Eicosapentaenoic acid代谢产物促进胰岛α细胞转分化为胰岛β细胞的作用研究
2024-01-22邢朝凤唐敏怡徐绮华张宗猛赵子建穆云萍李芳红
邢朝凤,唐敏怡,徐绮华,王 帅,张宗猛,赵子建,穆云萍,李芳红
(广东工业大学生物医药学院,广东 广州 510006)
1型糖尿病(type 1 diabetes mellitus,T1DM)主要是由自身免疫T淋巴细胞介导胰岛β细胞破坏,导致胰岛素分泌严重不足引起[1]。目前T1DM治疗以终身每日注射重组胰岛素为主,然而,注射外源性胰岛素时常发生严重的低血糖等不良反应。大量研究尝试利用特异的单克隆抗体作为自身免疫的拮抗药物[2],以干预自身免疫系统攻击胰岛的进程。但是这类药物非常昂贵,有各种副作用,不适合长期应用于T1DM治疗。
自Edmonton Protocol 10公布以来,异体胰岛移植曾经为T1DM患者终身摆脱胰岛素治疗,根治糖尿病带来很大的希望。2000年的研究揭示了使用胰岛移植替代胰岛素疗法治疗T1DM的潜力,7名患者持续1年没有补充外源性胰岛素[3]。然而,由于胰腺供体的匮乏,并且移植的胰岛在一年左右开始纤维化[4],长期存活效果欠佳,由于患者需要长期使用免疫抑制剂和抗排异反应的药物等一系列原因,近期实施临床胰岛移植的患者数量大幅减少,已经无法成为摆脱注射胰岛素治疗T1DM的新型疗法。因此,探索低免疫,增加胰岛素分泌的疗法,成为糖尿病治疗倍受期待的选择。
最近的一系列遗传学和临床研究揭示了ω-3多不饱和脂肪酸(ω-3 PUFAs)在抑制自身免疫以及保护和增强胰岛功能方面的机制和治疗潜力[5]。在动物研究中,有研究表明,妊娠期和哺乳期母体饮食中ω-6/ω-3 PUFAs的比率较低(3.0),推迟了非肥胖型糖尿病(non-obese diabetes,NOD)小鼠后代糖尿病的发病时间[6]。两项大规模临床研究显示,通过对婴儿和儿童食用富含EPA和二十二碳六烯酸(docosahexaenoic acid,DHA)的药物进行早期干预,对T1DM的发生发展具有明显的预防和治疗作用。Stene等[7]在一项基于大规模人群的病例对照研究中报道,在出生后第一年饮食补充鱼肝油和维生素D可明显降低儿童期T1DM的发病率。越来越多的研究关注胰腺α细胞的重编程,因为它们在发育过程中与β细胞密切相关[7]。本课题组前期研究发现无论是mfat-1基因(内源性的将ω-6 PUFAs转化为ω-3 PUFAs)治疗或者ω-3 PUFAs饮食干预NOD小鼠,小鼠胰腺组织切片都显示了胰岛α细胞与胰岛β细胞共定位的现象[8]。mfat-1诱导的ω-3 PUFAs组织富集也被证明可以改善链脲佐菌素(STZ)诱导的高血糖症[9]。对mfat-1小鼠胰腺组织中ω-3PUFAs分析发现EPA的含量明显上调,揭示EPA在糖尿病的发生和发展中起着重要作用。但EPA的不同代谢物是否在介导胰腺细胞的转分化中起作用仍有待确定。已有研究发现[10],在人和大鼠胰腺中有大量的环氧二十碳三烯酸(EET)产生,这提供了CYP环氧合酶可能调节β细胞功能的潜在证据[11]。CYP450家族酶可以分解PUFAs产生环氧化物,随后通过可溶性环氧化物水解酶(sEH)代谢为相应的邻近二醇,二羟基二十碳三烯酸[12]。值得注意的是,源自EPA代谢物代谢的CYP 450途径的17(18)-EEQ和17(18)-DiHETE在mfat-1小鼠中急剧升高[14]。最近的一项研究还表明,DHA/EPA的膳食补充剂促进了大鼠胰腺中ω-3衍生的环氧二十碳水体烯酸(EEQ)的合成,例如17(18)-EEQ[14]。
