,平均密度分别为2.29×103ind·m-2和1.04×103ind·m-2。差异性检验显示河网边缘样点的大型底栖动物丰富度和密度均较河网中心位置高(图2)。

图2 不同河网位置大型底栖动物分类单元数和密度箱线图Figure 2 Boxplots displaying differences in taxonomic units and density of benthic macroinvertebrates between the edge and center positions of the Liujiang River network
平均相对丰度大于1%的大型底栖动物分类单元在河网边缘有12 个,在河网中心有10 个(表2)。花翅蜉属(Baetiellasp.)和高翔蜉属(Epeorussp.)为河网边缘独有的优势分类单元。在河网边缘和中心位置样点共有的10 个优势分类单元中,四节蜉属(Baetissp.)、宽基蜉属(Choroterpessp.)、扁蜉属(Heptageniasp.)、细蜉属(Caenissp.)、锯形蜉属(Serrattellasp.)等蜉蝣目类群和蚋(Simuliumsp.)在河网边缘的平均相对丰度较高,而直突摇蚊亚科(Orthocladiinae)、长足摇蚊亚科(Tanypodinae)、摇蚊亚科(Chironominae)、朝大蚊属(Antochasp.)等双翅目类群则在河网中心的平均相对丰度较高(表2)。
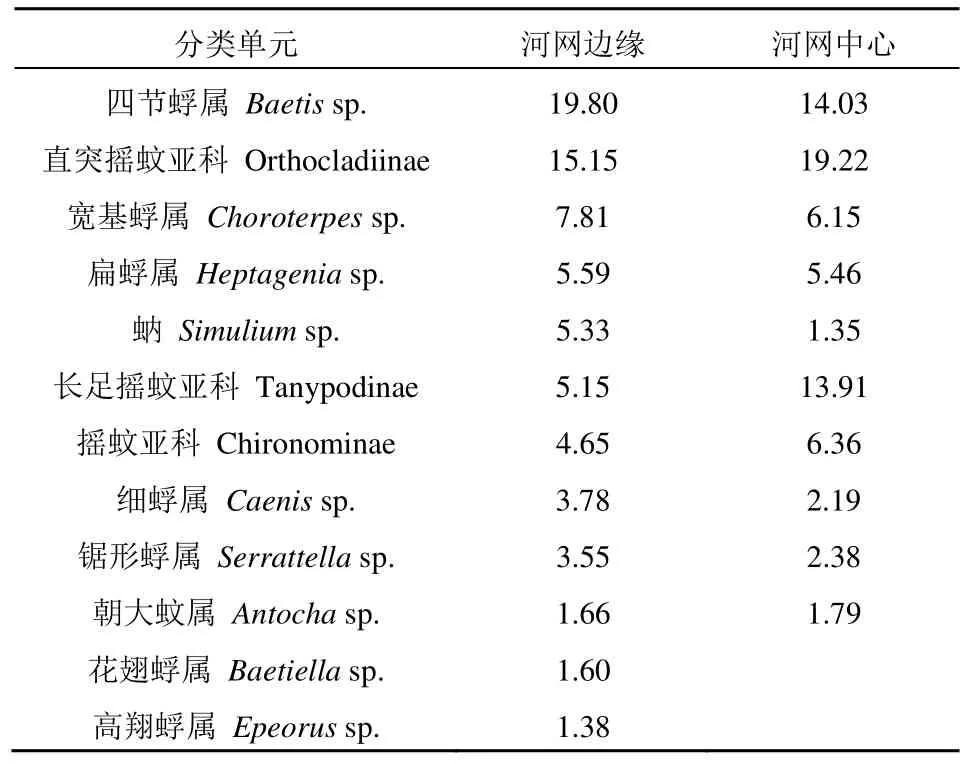
表2 不同河网位置优势分类单元相对丰度Table 2 Relative abundance of dominant taxonomic units of benthic macroinvertebrates between the edge and center positions of the Liujiang River network %
2.2.2 群落相似度差异
非度量多维尺度分析(NMDS)排序图显示,柳江水系河网边缘位置与和中心位置的样点在NMDS 轴1 上有明显分离(图3)。置换多元方差分析(PERMANOVA)表明,河网边缘和中心位置样点的大型底栖动物群落组成存在显著差异(F=4.96,P=0.001)。
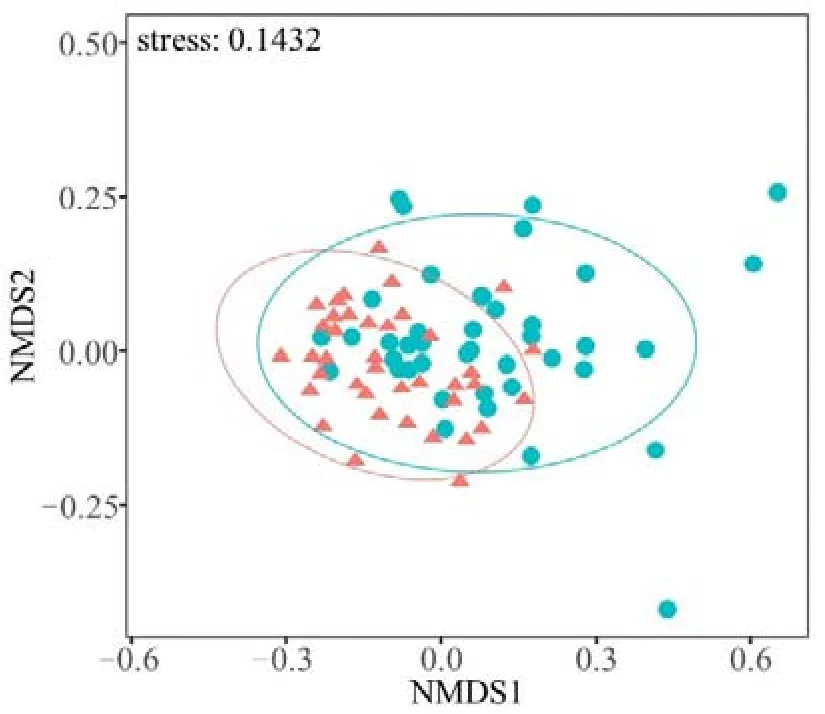
图3 基于Bray-Curtis 相异度的柳江大型底栖动物群落组成非度量多维尺度分析(NMDS)排序图Figure 3 Non-metric Multidimensional Scaling(NMDS) plot based on inter-site Bray-Curtis dissimilarity of benthic macroinvertebrate communities in the Liujiang River network
2.3 不同河网位置大型底栖动物β 多样性与环境距离、空间距离的关系差异
用“bio-env”筛选出与河网边缘大型底栖动物群落最相关的环境因子。其中,溶解氧和水深与河网边缘和中心位置的大型底栖动物组成都显著相关;此外,河网边缘样点的大型底栖动物组成还与化学需氧量有关,而河网中心位置样点则还受流速、总氮的影响(表3)。

表3 大型底栖动物β 多样性与空间距离(陆上、河道)和环境距离的相关性Mantel 检验Table 3 The correlations between benthic macroinvertebrate beta diversity and spatial distances (Overland, Watercourse)and environmental distances with Mantel test and partial Mantel test
