Quiet in the nest: The nest environment attenuates song in a grassland songbird
2024-01-22SrhDoneyMikylBornisRynNorrisAmyNewmnHetherWillimsStphnieDouetDnielMennill
Srh L.Doney, Mikyl M.K.Bornis, D.Ryn Norris, Amy E.M.Newmn,Hether Willims, Stéphnie M.Douet, Dniel J.Mennill
aDepartment of Integrative Biology,University of Windsor,Windsor,Ontario,N9B 3P4,Canada
bDepartment of Integrative Biology,University of Guelph,Guelph,Ontario,N1G 2W1,Canada
cBiology Department,Williams College,Williamstown,MA,01267,USA
Keywords:Acoustic environment Nest environment Songbird Sound attenuation Vocal learning
ABSTRACTThe nest environment may limit the ability of nest-bound birds to hear sounds from the outside world.In vocal learning species, such as humans and songbirds, it is vital for young animals to hear the voices of conspecific animals early in life.In songbirds, nest structure varies considerably across species, and the resulting impact on sound transmission may have consequences for vocal learning in nestlings.In this study, we tested the hypothesis that the nest environment attenuates song of Savannah Sparrows (Passerculus sandwichensis), grassland songbirds that build nests on the ground under cover of dense vegetation.We recorded live Savannah Sparrows singing at variable distances from 21 nests.We recorded songs using one microphone inside the nest (the typical position of a nestling) and another placed 1 m directly above the nest (a typical position of an adult).We found a substantial reduction in signal-to-noise ratio, where songs recorded inside the nest were an average of 11 dB lower than songs recorded directly above the nest.We estimate that the attenuation imposed by the nest reduced the maximum acoustic environment from 117.7 m (for recordings above the nest), to 78.6 m (for recordings within the nest), which is analogous to listening from a position 39.1 m farther away from the singer.Previous research estimated that song transmits up to 100 m in this species, so any adult male within 100 m of a young bird was previously considered a potential vocal tutor.By reducing the nestling acoustic environment from 100 m to 78.6 m, the number of male tutors available to nestlings is reduced by an average of 27%.Given the growing evidence that song learning begins very early in life, future research on vocal development should account for how the structural properties of the natal environment affect the songs that nestlings hear.
1.Introduction
Nest-bound birds listen to the outside world from within the nest.Females listen to songs of potential mates during the laying period(Mennill and Ratcliffe, 2004); parents listen to coordination signals of their mates during the incubation and provisioning periods (Boucaud et al., 2017; Ferree et al., 2021); and nestlings listen to signals of their parents during the nestling period (Platzen and Magrath, 2004).For birds that exhibit vocal learning, a behaviour that has evolved independently in several animal taxa, including songbirds, parrots, and hummingbirds (Nowicki and Searcy, 2014), young birds listen to the songs of tutors from inside the nest.Young birds deprived of exposure to adult song learn crude approximations of typical song (Marler and Peters, 1987), yet little is known about the importance of early exposure to adult song for vocal learning.
Historical studies of songbird vocal learning were conducted in lab settings on post-fledging individuals (animals >10 days old; Marler,1970).However, there has been a recent shift to investigations of vocal development at the embryonic and nestling stages (Kleindorfer et al.,2018; Katsis et al., 2018; Rivera et al., 2019; Hudson et al., 2020), when young birds are limited to listening to tutors from inside the nest.In Zebra Finches (Taeniopygia guttata), embryos and nestlings can discriminate between conspecific and heterospecific song (Rivera et al.,2019).Furthermore, vocalizations heard by Zebra Finch embryos influence song learning and reproductive success later in life (Katsis et al.,2018).Nestling Golden-crowned Sparrows (Zonotrichia atricapilla)discriminate between conspecific song and song of a closely related,sympatric sister species (Hudson et al., 2020).In Superb Fairy-wrens(Malurus cyaneus) prenatal sound discrimination appears to lead to greater vocal complexity in adulthood (Kleindorfer et al., 2018).A recent experiment altering song type exposure in nestling Swamp Sparrows (Melospiza georgiana) provides evidence that nestling acoustic experience can improve sound discrimination (Schroeder and Podos,2023).Collectively, these lines of evidence from diverse species of songbirds highlight an emerging theme of the importance of the early vocal learning period.