本研究专注于 ω-3 PUFAs 饮食干预促进T1DM小鼠血糖和胰岛素分泌正常化的能力,进一步探讨了EPA代谢物,特别是来自CYP途径17(18)-EEQ和17(18)-DiHETE的代谢物,在促进胰岛α细胞向β细胞反式分化中的作用。
1 材料与方法
1.1 材料
1.1.1实验动物 SPF级雄性C57BL/6J小鼠(7周)购自江苏集萃药康生物科技有限公司。用于α细胞谱系示踪的GCGicre小鼠从caygen(B6; 129S-Gcgtm1.1(icre)Gkg/J(The Jackson laboratory stock #030663)购买,并与从上海南方模式生物科技股份有限公司购买的Rosa26tdTomato(B6;Cg-Gt (ROSA) 26Sortm9 (CAG-tdTomato) Hze/J (The Jackson laboratorystock #007909)杂交[15]。实验小鼠均饲养于华南理工大学大学城校区SPF级实验动物中心,使用许可证号:SYXK(粤)2017-0178。研究方案获得华南理工大学动物实验伦理委员会批准。
1.1.2主要试剂 STZ(S0130)和鼠抗glucagon(G2654)单克隆抗体购自 Sigma公司;兔抗insulin(4590s)单克隆抗体、兔抗Alexa Flour 488(4412s)荧光二抗和鼠抗Alexa Flour 555(4409s)荧光二抗均购自Cell Signal Technology公司;97%EPA乙酯(批号:02200201)购自成都国为生物医药有限公司;75%鱼油(EPA ∶DHA=25% ∶50%)购自上海贺普实业有限公司;小鼠胰岛素ELISA试剂盒(90080)购自Crystal Chem公司。
1.1.3仪器 气相色谱仪(gas chromatography,GC,美国Agilent);激光共聚焦(德国 Zeiss);酶标仪(瑞士Tecan);低温离心机(美国Thermo);石蜡包埋机(德国SLEE);冷冻台(中威电子仪器有限公司);石蜡切片机(美国Thermo)。
1.2 方法
1.2.1T1DM小鼠模型的建立与分组 7周龄野生型小鼠,腹腔注射溶于无菌柠檬酸缓冲液(pH 4.5,0.05 mmol·L-1柠檬酸钠)的STZ,剂量为60 mg·kg-1,连续给药5 d。1周后尾静脉血糖连续2周检测高于11.1 mmol·L-1确认为糖尿病小鼠模型。随后进行以下实验分组:STZ组、NC组(只注射柠檬酸缓冲液)、EPA组、鱼油组(Fish oil)。
1.2.2ELISA法检测小鼠血清中胰岛素水平 糖耐受实验过程中分别在0、30、60和90 min取小鼠尾静脉血,分离血清,按照小鼠胰岛素ELISA试剂盒说明书检测糖耐受过程中各时间点小鼠胰岛素水平。
