异补骨脂素介导BMP2/Runx2/Osx信号通路促进成骨细胞增殖和分化的研究*
2024-01-22张有为黄廷锐唐德志
张有为,黄 晨,黄廷锐,唐德志**
(1.陕西省宝鸡市中心医院 宝鸡 721008;2.上海中医药大学附属龙华医院 上海 200032)
雌激素缺乏是绝经后妇女骨质疏松症(PMOP)的首要病因。对PMOP 的预防可采用雌激素替代疗法,但长期服用雌激素可明显增加患者发生癌症的危险性[1-2]。因此,寻找一种既有效又安全的雌激素类药物是当前本领域的研究热点。大量文献报道,植物雌激素可促进成骨细胞的增殖和分化,抑制骨吸收,诱导骨形成[3-6]。异补骨脂素属于呋喃香豆素类化合物,为植物雌激素之一[7]。现代药理研究发现,异补骨脂素具有促进骨代谢的作用,但其机制尚不清楚[8-9]。本实验旨在探讨异补骨脂素对成骨细胞增殖和分化的影响,并利用BMP2 体外条件性基因敲除技术从基因和蛋白的水平揭示其作用机理,为异补骨脂素用于临床治疗骨质疏松症提供全面、可靠的实验依据。
1 材料与方法
1.1 实验材料
1.1.1 实验细胞和实验动物
前成骨细胞株OCT1 细胞和BMP2loxp/loxp小鼠由美国罗切斯特大学骨科系陈棣教授馈赠。
1.1.2 实验药物
异补骨脂素标准品由上海药品检疫局购买,纯度达到95%以上;BMP2 纯化蛋白由浙江华东基因技术研究所购买,纯度超过90%。
1.1.3 实验试剂
胎牛血清(PAA 公司),a-MEM 干粉培养基(CALBIOCHEM 公司),0.25%含EDTA 的胰蛋白酶(PAA 公司),青-链霉素(PAA 公司),G-418 抗体(CALBIOCHEM 公司),二甲基亚砜DMSO(SIGMA 公司),MTT(SIGMA 公司),TRIZOL(晶美生物),Quant Reverse Transcriptase 逆转录试剂盒(TIANGEN 公司),高质纯化DNA胶回收试剂盒(GENMEN公司),兔抗鼠Runx2 单克隆抗体,兔抗鼠Osx 单克隆抗体,兔抗鼠β-actin 单克隆抗体和羊抗兔HRP 多克隆抗体(Cell signaling 公司),Ad-GFP 和Ad-Cre 腺病毒(上海吉凯生物公司)等。
1.1.4 实验设备
细胞培养箱(热能公司),SW-CJ-2FD超净工作台(AIRTECH 公司),倒置相差显微镜(Olympus 公司),倒置相差显微镜(Olympus 公司),AF100 制冰机(SCOTMAN 公司),高速离心机(Eppendorf公司),MK-Ⅲ型酶标仪(Dynex 公司),DU 800/VIS 紫外分光光度计(Beckonman 公司),Rotor Gene 3000 荧光PCR 仪(基因公司),Odyssey远红外扫描仪(LI-COR公司)等。