The structure and environment of bird nests may obstruct sound and limit the acoustic environment of nest-bound birds (Blumenrath et al.,2004; Lampe et al., 2004; Mennill and Ratcliffe, 2004; Grabarczyk and Gill, 2019).In the context of song learning, during the earlier days or weeks of life, songbird nestlings are confined inside nest structures built to shield them from weather and predators (Collias, 1997).It is worthwhile to consider how the physical environment of nestlings influences the sounds that young birds hear.In this study, we investigate the acoustic environment of nestling Savannah Sparrows (Passerculus sandwichensis) which live in the open grassland habitats found on Kent Island, New Brunswick, Canada, in the Bay of Fundy.In this population,vocal learning begins in the natal summer, specifically within the first weeks of life (Mennill et al., 2018).In other sparrow species, the early song learning period was previously estimated to occur between 10 and 60 days post hatch (Marler and Peters, 1987), and Savannah Sparrows have traditionally been assumed to follow a similar timeframe (Wheelwright et al., 2008).Savannah Sparrows are known to learn their songs during a sensitive period during their first year, and males produce only one song type for their entire adult lives (Wheelwright and Rising, 2020;Thomas et al., 2021).Previous studies on the Kent Island population suggest that most individuals produce a song learned not from their social father, but from other adult males in neighbouring territories during their natal year or first breeding year (Wheelwright et al., 2008).A more recent experiment in the wild demonstrates that it is vital for tutor songs to be heard during both the natal summer and again at the onset of the first breeding season (Mennill et al., 2018).Male song rate in this population decreases significantly throughout the breeding season,dropping to the lowest rate during the fledgling period (Moran et al.,2019).Therefore, most songs heard during the sensory period for song learning are likely heard during the nestling phase.Previous research estimated that male song transmits up to 100 m from the signaler(Wheelwright et al., 2008); however, females construct nests on the ground under cover of dense, grassy vegetation which may attenuate sounds for individuals inside the nest, reducing their hearing range to closer distances, although this has never been studied empirically.
To test the hypothesis that the nest environment attenuates song, we recorded male Savannah Sparrows singing at various song posts around nests using a pair of microphones: one microphone inside a nest (the typical position of a nestling) and another placed directly above the nest,1 m high (the typical observed singing position of adults from song posts at the top of vegetation).We predicted that songs recorded from within nests would have lower signal-to-noise ratios than songs recorded above nests.Given that Savannah Sparrow nests are surrounded by varying amounts of vegetation, we also predicted that songs recorded from more densely covered nests would have lower signal-to-noise ratios than songs recorded from more open nests.We were motivated by a desire to develop a deeper understanding of the nestling acoustic environment,and thereby improve future investigations of vocal learning.
2.Materials and methods
2.1.Data collection
We collected recordings of a long-studied population of Savannah Sparrows at the Bowdoin Scientific Station on Kent Island, New Brunswick, Canada (44◦35ʹ N, 66◦46ʹ W; Dixon, 1978; Burant et al., 2022).Our study area consisted of an open, grassy, 10 ha field in the central part of the island, surrounded by a spruce-fir forest to the north and west, and a tidal beach to the east.The field continues relatively undisturbed to the south tip of the island, however the annually monitored study area only includes the northern 10 ha of this field (Dixon, 1978;Burant et al., 2022).Each year between May and late July, all individuals in this population are marked with unique combinations of coloured leg bands, all nests are located, and individuals are monitored daily.Territory locations are mapped onto a scaled map of the study site,based on observations of pair locations, singing locations, and nest locations throughout the breeding season (Burant et al., 2022).As part of ongoing investigations of song in this population, we record the songs of all males present each breeding season, while observing coloured leg bands (Hensel et al., 2022).Because birds were individually marked,and individuals are highly territorial, we are confident that all male songs were recorded each year.The study area held 43 breeding males in 2021 and 37 breeding males in 2022.
We studied sound transmission into the nest by collecting recordings within 21 nests (11 nests in 2021 and 10 nests in 2022).We sampled nests between July 11–29 in 2021, and between June 19–25 in 2022 (the onset of the breeding season was much earlier in 2022 compared to 2021).To ensure we did not disturb breeding pairs, we collected recordings soon after nestlings had fledged from the nests.We only used nests where young successfully fledged and did not collect recordings at nests that were depredated, since the predators might have damaged or changed the nest structure.We collected recordings at nests from across the entire study area.The average distance between sampled nests was 142.7 m in 2021 and 157.6 m in 2022 (Appendix Fig.S1).We collected all recordings in good weather conditions when there was no rain or heavy wind.We collected recordings between 09:00 and 12:00, unless no adults sang nearby, in which cases we repeated those recordings at dawn or dusk when male song output was higher (Moran et al., 2019).
We recorded adult males while they spontaneously sang at various distances from the nest.We sampled recordings in stereo with an automated digital recorder (Fig.1; Wildlife Acoustics Song Meter SM2;44.1 kHz sampling frequency; 16-bit accuracy; WAV format).We mounted one of the microphones inside the nest, on the ground, at a typical nestling position; we mounted the other microphone directly above the nest at a typical singing height of an adult bird (~110 cm high;Fig.1).Both microphones faced the same direction.Between subsequent recording locations, we alternated which microphone we positioned above the nest and within the nest, and which microphone we connected to the left and right channel of the recorder, to avoid any influence of variation in microphone sensitivity.