1.2.3小鼠胰岛提取及培养 腹腔注射2.5%阿佛丁麻醉安乐死小鼠,向胰腺注射预冷的胶原酶P(1 g·L-1)(罗氏,11249002001,瑞士)。在38 °C下消化10 min,加入STOP溶液(含有10%胎牛血清的RPMI 1640),离心2 min(300×g)。将沉淀重悬于10 mL室温Hiatopaque 1100(Hiatopaque 1077和Hiatopaque1119 1 ∶1混合),再加入10 mL RPMI 1640,随后以离心20 min(900×g),收集组织和培养基界面的胰岛。胰岛在含有 10% 胎牛血清的RPMI 1640中培养(Gibco,美国),加入100 kU·L-1青霉素和 100 μg·L-1链霉素(Hyclone,美国)[16]。
1.2.4细胞培养 利用小鼠胰岛α细胞系αTC1进行体外培养。在细胞培养基中分别加入0.3 μmol·L-117(18)-EEQ和0.3 μ mol·L-117(18)-DiHETE,对照组加入相应浓度的酒精(EtOH),即分为EtOH组、17(18)-EEQ组、17(18)-DiHETE组。培养48 h之后,经过固定,利用荧光染色标记胰岛素(insulin)和胰高血糖素(glucagon),并使用荧光共聚焦显微镜观察记录。
1.2.5免疫荧光法检测细胞和小鼠胰腺组织中insulin与glucagon表达水平 将细胞系以5×105·L-1的密度接种,EPA代谢产物处理后,PBS洗去残留培养基,4%多聚甲醛固定30 min,PBS清洗后用1 ∶1 000的Triton-100室温下穿透20 min,清洗后用1%牛血清白蛋白室温下封闭1 h,吸去封闭液,加入一抗(1 ∶100 兔抗insulin,1 ∶1 000鼠抗glucagon)4 ℃孵育过夜,PBS清洗3次后,加入二抗(1 ∶1 000兔抗Alexa Flour 488,1 ∶1 000鼠抗Alexa Flour 555),室温避光孵育1 h,PBS清洗后滴加DAPI室温避光孵育5 min,PBS清洗,滴加抗荧光淬灭封片剂,在荧光显微镜下拍摄。
1.2.6脂肪酸的分离提取 将小鼠胰腺组织研磨后置于玻璃管中,加入4 mL甲醇(美国TEDIA,),2 mL氯仿(TEDIA)和1.5 mL水(TEDIA),涡旋;室温静置15 min;离心30 min(16 ℃,1 500×g);吸取下层液体转移至新的玻璃管中;氮吹仪吹干后加入1.5 mL 三氟化硼(上海阿拉丁),90 ℃水浴中孵育30 min;随后,室温静置10 min;加入4 mL戊烷(美国TEDIA),1.5 mL水,涡旋;离心2 min(16 ℃,1 500×g);转移上层液体至新的玻璃管中,放置于氮吹仪吹干,100 μL庚烷,涡旋混匀;混样品使用GC气相色谱仪检测。
2 结果