1.2 实验方法
1.2.1 细胞复苏培养
取出装有OCT1细胞的冻存管,按常规方法进行细胞活性复苏,计数细胞,按1×105·mL-1的密度接种于无菌培养瓶中,置于37°C、5% CO2培养箱中培养。次日更换1 次培养液后再继续培养,以后每3-4 天换1 次液。细胞培养液采用含10%胎牛血清、100 U·mL-1青-链霉素和300 μg·mL-1的G-418抗体的a-MEM培养基。
1.2.2 分组方法
①正常对照组,普通培养液;②异补骨脂素低剂量组,浓度为10 μg·mL-1;③异补骨脂素中剂量组,浓度为30 μg·mL-1;④异补骨脂素高剂量组,浓度为60 μg·mL-1;⑤阳性对照组,BMP2纯化蛋白50 ng·mL-1。
1.2.3 MTT法观察细胞增殖情况
取经常规胰蛋白酶消化后的传代细胞,计数细胞,调整细胞密度为1×104·mL-1左右。将细胞移入一块96孔培养板中,每孔200 mL,然后放入37℃ 5% CO2培养箱中培养。培养24 h 后,更换无血清培养液,使细胞生长同步化。于第2 天吸去孔内的培养液,按上述分组方法分别给药,每组8个复孔,每孔200 μL含药无血清细胞培养液。之后,再放入37℃ 5% CO2培养箱中继续培养。于培养48 h后,取出培养板进行MTT 法检测。先往每孔加入20 μL MTT 溶液(5 mg·mL-1),放入37℃ 5% CO2培养箱中孵育4 h 后,弃去上清液,每孔加入DMSO 180 μL,充分震摇混匀,使结晶完全溶解,立即上酶标仪测定OD 值,波长为490 nm,以同时做的单纯培养液为空白对照孔。
1.2.4 实时荧光定量RT-PCR反应
细胞加药培养48 h 后取出,分别倒尽培养液,采用TRIzol法提取细胞总mRNA。提取后用紫外分光光度计选择波长260 nm 和280 nm 检测样本OD 值(即光吸收值),以超纯水作为空白对照调零,测得样本A260/A280 比值在1.8-2.0,证明RNA 抽提成功,并用1%的琼脂糖凝胶进行电泳,以检测总RNA 的完整性。逆转录后进行进行两步法扩增。PCR 反应体系为20 μL,扩增仪中95℃ 10 min,再扩增(95℃变性20 s,62℃退火5 s,72℃复性25 s)40 个 循环,72℃延 伸10 min。制备标准曲线后,用Rotor Gene6.0 软件自动进行绝对定量分析,以每一样体所含目的基因的拷贝数和其β-actin 内参基因的拷贝数的比值表示目的基因的表达量。其引物序列及参照物β-actin 引物序列由大连宝生物公司合成,引物序列见表1。