Fig.1.Diagram of our study design, showing a stereo recorder deployed at a Savannah Sparrow nest with one microphone mounted 1 m above the nest and a second microphone mounted inside the nest.Two birds are illustrated producing songs at different distances from the nest.
After deploying a recorder at a nest site, we stepped back 5 m and waited for nearby adult males to sing.Upon hearing a male sing, we noted the exact time, the singing location of the male, and the singer’s identity if possible (by visually observing coloured leg bands).After the male finished singing, we approached his song post and used a handheld Global Positioning System (GPS) unit (Garmin model IPH-02401)to collect coordinates of his song post.We aimed to record at least one song from multiple males at various distances from each nest; however,we ceased recording at nests after 1 h, regardless of the number of songs recorded.Lastly, we used the GPS unit to collect coordinates of the nest location.We obtained locations of singers and nests in an open environment with a clear view of the sky, with a GPS accuracy of <3 m.We used QGIS software to map all nest locations and male singing locations(we used the EPSG: 2953 Coordinate Reference System for New Brunswick, Canada).We used the distance matrix algorithm function of QGIS to calculate the exact distances between each singer and the nest he was recorded from.
To evaluate whether sound attenuation was influenced by vegetation density around the nest, we quantified vegetation density surrounding the 10 nests that we recorded in 2022.We created a 7.5 cm diameter disk (the average outer diameter of Savannah Sparrow nests; Wheelwright and Rising, 2020), and coloured it with a checkerboard pattern made of orange and black 1 cm-by-1 cm squares.We placed this disk into each nest and stood directly above the nest to count the number of disk squares visible through the vegetation.We then divided the number of obstructed squares by the total number of disk squares to determine the proportion of the nest that was concealed from directly above.We then multiplied this number by 100 to obtain a percentage of the nest that was concealed, as a measure of nest cover (as in Holmes et al., 2020).We used visual concealment as a proxy for acoustic concealment.In addition, we determined the dominant plant species at each of the 10 nests measured in 2022, to evaluate whether sound attenuation was influenced by the plant species.We determined the dominant plant species by visually observing the most abundant species immediately covering each nest.
2.2.Song analysis
We annotated recordings in Syrinx PC sound analysis software (J.Burt, Seattle, WA, U.S.A.), viewing both recorder channels as stereo sound files.We isolated instances when Savannah Sparrow song was detected by at least one microphone.Some adult males sang repeatedly from the same location during a recording; however, we used only one song per individual for each unique song post.We extracted stereo recordings of songs from both microphone positions in Syrinx, and measured amplitudes in Avisoft SAS Lab Pro (R.Sprecht, Berlin, Germany).We measured the amplitude of four common note types in Savannah Sparrow songs (the “i” note from the introduction; the “ch”note or “dash” note from the middle segment; and the “is” note from the trill segment; terminology from Williams et al., 2013).For each note, we measured both the amplitude of the note and the amplitude of background noise, obtained from a 0.05 s silent interval immediately before or after the note.In 12% of the notes we measured, we could not accurately measure a silent interval immediately before or after, either because of overlapping background noise, or because the silent interval near the measured note was too short.In these cases, we calculated the average noise amplitude from two 0.05 s intervals immediately before and after the song.We used the Automated Parameter Measurements feature of Avisoft, applying an amplitude threshold of 10 dB relative to the maximum, an FFT length of 512, and a frame size of 93.5%, to measure each note and noise interval from the waveforms, while viewing spectrograms of each song.In each case, Avisoft automatically measured the amplitude every 0.006 s throughout the note or noise duration.For notes, we selected only the note of interest to calculate amplitude, ensuring that Avisoft only targeted the desired note, and avoided any background sounds.For noise intervals we selected 0.05 s interval immediately before measured notes, and Avisoft measured amplitude across that entire period.In cases where low frequency sounds were visible on the spectrogram but did not overlap in frequency with the Savannah Sparrow song, we applied a high-pass filter to remove the low frequency sounds, using the same filter settings for each pair of note and noise measurements.After measuring note and noise amplitude, we calculated signal-to-noise ratio by subtracting the noise amplitude from the note amplitude (as in Halfwerk et al., 2012;Rehberg-Besler et al., 2016).