2.1 ω-3 PUFAs饮食干预对T1DM小鼠随机血糖和葡萄糖稳态的影响结果显示,与NC组相比,STZ组小鼠随机血糖水平明显升高,且随着造模时间的延长,血糖水平持续升高;而给予EPA和Fish oil饮食干预后小鼠血糖水平明显降低(Fig 1A),表明外源性ω-3 PUFAs对T1DM小鼠有明显的治疗作用。进一步通过葡萄糖耐量试验(injection glucose tolerance tests,IGTT)检测ω-3 PUFAs饮食干预对T1DM小鼠葡萄糖稳态的影响,STZ给药后第13周,分别向各组小鼠腹腔注射葡萄糖,监测血糖水平。结果显示,注射葡萄糖后各组小鼠血糖开始升高,从第15 min开始,EPA组和Fish oil组小鼠血糖明显低于STZ组(Fig 1B),且两组的曲线下面积(area under the curve,AUC)明显小于STZ组,虽然Fish oil组和EPA组随机血糖没有明显差异,但是EPA组相较于Fish oil组随机血糖有下降趋势,且EPA组的AUC明显低于Fish oil组,说明EPA在小鼠血糖的过程中可能起着更重要的作用,此结果也有待进一步的研究。以上结果显示ω-3 PUFAs明显改善T1DM小鼠对葡萄糖的吸收与利用,降低血糖总量(Fig 1C)。此外,无论是空腹状态,还是给予葡萄糖刺激,EPA组和Fish oil组小鼠血清胰岛素分泌水平均明显高于STZ组(Fig 1D),表明ω-3 PUFAs明显改善T1DM小鼠胰岛β细胞的胰岛素分泌功能。
2.2 ω-3 PUFAs饮食干预对T1DM小鼠胰腺中PUFAs含量和比例的影响利用气相色谱检测各组小鼠胰腺中各种PUFAs含量,结果如Tab 1所示,与STZ组相比,EPA组和Fish oil组小鼠胰腺组织中α-LA、EPA以及C22:5含量均明显增加(P<0.01);此外,EPA和 Fish oil干预明显降低胰腺组织中ω-6/ω-3 PUFAs的比例,为进一步探究ω-3 PUFAs防治T1DM的作用及其内在机制奠定了基础。
2.3 ω-3 PUFAs饮食干预对T1DM小鼠胰岛β细胞再生的影响为进一步探究ω-3 PUFAs饮食干预改善T1DM小鼠随机血糖和葡萄糖稳态的内在机制,利用免疫荧光技术观察各组小鼠胰岛分泌胰岛素与胰高血糖素功能的变化。结果显示,与STZ组相比,EPA组和Fish oil组小鼠部分胰岛细胞恢复了分泌胰岛素的功能,且存在可同时分泌胰岛素和胰高血糖素的细胞(Fig 2),表明ω-3 PUFAs饮食干预可促进胰岛α细胞转分化为β细胞,进而维持T1DM小鼠正常的血糖浓度以及糖代谢水平。
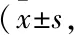
Tab 1 Analysis of ω-3 PUFAs and ω-6 PUFAs composition and rations in pancreatic samples of mice n=5)
Fig 1 ω-3 PUFAs normalized glucose metabolism in T1DM mice
2.4 tdTomato荧光蛋白特异性标记胰岛α细胞模型鼠的鉴定将GCGicre和R26TdTomato小鼠杂交,得到tdTomato荧光蛋白特异性标记胰岛α细胞的小鼠[16],剪取子代小鼠的脚趾提取DNA,通过聚合酶链式反应(polymerase chain reaction,PCR)对小鼠基因型进行鉴定,并区分野生型和GCGicre;R26TdTomato小鼠(Fig 3)。并通过qRT-PCR检测胰岛α细胞荧光标记小鼠的拷贝数,挑选高拷贝的荧光标记小鼠进行后续实验。