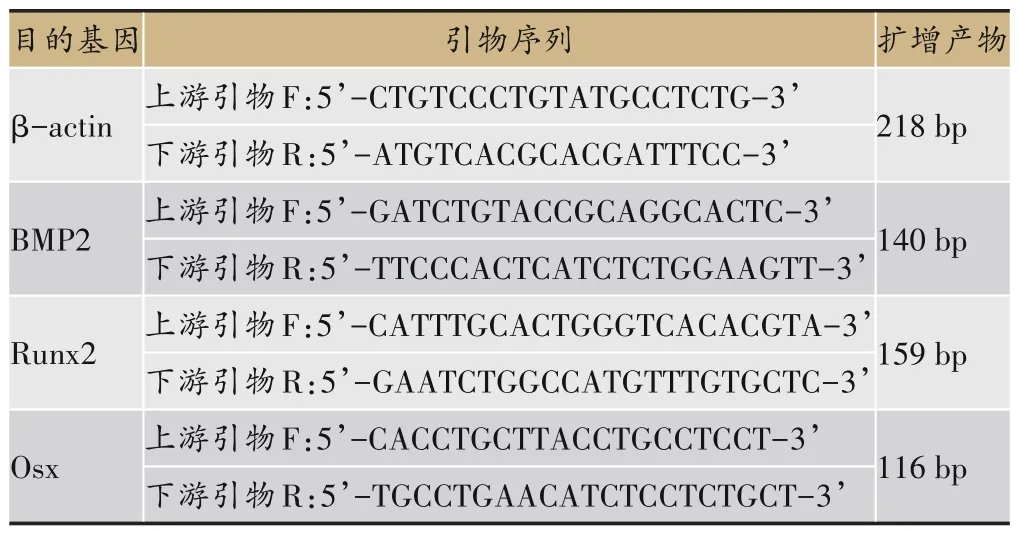
表1 Real time RT-PCR检测所检测的基因引物序列
1.2.5 Western blot检测
细胞加药培养48 h 后取出,分别倒尽培养液,裂解细胞,提取蛋白并定量。将每组蛋白溶液调整至相同浓度,经煮沸变性后,在SDS-聚丙烯酰胺凝胶上电泳。电泳结束后,将凝胶上的蛋白转移至PVDF 膜上,采用5%脱脂奶粉封闭1 h,振荡漂洗干净后,分别加入兔抗鼠BMP2、Osx 和β-actin 单克隆一抗,4℃过夜。次日取出,振荡漂洗干净后,加入羊抗兔HRP 多克隆二抗,孵育1 h,振荡漂洗干净后,在Odessey 远红外扫描仪上检测蛋白条带,保存图像。
1.2.6 BMP2体外条件性敲除实验
从出生3 天的BMP2loxp/loxp小鼠中分离原代颅骨成骨细胞,然后以1×106个·mL-1的密度接种到六孔培养板上进行培养。利用A d-GFP 或Ad-Cre(病毒滴度:4×108pfu·mL-1)感染2 天后,用或不用异补骨脂素(最佳浓度)分别干预48 h,部分细胞用来抽提总RNA,逆转录成cDNA,利用Real-time PCR 反应检测BMP2、Runx2 和Osterix(Osx)等基因表达,操作方法和步骤同前所述。部分细胞用10%福尔马林固定,加入含0.5 mg·mL-1对硝基苯磷酸的AMP 缓冲液(0.5 mol·L-1甲基二戊烷,2-丙胺醇,2 mmol·L-1氯化镁,pH=10.3)染色30 min,直到颜色变化以检测碱性磷酸酶活性。
1.2.7 统计分析
计数指标以均数±标准差(±s)表示。借助SPSS 11.0 软件包,多样本之间进行One-Way ANOVA 分析,两两样本之间进行t检验。检验水准取双侧α=0.05。
2 结果
2.1 形态学观察结果
倒置显微镜下观察,在给药培养48 h 后,各组细胞的数量及形态与正常组相比均无明显异常。见图1。
图1 大体观察(×10)
2.2 MTT法检测结果
在给药培养48 h后(见表2),异补骨脂素能促进成骨细胞的增殖,但以低剂量(10 μg·mL-1)组最明显(P<0.05),而中剂量(30 μg·mL-1)组和高剂量(60 μg·mL-1)组作用不明显(P>0.05);阳性对照组BMP2 纯化蛋白亦能明显促进成骨细胞的增殖(P<0.05)。
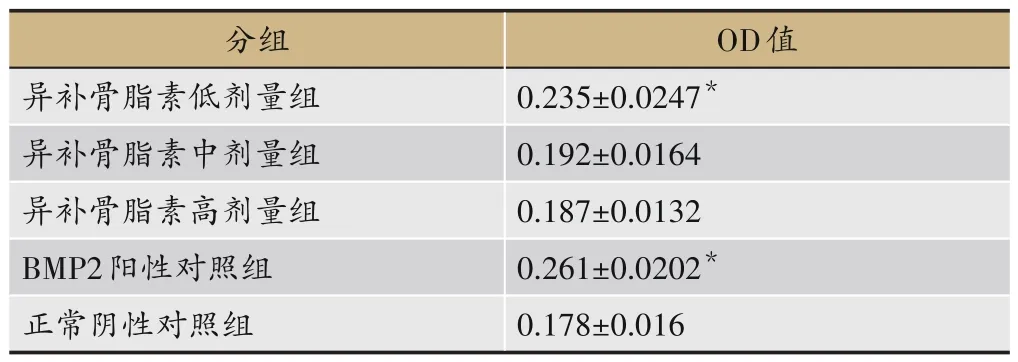
表2 成骨细胞增殖情况
2.3 实时荧光定量PCR反应结果