2.3.Statistical analyses
We analyzed 68 Savannah Sparrow songs from 21 nests (32 songs from 11 nests in 2021; 36 songs from 10 nests in 2022).Overall, we analyzed an average of 3.2 ± 1.6 individual songs per nest site (range 1–7).These songs were produced by 26 different males throughout the study area (6 males recorded in only 2011; 15 males recorded in only 2022; 5 males recorded in both years).Overall, we analyzed an average of 2.5 ±1.4 males per nest site (range 1–6).Male identity was confirmed by observing coloured leg bands in the field and through comparisons of spectrograms in our library of recordings of all males in the population.The social father was recorded at the nest site for 7 nests, whereas only neighbouring males were recorded singing at the other 14 nests.Some of the 26 singers were recorded repeatedly at different nests or at different singing positions from a nest; given that our unit of replication is a song produced at a specific distance from the nest, and the comparison of how that song transmits to a position above the nest and within the nest, we considered each song post to be an independent datapoint.We confirmed that this decision did not influence our results by randomly selecting one song for each male and running our analyses again; the patterns did not change.Within the full songs, we measured 184 notes of four common note types.Given that we measured the signal-to-noise ratio of multiple notes for each song, we calculated the average signalto-noise ratio of all notes within each song to account for any differences across note types.
Our measurements of signal-to-noise ratios of above-nest and withinnest recordings, bird distance, and nest cover were not normally distributed (Shapiro-Wilks test:p< 0.05), and therefore, we used a Wilcoxon signed-rank test to conduct a pair-wise comparison of abovenest and within-nest signal-to-noise ratios.We report the mean and standard deviation for each microphone position and present the effect size of this comparison using standardized mean difference (or Cohen’sd).In addition, we conducted separate post hoc analyses for each note type measured (“i”, “ch”, “dash”, or “is”) again using Wilcoxon signedrank tests to compare signal-to-noise ratios for each note type, to evaluate whether the general pattern remained across note types.We conducted a multivariate linear regression to compare above-nest and within-nest signal-to-noise ratios; our dependent variables were the recorded average signal-to-noise ratios and our independent variables were distance from the singer and amount of nest cover.Signal-to-noise ratios are presented in dB.Given that we calculated signal-to-noise ratio by subtracting the noise amplitude from the signal amplitude (as in Halfwerk et al., 2012; Rehberg-Besler et al., 2016), signal-to-noise ratio values were occasionally negative; this occurred only when the signal was distant and the measured noise amplitude was effectively equal to the signal amplitude, resulting in a very small (<4.3) negative value.Lastly, we used a Kruskal-Wallis test to evaluate signal-to-noise ratio for different dominant plant species surrounding each nest.
To better understand the implications of our findings for how many tutors young birds can hear from within the nest, we georeferenced and digitized the 2021 and 2022 territory maps using QGIS.We calculated maximum territory diameter of each male using the following QGIS formula: “distance (centroid (minimal_circle ($geometry)), point_n(minimal_circle ($geometry), 1)) × 2”.To measure how many vocal tutors occur within the acoustic environment around each nest, we created a model in QGIS to count the number of neighbouring males who had at least 10% of their territory within a specified distance from each nest site (males with less than 10% of their territory falling inside a neighbour’s acoustic environment will likely be heard rarely, if at all).Based on the previous estimate that the acoustic environment of young Savannah Sparrows extends to 100 m from a young bird’s location (this distance is based on the estimate that songs transmit effectively up to 100 m in this species; Wheelwright et al., 2008), we used a buffer radius of 100 m to represent the traditionally defined acoustic environment.We used this distance, rather than the 117.7 m distance that we estimated for above-nest microphones in this study, to better compare our results to previous estimates of young Savannah Sparrows’ acoustic environment.To represent the acoustic environment from the perspective of inside the nest, under attenuating vegetation, we used a second buffer radius equal to the distance at which we estimated signal-to-noise ratio to equal zero from the results of this study: 78.6 m (see Results).In QGIS, the 100 m and 78.6 m buffers identified all male territories with at least 10% of their territory area within 100 m (for the traditional acoustic environment) or 78.6 m (for the nestling acoustic environment)of each nest site.We determined the number of male tutors within both the traditional and nestling acoustic environment of each nest site by counting the number of male territories overlapped by each buffer.We then compared the number of male tutors found within each buffer.The number of male tutors was normally distributed (Shapiro-Wilks test:p>0.05), so we used a pairedt-test to compare the number of male tutors found within each acoustic environment.We report the mean and standard deviation for each and present the effect size of this comparison using standardized mean difference (or Cohen’sd).
We completed all statistical tests in R 4.2.2 (R Core Team, 2022).We used the lm function in R for linear regressions.We interpret our analyses using language of evidence (following Muff et al., 2021), using terms of no or little evidence, weak evidence, moderate evidence, and strong evidence.In addition, we interpret the strength of our analyses with effect sizes (as in Cohen, 1988) using terms of small, medium, or large effect.