Mantel test 分析显示(表3),河网边缘大型底栖动物β 多样性与环境距离、陆上距离、河道距离皆显著相关,两个空间距离对大型底栖动物β 多样性的单独影响相近,而环境距离的单独影响约为空间距离的2 倍。河网中心大型底栖动物β 多样性与环境距离、陆上距离显著相关,但陆上距离主要通过与环境距离的协同作用影响β 多样性。
3 讨论
通过分析柳江水系的理化环境指标发现,河网边缘位置电导率、总氮、化学需氧量和水深比河网中心低,而溶解氧和流速比河网中心高。河网边缘位置因为水浅、光照充足,水生植物光合作用强,溶解氧含量高。而中心位置水深增加,水生植物光合作用减弱,溶解氧减少。现场调查时发现,中心位置样点附近多有城镇,并且存在工业污染。显然,过多的人类活动可能是导致水体中总氮、化学需氧量和电导率升高的主要原因,以往关于柳江的研究也证实了这点(李鑫等,2021;张婉军等,2021)。此外,河网中心位置由于水域开阔,仅在浅水缓流区采样,所以采样时测定的流速较低。
本研究发现,柳江水系不同河网位置的环境特征和大型底栖动物群落格局存在明显差异。河网边缘大型底栖动物物种丰富度和密度都显著高于河网中心,这与河网边缘位置水浅、溶解氧含量较高等环境特点有关(Richardson,2019)。当然,因河网边缘位置大部分样点为可涉水河段,为全河段采样,而河网中心位置由于水较深,大部分样点仅在浅水区采样,采样不充分等因素也可能影响了河网中心的大型底栖动物多样性。与柳江河网位置的环境特点相对应,调查发现虽然河网边缘与河网中心位置的优势分类单元大都为蜉蝣目和双翅目,但耐水流冲刷的能力强,更适应高氧环境(Poff et al.,2006;Belmar et al.,2013)的蜉蝣目类群平均相对丰度在河网边缘位置更高。而更适应相对低氧环境的双翅目类群平均相对丰度则在河网中心位置较高(Calapez et al.,2021)。此外,边缘位置的大型底栖动物还受化学需氧量影响。因为这些位置的河流靠近河岸带(Turunen et al.,2017),有机物(如枯枝落叶)较多,影响水体中化学需氧量的浓度(杨钙仁等,2012),但这些有机物也可作为大型底栖动物的食物来源(Oester et al.,2023),从而影响其生长。中心位置的大型底栖动物还受流速和总氮影响。较慢的流速可能造成溶解氧供应不足,影响需氧量高的大型底栖动物生存。总氮的增加可能引发藻类的大量生长(Dodds,2006),进而影响大型底栖动物的分布。本研究表明无论是在河网边缘还是河网中心,环境距离与大型底栖动物β 多样性均显著相关。并且环境距离与大型底栖动物β 多样性的相关性比空间距离更高,说明环境因子对大型底栖动物群落组成的影响比空间因子更大(Gothe et al.,2013;He et al.,2020;Zheng et al.,2023)。
通过分析不同河网位置空间距离与大型底栖动物β 多样性的关系,发现空间距离(陆上距离、河道距离)与河网边缘的大型底栖动物β 多样性显著相关,但在河网中心则关联性不显著。因为河网边缘的群落之间相对孤立,大型底栖动物在这些位置之间扩散更加困难而导致群落相异度增加(Wang et al.,2018;Gauthier et al.,2020;Li et al.,2021)。而在河网中心受水流单向流动和交汇作用等的影响,不同样点间的大型底栖动物物种交换、补充变得相对容易。所以,河网中心的大型底栖动物群落格局主要受环境因子的影响。
4 结论
通过比较柳江水系不同河网位置的大型底栖动物群落格局发现,河网边缘大型底栖动物物种丰富度和密度都显著高于河网中心位置。溶解氧、水深、化学需氧量、总氮和流速是影响大型底栖动物群落空间分布的主要环境因子。大型底栖动物群落格局在河网边缘受环境因子和空间因子的共同影响,在河网中心则主要受环境因子影响。