结果显示(见表3),异补骨脂素能促进成骨细胞BMP2 mRNA 的表达,以低剂量组(10 μg·mL-1)最明显(P<0.05),中剂量组(30 μg·mL-1)次之(P<0.05);阳性对照组BMP2 纯化蛋白亦可以显著增加成骨细胞BMP2 mRNA 的表达(P<0.05)。异补骨脂素低剂量(10 μg·mL-1)和 中 剂 量(30 μg·mL-1)能 明 显 促 进成骨细胞Runx2 mRNA 的表达(P<0.05),而高剂量(60 μg·mL-1)组成骨细胞Runx2 mRNA 表达量比正常对照组偏低,但无统计学意义。阳性对照组BMP2 纯化蛋白亦可以显著增加成骨细胞BMP2 mRNA 的表达(P<0.05)。异补骨脂素低剂量组(10 μg·mL-1)和中剂量组(30 μg·mL-1)能明显促进成骨细胞Osx mRNA 的表达(P<0.05),而高剂量(60 μg·mL-1)组成骨细胞Osx mRNA 表达量比正常对照组偏低,但无统计学意义。阳性对照组BMP2纯化蛋白亦可以显著增加成骨细胞BMP2 mRNA的表达(P<0.05)。
2.4 Western blot检测结果
结果显示(见图2),异补骨脂素低剂量(10 μg·mL-1)和中剂量(30 μg·mL-1)能增加成骨细胞中Runx2蛋白的表达,分别提高了2.18倍和1.25倍;阳性对照BMP2 纯化蛋白也能增加成骨细胞中Runx2 蛋白的表达,提高了2.75倍。
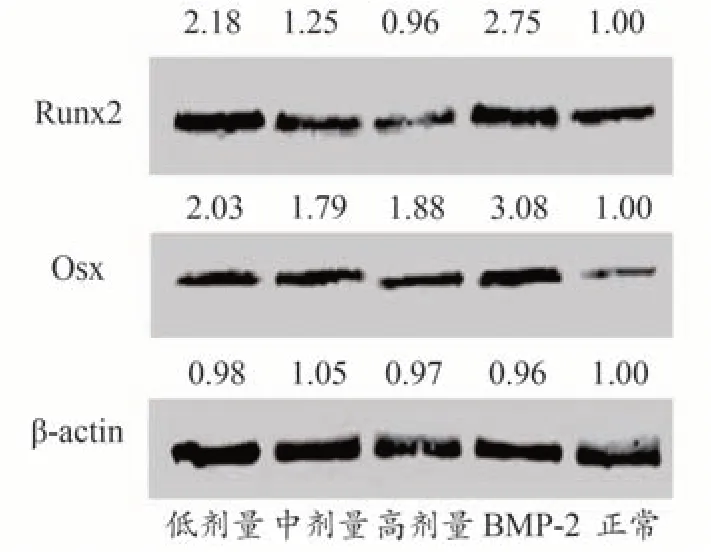
图2 各组Runx2、Osx蛋白表达量的比较
异 补 骨 脂 素 低 剂 量(10 μg·mL-1)、中 剂 量(30 μg·mL-1)和高剂量(60 μg·mL-1)均可提高Osx 蛋白的表达量,分别提高了2.03 倍、1.79 倍和1.88 倍;阳性对照BMP2 纯化蛋白也能增加成骨细胞中Osx 蛋白的表达,提高了3.08倍。
2.5 BMP-2体外条件性敲除实验结果
为进一步研究异补骨脂素诱导成骨细胞分化的作用机理,从Bmp2loxp/loxp小鼠中分离培养颅盖骨成骨细胞,用Ad-Cre 慢病毒体外将BMP2 基因敲除(Ad-GFP做为野生型对照)。Real-time PCR 反应检测结果显示,经Ad-Cre 慢病毒干预后成骨细胞中BMP2 的表达显著降低,BMP2 基因敲除效率达80%以上,说明BMP2基因敲除成功,如图3所示。之后分别给予最佳浓度(10 μg·mL-1)的异补骨脂素干预,结果发现异补骨脂素可明显促进Ad-GFP 处理的野生型对照组成骨细胞中Runx2(图4)和Osx(图5)基因的表达,分别提高了7.2倍和5.9倍。但这种作用在BMP2缺失的成骨细胞中部分消失,仅仅提高了4.6 倍和3.2 倍。通过碱性磷酸酶(ALP)染色,发现异补骨脂素可明显促进Ad-GFP 处理的野生型对照组成骨细胞中ALP 染色强表达,而这种作用在BMP2 缺失的成骨细胞中部分减弱(图6)。上述结果均表明异补骨脂素促进成骨细胞分化具有BMP2部分依赖性。