3.Results
3.1.Sound attenuation and microphone location
We found strong evidence for a large effect of microphone position on sound attenuation (Fig.2).Savannah Sparrow songs had a lower signal-to-noise ratio when they were recorded within the nest (8.4 ±9.4 dB) than above the nest (19.5 ± 9.7 dB; Figs.3 and 4; Wilcoxon:W=2335,p< 0.001,n= 68; effect size = 0.97).In a post hoc analysis evaluating each note type separately, we found strong evidence that this same pattern held true for each note type separately (“i” notes:W=2125,p<0.001,n=65; “ch” notes:W=731,p<0.001,n=38; “dash”notes:W=265,p<0.001,n=23; “is” notes:W=1646,p<0.001,n=57; Appendix Fig.S2).

Fig.2.Sound spectrograms depicting the song of a male Savannah Sparrow recorded simultaneously from above the nest (upper spectrogram) and within the nest (lower spectrogram).This song was sung by a male 27 m from the nest.
3.2.Sound attenuation and singer distance
Song attenuation increased as the distance between the singer and the nest increased (Fig.5), both for songs recorded within the nest(multivariate linear regression:adjR2=0.48,F1,66=62.7,p<0.001,n=68) and above the nest (adjR2=0.48,F1,66=62.3,p<0.001,n=68).We estimated the average distance at which sound becomes indistinguishable from background noise as thex-intercept of the regression line (i.e.,a signal-to-noise ratio of zero).We found that the distance at which male songs became indistinguishable from background noise in above-nest recordings was 117.7 m but only 78.6 m for within-nest recordings.Both regression lines shared similar slopes, revealing consistent negative relationships for above-nest and within-nest songs.By comparing thexintercepts of the two regression lines, we estimated that the loss in audibility between the above-nest and within-nest microphone was equivalent to listening to a bird from a position 39.1 m farther away.
To better understand how the sound attenuation imposed by the nest environment influences the number of song tutors that nest-bound birds may hear, we created two circular buffers around each nest in QGIS: one buffer with a radius of 100 m to represent the traditional size of the Savannah Sparrow acoustic environment (Wheelwright et al., 2008),and one buffer with a radius of 78.6 m to represent the acoustic environment from within the nest.We found strong evidence for a large effect of acoustic environment size and the number of male tutors.On average, we calculated 15.0 ±3.3 males were potential tutors given the previously estimated transmission distance of 100 m, but only 10.9 ±2.9 males were potential tutors given our inferred transmission distance of 78.6 m.This average reduction of 4.1 ± 2.1 males provides strong evidence that the nestling acoustic environment of Savannah Sparrows is about 27% smaller than previously assumed (t-test:t=8.9,n=21,p<0.001, effect size = 1.1).
3.3.Sound attenuation,nest concealment,and nest vegetation
We found no evidence that the signal-to-noise ratio of male song varied with nest concealment (the percentage of the nest that was visually obstructed by vegetation when viewing from immediately above the nest; linear regression:adjR2=-0.02,F1,34=0.29,p=0.59,n=36 songs recorded in 2022).The average nest concealment was 92.3%,ranging from 61 to 100% (n=10 nests).We found five dominant plant species immediately covering the 10 nests sampled in 2022: Lowbush Blueberry (Vaccinium angustifolium;n= 4 nests), Fireweed (Chamaenerion angustifolium;n= 1 nest), Northern Goldenrod (Solidago multiradiata;n=1 nest), grass spp.(n=2 nests), and Red Raspberry (Rubus idaeus;n= 2 nests).We found no evidence that the dominant plant species influenced signal-to-noise ratio (Kruskal-Wallis: Chi-squared =6.0,p=0.20,n=10).
4.Discussion
We simultaneously recorded live Savannah Sparrow songs from two perspectives, within the nest and directly above the nest, to determine whether song transmission is affected by the nest environment.We found strong evidence that songs recorded within the nest are more attenuated than those recorded above the nest: the signal-to-noise ratio was 11 dB lower for songs recorded within the nest, a major reduction in average sound amplitude.Based on our regression analyses, we estimated the maximum transmission distance for Savannah Sparrow song to be only 78.6 m from the perspective of nestlings within the nest.The nestling acoustic environment in Savannah Sparrows is smaller than previously estimated, reducing the number of male tutors by 27%.Overall, these results support our hypothesis that the nest environment attenuates song: the acoustic environment of nest-bound Savannah Sparrows is limited by the nest environment.