2.5 EPA代谢产物对胰岛细胞胰岛素分泌的影响提取GCGicre;R26TdTomato小鼠原代胰岛细胞,培养胰岛αTC1细胞系细胞,观察ω-3 PUFAs体外条件下对胰岛细胞功能的影响。结果显示,与单独STZ处理组相比,加入EPA代谢产物17(18)-EEQ或17(18)-DiHETE处理48 h后,两组小鼠原代胰岛细胞均可产生胰岛素(Fig 4),并且出现tdTomato荧光蛋白与胰岛素共定位的现象;同样,17(18)-EEQ或17(18)-DiHETE干预可引起胰岛αTC1细胞系细胞分泌胰岛素(Fig 5),表明EPA代谢产物在体外培养条件下可促进小鼠原代胰岛α细胞和αTC1细胞系细胞分泌胰岛素。
3 讨论
尽管T1DM的发病率和患病率存在很大地理差异,但仍然是很多国家中影响儿童和青少年健康的严重慢性病之一。β细胞的增殖发生在人类发育的早期,在成年人中几乎退化到检测不到的水平[17]。近年来,多项研究探索替代疗法在T1DM治疗中的应用,尝试应用干细胞、前体或分化细胞代替恶化的胰岛β细胞,并在体外模拟胚胎β细胞发育领域取得了重大进展,但仍需进一步分析β细胞发生和发育的遗传程序,进而能够产生完全分化的β细胞。基于此,产生胰高血糖素的α细胞及其转分化为β样细胞的能力在T1DM治疗领域具有很大的开发应用前景。
团队前期研究表明,营养干预或基因疗法(通过mfat-1基因)增加体内ω-3 PUFAs水平可以预防和逆转由Th1、Th2、Th17和Treg亚群失衡引起的自身免疫和糖尿病的发生,并存在胰腺α细胞转分化为胰岛素分泌细胞的迹象[8]。研究已证实,T1DM患者胰腺中仍有大量具有生理功能的α细胞[7],因此,α细胞可能在修复糖尿病患者β细胞损伤方面有巨大的潜在作用。本研究发现,EPA和鱼油饮食干预后T1DM小鼠胰腺中,以及EPA代谢产物作用后小鼠原代胰岛细胞或αTC1细胞中均出现明显的胰高血糖素与胰岛素共定位,表明ω-3 PUFAs饮食干预可促进胰岛α细胞向β细胞的转分化,从而使一些α细胞显示出“双重激素”状态,分泌胰岛素,增强机体对葡萄糖的吸收和利用,改善T1DM小鼠血糖水平和葡萄糖稳态。
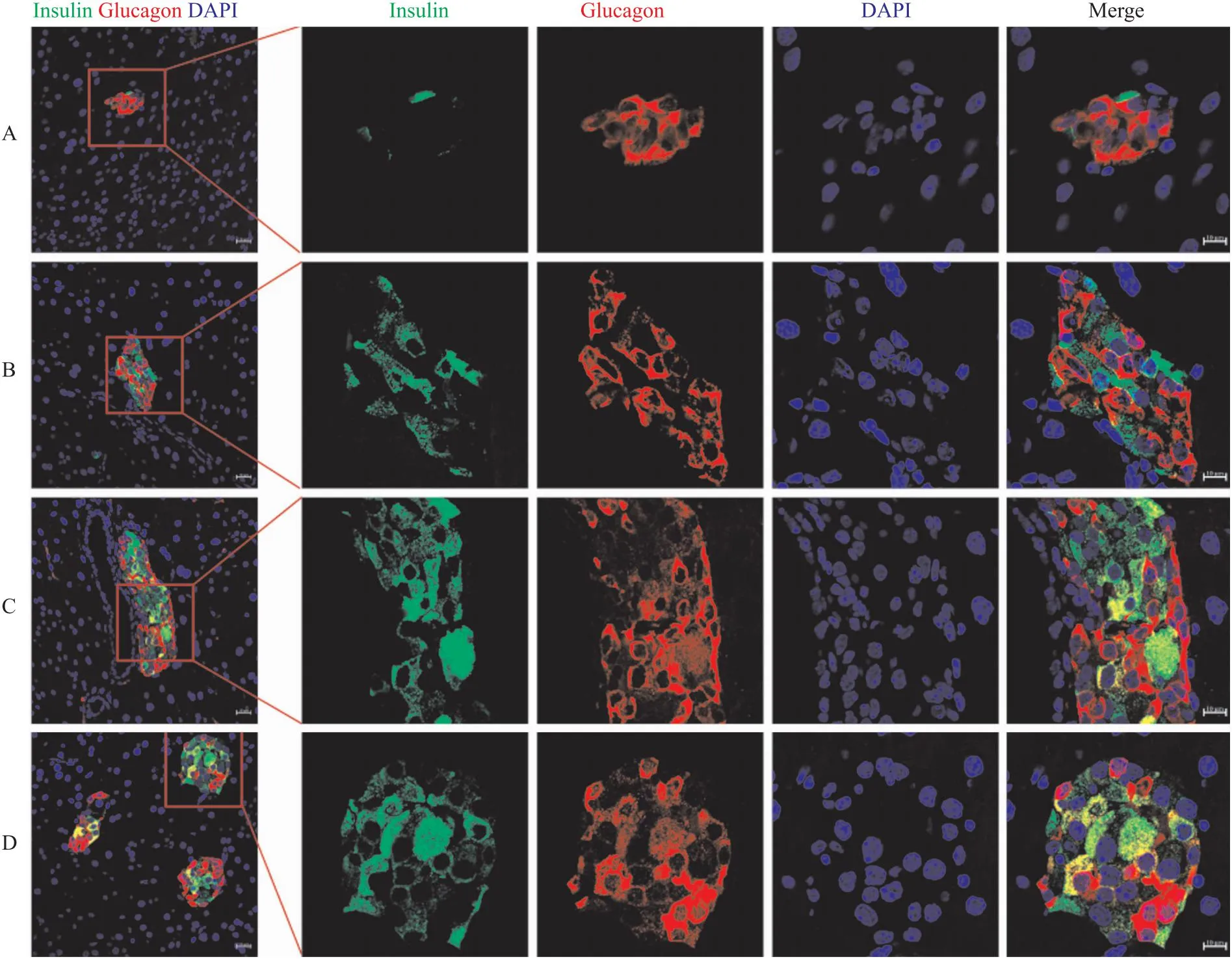
Fig 2 Islet and β cell regeneration and colocalization of α cells and β cells in diabetic mice treated with ω-3 PUFAs

Fig 3 Genotype of tdTOMATO fluorescent protein specifically labels produced in pancreatic α cell mice
综上所述,本研究首次证明ω-3 PUFAs饮食干预可通过促进胰腺α细胞向β细胞的转分化缓解STZ诱导的高血糖,尤其值得注意的是,EPA代谢物在胰岛不同细胞的转分化过程中发挥重要作用。本研究结果为T1DM的临床治疗提供了一种新的潜在干预策略。

Fig 5 Confocal images of αTC1 (insulin, green; glucagon, red; DAPI, blue)