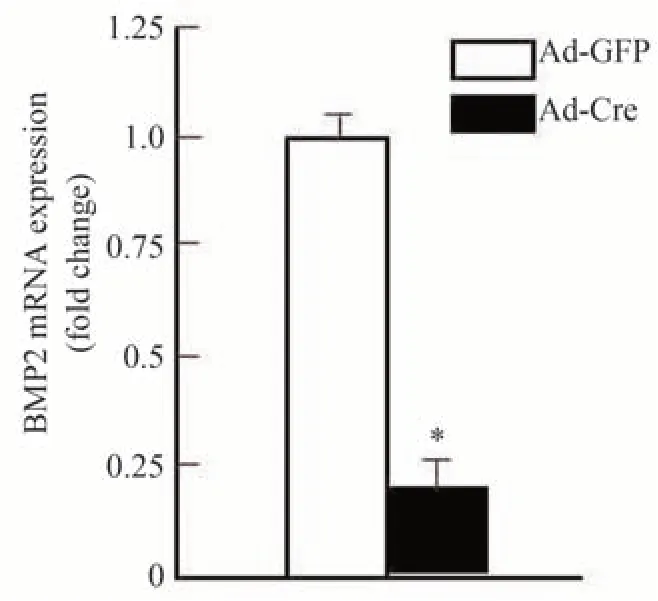
图3 BMP2基因敲除效率
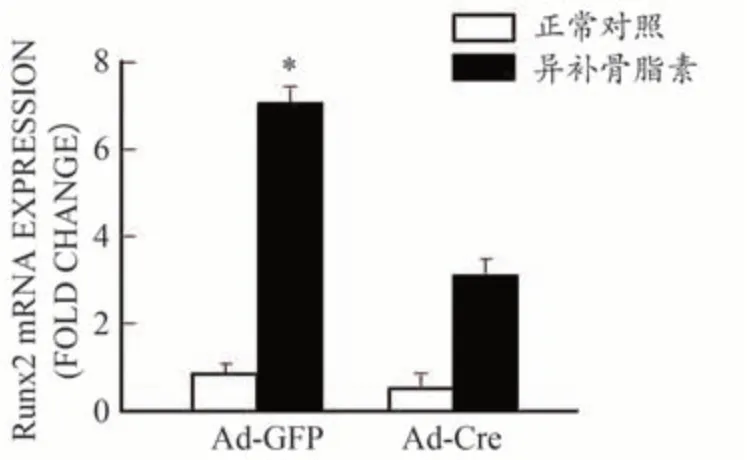
图4 异补骨脂素促进Runx2基因表达部分依赖于BMP2
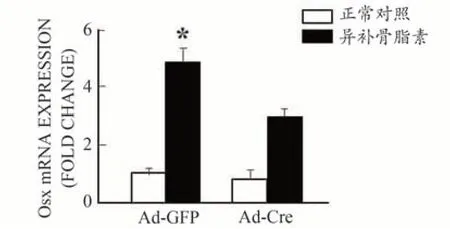
图5 异补骨脂素促进Osx基因表达部分依赖于BMP2
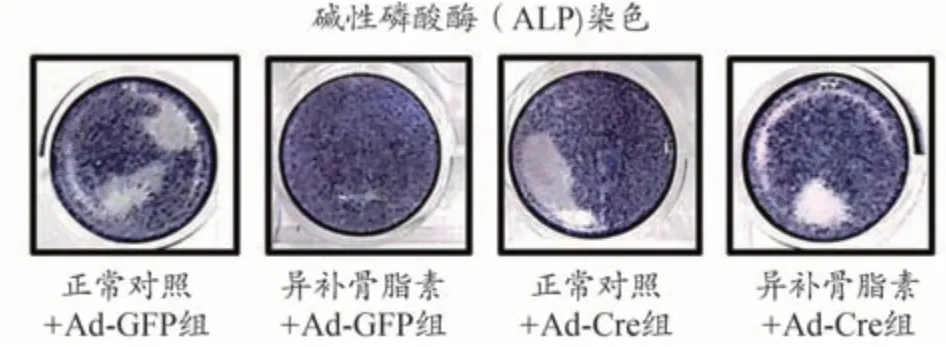
图6 异补骨脂素促进ALP表达部分依赖于BMP2
3 讨论
成骨细胞在骨代谢的体外研究中非常重要,为研究具有刺激骨形成药物及作用机理提供重要的细胞模型。动物成骨细胞一般来源于胎鼠或新生鼠的颅骨,这些细胞所处的分化时期不同,性质不完全相同[10]。本实验中,选用的OCT1细胞株是从过表达骨钙素转基因小鼠的颅盖骨中分离获得的成骨细胞,生长旺盛、性质稳定,可以大量满足实验研究的需要[11]。
中医学认为,肾主骨生髓,一旦肾气衰微,肾精不足,则会发生骨质疏松,正如《素问·痿论》中所说“骨枯髓减,发为骨痿”。肾中精气宜充不宜亏,治疗骨质疏松症以补肾益精为主,代表方剂如肾气丸、左归丸、右归丸等,药物有仙灵脾、补骨脂、熟地、骨碎补、杜仲、枸杞、牛膝等[12-13]。实验研究发现,这些补肾方药可以提高成骨细胞的增殖率、增加ALP 活性,因而促进成骨细胞骨形成功能[14-17]。但对于这些补肾方药中的具有促进成骨细胞增殖的有效组分尚缺乏系统的研究。通过本实验发现,补骨脂中的有效组分异补骨脂素能明显促进成骨细胞的增殖,并以10 μg·mL-1浓度作用最强。
1965 年,Urist[18]发现在脱钙的骨基质中存在着一种特殊的蛋白质,能诱导异位成骨,后来就被命名为骨形态发生蛋白(Bone morphogenetic protein,BMP)。迄今为止发现了20 多种BMP,除BMP1 外,其他BMPs均属于TGF-β 超家族成员,可通过旁分泌和自分泌的形式诱导骨、软骨及与骨有关的结缔组织的形成,还能诱导骨源和非骨源性细胞的成骨分化[19]。在成骨细胞中,BMPs 信号转导途径对骨形成起至关重要的作用。BMP2被认为是TGF-β超家族中活性最高且能单独诱导成骨的因子,其高表达可以促进成骨细胞的分化[20-21]。通过本实验发现,异补骨脂素能促进BMP2的表达,从而诱导成骨,并以低剂量(10 μg·mL-1)及中剂量(30 μg·mL-1)作用较强,而高剂量(30 μg·mL-1)作用较弱。