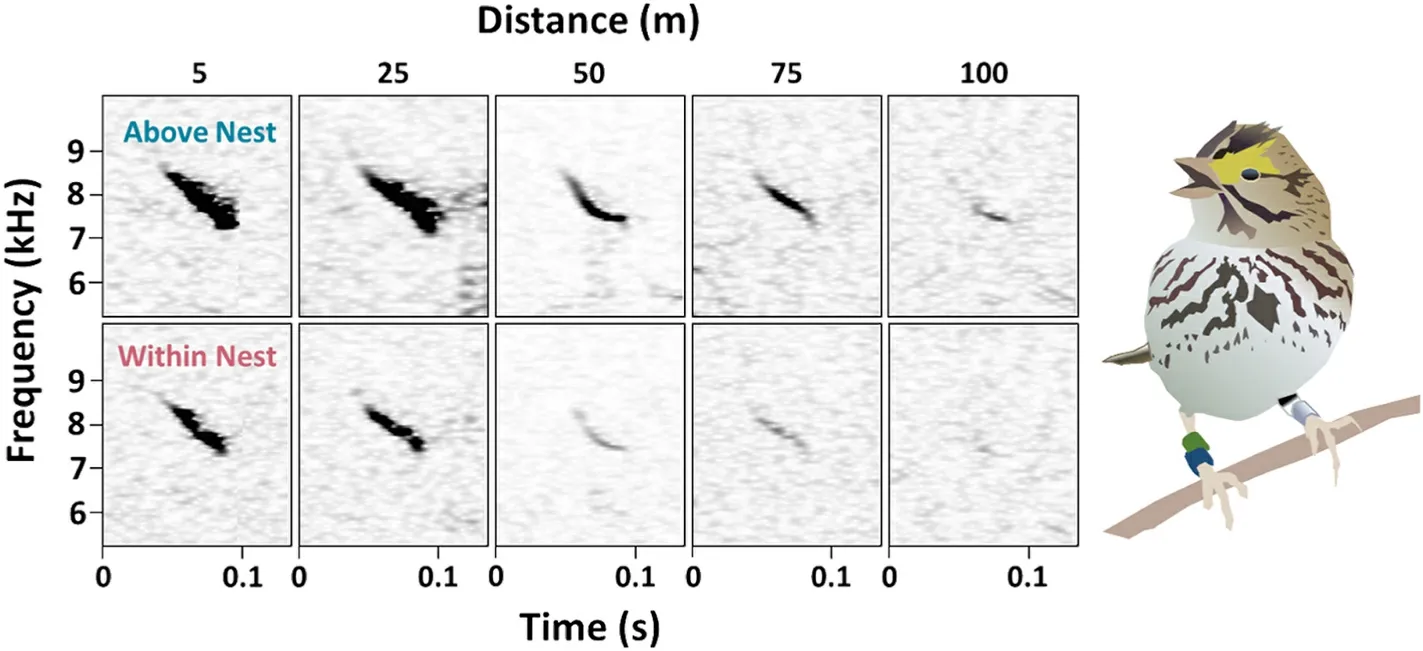
Fig.3.Sound spectrograms depicting “i” (introductory) notes from Savannah Sparrow songs recorded simultaneously from above the nest (top row) and within the nest (bottom row) for five different songs at five different distances from the nest.Sounds experienced more attenuation when recorded from within the nest (i.e.,bottom row vs top row), and as the distance between the singer and the microphones increased (i.e., moving from left to right).

Fig.4.Average signal-to-noise ratios for songs recorded above a Savannah Sparrow nest (blue) and the same songs recorded from within a nest (red).On average, notes recorded above the nest had signal-to-noise ratios 11 dB greater than the same notes recorded from within the nest.(For interpretation of the references to colour in this figure legend, the reader is referred to the Web version of this article.)
Sound naturally attenuates as it travels between a signaler and receiver due to spherical spreading (Wiley and Richards, 1978).Beyond spherical spreading, excess attenuation is caused by several factors,including weather, vegetation, or other physical barriers (Wiley and Richards, 1982).Savannah Sparrows build their nests on the ground,under cover of dense herbaceous vegetation (Wheelwright and Rising,2020).Foliage attenuates sound, and past research suggests that excess attenuation in closed habitats can equal 10 dB per 100 m (Wiley and Richards, 1982).Similarly, we found that songs recorded from within the nest, under dense vegetation, had a signal-to-noise ratio 11 dB lower than songs recorded above the nest, outside of the dense nest environment.In our analyses of vegetation density, we found no evidence that sound attenuation varies with the percentage of the nest concealed by vegetation, or the dominant plant species covering nests, although our sample size for these analyses was small (n =10 nests).
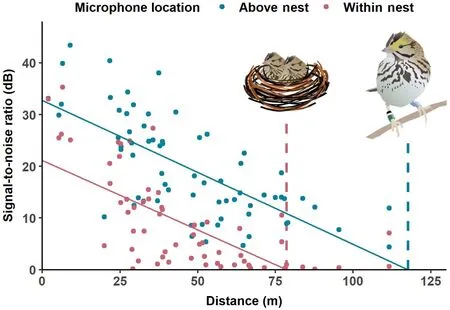
Fig.5.Average signal-to-noise ratio of Savannah Sparrow songs recorded from two microphones, one above a Savannah Sparrow nest (blue) and one within the nest (red), compared to the distance between the singer and the nest.The nest environment creates an acoustic handicap, in that songs recorded within the nest are more attenuated at all distances and become undetectable at an estimate of 78.6 m from the nest (i.e., the intercept of the red within-nest regression line on thex-axis).
Previous song transmission studies have found that the dense structure of nest cavities influences sound attenuation (Blumenrath et al.,2004; Lampe et al., 2004; Mennill and Ratcliffe, 2004; Grabarczyk and Gill, 2019).As in our study, prior investigations of cavity nests compared sound transmission within versus outside the nest environment, suggesting that the cavity exterior surrounding nests causes increased attenuation, reducing song transmission to the nest in the cavity interior.In Great Tits (Parus major), the cavity interior showed excess attenuation (attenuation in addition to that already produced by spherical spreading) of sound amplitude of 4 dB (Blumenrath et al.,2004).In the Pied Flycatcher (Ficedula hypoleuca), nest boxes accounted for 10 dB of excess attenuation in song (Lampe et al., 2004).In House Wrens (Troglodytes aedon) signal-to-noise ratio was ~10 dB lower inside the nest cavity than outside the nest cavity, however this was only found in territories with more anthropogenic noise (Grabarczyk and Gill,2019).In Black-capped Chickadees (Poecile atricapillus), a microphone mounted inside a nest cavity revealed that song amplitude decreased as the angle from the cavity hole increased (Mennill and Ratcliffe, 2004).While not directly comparing sound transmission within versus outside the nest environment, this study showed that the nest structure reduced sound amplitude by 6 dB; from -25 dB when the signal was produced in line with the nest hole, to -31 dB when the signal was produced opposite to the nest hole (Mennill and Ratcliffe, 2004).These prior investigations of cavity nests focused on the perspective of females listening from within the nest and explored the implications for intra-pair communication and extra-pair mating.Although it was not addressed by these previous studies, song attenuation by the nest environment may also have consequences for vocal learning nestlings.
Given the recent evidence that sounds heard during the embryonic and nestling periods may have important implications for song learning(Katsis et al., 2018; Kleindorfer et al., 2018; Schroeder and Podos,2023), it is important to consider how sound attenuation by the nest environment influences the acoustic environment of nestlings, to improve future research on early vocal learning.Many songbird nestlings are confined to their nests until they fledge.Previous studies show that young songbirds preferentially learn from the tutors with the least degraded songs (Morton et al., 1986; Peters et al., 2012), and tutors with songs that transmit most effectively through the environment (Hansen,1979; Moseley et al., 2018).Given the attenuating properties of nests that we have demonstrated here, only tutors close to the nest are likely to influence vocal learning at this stage.In the Kent Island Savannah Sparrow population, territories are small and closely positioned; in 2021 and 2022 the number of breeding males was 37 and 43, and the average male territory diameter was 57.6 m and 59.9 m (Sharma et al., personal observation).Previous research estimated that adult male song transmits up to 100 m (Wheelwright et al., 2008).Therefore, an average nest in this population will have 15 male territories within 100 m (including the social father’s territory).If we reduce our understanding of the acoustic environment of nestlings from 100 m to 78.6 m (the distance at which Savannah Sparrow songs appear to be indistinguishable above background noise in our dataset; Fig.5), the average number of tutors surrounding nests decreases by an average of 4 males, or by 27%.This not only alters the number of male tutors and songs that we expect nestlings to hear, but also alters the song diversity and the types of notes we expect nestlings to be exposed to.These findings have implications for vocal learning behaviour of any species where young are confined in protected environments at the onset of learning.The specific restriction imposed by the environment will vary depending on the structure of the natal environment, the habitat, the territory size, and the vocal behaviour of the adult song tutors.
We found no evidence that sound attenuation differs across nest concealment or dominant plant species.One explanation for this result is that the presence of any vegetation surrounding a nest is sufficient to attenuate sound, and small variation in the amount of vegetation, or the type of vegetation, does not have a large effect on sound transmission.In a broad-scale comparison of sound transmission through different nest materials, most nest substrates had similar effects on sound transmission(Potvin, 2019), providing evidence that the plant species around the nest may be unlikely to drive variation in sound transmission.Another possible reason is that Savannah Sparrows on Kent Island conceal nests under dense vegetation to protect offspring from predators (Weidinger,2002), specifically from American Crows (Corvus brachyrhynchos;Wheelwright et al., 1997).In this case, the cost of nest predation may outweigh the benefit of maximizing the nestling acoustic environment,and Savannah Sparrows may preferentially build nests under denser vegetation, despite the resulting acoustic handicap.Lastly,sound-attenuating effects within the nest may also be due to the placement of the nest on the ground.Signal reception can be improved by listening from higher perches (Dabelsteen et al., 1993; Holland et al.,1998; Mathevon et al., 2005), but nestlings cannot adjust their listening height and may be at a disadvantage when listening to tutors from their position in nests on the ground.