Runx2属于Runt结构域基因家族成员,是与BMPs信号转导的下游分子Smad1 和Smad5 相互作用,从而调节成骨细胞的分化[22-24]。在没有BMP2 作用时,Runx2 单独表达虽然能抑制成肌细胞分化,但不能上调骨钙素的表达,而在BMP2 作用下,过表达Runx2 诱导产生的骨钙素水平高于BMP-2 的单独作用,说明BMP2 和Runx2 具有协同作用。以上结果均表明Runx2 在BMP-2 激活成骨的信号途径中起了非常重要的作用[25-26]。Runx2 是成骨细胞分化所必需的关键转录因子,可协同其下游的Osx 调节成骨细胞特异性基因的表达,从而影响成骨细胞分化[27-28]。通过本实验,发现异补骨脂素可以诱导Runx2 mRNA 和Osx mRNA 的高表达,并以低剂量(10 μg·mL-1)和中剂量(30 μg·mL-1)作用最强。
条件性基因敲除(Conditional gene knockout)是指为将某个基因的修饰限制于小鼠某些特定类型的细胞或发育的某一特定阶段的一种特殊的基因敲除方法[29]。实际上是在常规的基因敲除的基础上,利用重组酶介导的位点特异性重组技术,在对小鼠基因修饰的时空范围上设置一个可调控的“按钮”,从而使对小鼠基因组的修饰的范围和时间处于一种可控状态。目前广泛应用方法就是利用Cre/LoxP 重组酶系统来研究小鼠特定组织器官或特定细胞中靶基因灭活所导致的表型。Cre 重组酶是一种位点特异性重组酶,能识别并介导两个LoxP 位点(序列)之间的特异性重组,使LoxP 位点间的基因序列被删除或重组[30]。如果将“LoxP Floxed”小鼠的细胞转染病毒介导的Cre 重组酶,可产生靶基因发生特定方式修饰的条件性敲除细胞,这是一种体外条件性基因敲除方法。在“LoxP Floxed”小鼠或细胞,正常情况下同野生型的一样,只有遇到Cre 重组酶时才会导致靶基因的突变[31]。本实验为进一步验证异补骨脂素通过介导BMP2 增加Runx2 和Osx 表达而诱导成骨细胞分化的作用机理,从BMP2loxp/loxp小鼠中分离培养颅盖骨成骨细胞,用Ad-Cre 慢病毒体外将BMP2 基因敲除,然后给予低剂量(10 μg·mL-1)异补骨脂素干预,发现异补骨脂素诱导的Runx2 和Osx 高表达呈BMP2 的依赖性。因此,本实验结果首次证明了异补骨脂素通过促进BMP2/Runx2/Osx 信号通路的激活,从而诱导成骨细胞分化。