In addition to considering the attenuating properties of the nest structure, we encourage future studies on vocal learning to determine whether nests have a directional effect on sound transmission to nestlings.In cavity-nesting Black-capped Chickadees, the location of the singer relative to the nest hole orientation influenced song transmission into the nest; songs were less attenuated when they were produced toward the nest entrance (Mennill and Ratcliffe, 2004).Therefore, for cavity-nesting species, the nest orientation will skew the perception of females and nestlings residing in the nest.In Savannah Sparrows breeding on Kent Island, nest structure varies considerably, including nest cups with one main entrance that is covered by less vegetation, nest cups without any distinct entrance and all sides accessible, and in some rarer cases, nest cups with a tunnel-shaped entrance.We expect a tunnel entrance might have an effect similar to a cavity nest hole, possibly creating a skewed nestling acoustic environment, where tutors found in the same direction as the nest opening are most influential.The directional properties of sound attenuation by bird nests remains an interesting area for future investigation.
In conclusion, we found that the nest environment attenuates the songs of Savannah Sparrows, and therefore influences the acoustic environment of nestlings by limiting the distance at which they can hear adult tutors.The hearing range within a nest is much smaller than the hearing range just 1 m above the nest.Other species are expected to show a similar effect, although the extent of sound attenuation is likely to depend on several factors, including song features, habitat characteristics, and variation in nest materials and nest structure.Ongoing research that explores the importance of early exposure to conspecific song, and the effect of this exposure on vocal learning, will be important for quantifying whether the acoustic limitations of the nest influence vocal learning.We stress the importance of considering how nest properties influence song transmission across species, so that we can improve our understanding of vocal learning.
Funding
We thank the Natural Sciences and Engineering Research Council of Canada (NSERC) for financial support through grants to D.J.M., D.R.N.,and A.E.M.N.We thank the Animal Behaviour Society, the American Ornithological Society, and the Society of Canadian Ornithologists for research grants to S.L.D.
Ethics statement
This research was completed using permits from the University of Windsor (Animal Care Permit AUPP-20-09) and Canadian Wildlife Service banding permit #10766.We limited invasive measures by reducing handling time as much as possible during banding and by keeping a safe distance from nest sites.We deployed stereo recorders at nest sites only after the nestlings fledged from their nests, to further reduce disturbance to breeding adults and nestlings.
CRediT authorship contribution statement
Sarah L.Dobney:Conceptualization, Formal analysis, Funding acquisition, Investigation, Methodology, Writing – original draft,Writing – review & editing.Mikayla M.K.Bornais:Investigation,Methodology, Writing – review & editing.D.Ryan Norris:Investigation, Writing – review & editing, Funding acquisition.Amy E.M.
Newman:Funding acquisition.Heather Williams:Writing – review &editing.St´ephanie M.Doucet:Writing – review & editing, Investigation.Daniel J.Mennill:Conceptualization, Formal analysis, Funding acquisition, Investigation, Methodology, Resources, Supervision,Writing – original draft, Writing – review & editing.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgements
We thank Bowdoin Scientific Station for logistical support; this paper represents paper #292 from the Bowdoin Scientific Station.We thank S.Sharma for assistance with georeferencing Savannah Sparrow territories.We thank H.Wilson, L.Nong, and S.Mueller for assistance with nest searching and bird banding.
Appendix A.Supplementary data
Supplementary data to this article can be found online at https://doi.org/10.1016/j.avrs.2023.100148.
杂志排行

Avian Research的其它文章
- Selecting the best: Interspecific and age-related diet differences among sympatric steppe passerines
- Morphology and morphometry of two hybridizing buntings at their hybrid zone in northern Iran reveal intermediate and transgressive morphotypes
- Characteristics of cross transmission of gut fungal pathogens between wintering Hooded Cranes and sympatric Domestic Geese
- Fecal DNA metabarcoding reveals the dietary composition of wintering Red-crowned Cranes (Grus japonensis)
- Short-term night lighting disrupts lipid and glucose metabolism in Zebra Finches: Implication for urban stopover birds
- Variaiton in the composition of small molecule compounds in the egg yolks of Asian Short-toed Larks between early and late broods