Selecting the best: Interspecific and age-related diet differences among sympatric steppe passerines
2024-01-22JuliZurdoPulmezpezAdriBrreroDnielBustillodeRosJulimezCtsMrgritReverterCristinrezGrndosMnuelMorlesJunTrb
Juli Zurdo, Pul Gómez-López, Adrián Brrero,b, Dniel Bustillo-de l Ros,b,Juli Gómez-Ctsús,b, Mrgrit Reverter,b, Cristin Pérez-Grndos, Mnuel B.Morles,b, Jun Trb,b
aTerrestrial Ecology Group,Department of Ecology,Universidad Aut´onoma de Madrid(TEG-UAM),Madrid,Spain
bCentro de Investigaci´on en Biodiversidad y Cambio Global,Universidad Aut´onoma de Madrid(CIBC-UAM),Madrid,Spain
cEcology Department/IMEM“Ram´on Margalef”,Universidad de Alicante,Alicante,Spain
Keywords:18S Faecal material Nestling diet Prey choice Resource partitioning
ABSTRACTParental food provisioning is crucial for the growth and survival of offspring.Growth rate depends on food quality and food supplied to offspring may differ from what adults use for their own.In the case of steppe passerine birds, detailed characterization on nestling dietary composition, as well as prey choice and resource partitioning among species, is a pending subject.Dietary differences between nestlings and adults remain also largely unexplored.By using faecal DNA metabarcoding, we described the diet of nestlings and adults of five shrub-steppe passerine species over the 2017–2019 breeding seasons in central Spain.We also monitored arthropod availability in the field to assess dietary selection.We expected interspecific dietary differences to limit competition for food resources among sympatric species, as well as parental selection of high quality prey for nestlings.We also predicted age-related differences, with nestlings being fed nutrient-rich prey more frequently than adults.The main arthropod orders provisioned to nestlings were Orthoptera, Julida, Araneae and Lepidoptera.Nestlings of the different species showed high interspecific diet overlap, indicating both a coincidence in growth needs among bird species and no or little limitation of the most profitable resources during the breeding season.Adults of all species showed higher diet richness than nestlings, and age-related differences in prey composition were mainly driven by the selection of the most easily digestible, larger protein- and calcium-rich prey for nestlings, which may favour their rapid growth, and avoiding highly sclerotized and less nutritional prey such as ants.Our study sheds light on the basic ecology and conservation of these declining steppe birds,indicating that interspecific competition may not be a major factor during the breeding season.Given the current global decline of arthropods, further long-term research would be necessary, along with the implementation of effective conservation measures that ensure a sufficient availability of resources identified as key prey in the diet of steppe bird nestlings.
1.Introduction
Nutrition in the early stages of development is a key factor affecting offspring growth, morphology, cognitive ability, and survival (Searcy et al., 2004; Arnold et al., 2007; Reed et al., 2014).In altricial birds,parental provisioning of food strongly conditions the quality of offspring nutrition (Naef-Daenzer and Keller, 1999), since nestlings are unable to feed themselves, and require an abundant and constant flow of nutrients to fuel their growth and development (Martin, 1987; Brodmann and Reyer, 1999).In wild populations, the quality of early nutrition is determined by several interacting factors, such as the spatio-temporal distribution and quality of food in the habitat (Brodmann and Reyer,1999; Naef-Daenzer et al., 2000; Radford, 2008), brood size, or the status of competing siblings (Nilsson and Gårdmark, 2001; Blanco et al.,2006).
Several studies have shown that birds’ diet can vary in relation to age(Barrett et al., 2007; Radford, 2008; McLeay et al., 2009; Orłowski et al.,2014; Davies et al., 2022), since nutritional requirements change over time (Davies et al., 2022).In many bird species, adults forage differently when searching for profitable prey for offspring than when feeding themselves (McLeay et al., 2009; Alonso et al., 2012; Jedlicka et al.,2017), often providing nestlings with food of higher quality than that used for self-maintenance (D¨anhardt et al., 2011; Orłowski et al., 2014;Arco et al., 2022).A clear example is provided by nestlings of most herbivorous birds, which are usually fed with protein-rich invertebrates(Klasing, 1998; White, 2011), needed for a rapid growth (Starck and Ricklefs, 1998).When consuming arthropods and other invertebrates,nestlings also meet their requirements for essential macronutrients other than proteins (e.g., fat and carbohydrates), energy, minerals, and water(Brodmann and Reyer, 1999; BureŠ and Weidinger, 2003; Arnold et al.,2010; Reynolds and Perrins, 2010; Razeng and Watson, 2015).Dietary differences between adults and nestlings have been also observed in some insectivorous passerine species such as Bluethroats (Luscinia svecica; Orłowski et al., 2014), Western Bluebirds (Sialia mexicana; Jedlicka et al., 2017) and Eurasian Reed Warblers (Acrocephalus scirpaceus;Davies et al., 2022).However, studies about age-related differences comparing multiple species that coexist in the same habitat are scarce(see Herlugson, 1982), which may provide a better understanding of selected prey provisioned to nestlings in a community context and a more comprehensive picture of trophic interactions in the habitat.
Iberian shrub steppes are among the most unique and threatened habitats in Europe (Sainz Ollero, 2013).They are considered a key habitat for the conservation of steppe birds, given the threats they are facing and the evident decline of their populations (Santos and Suárez,2005; Gómez-Catasús et al., 2018; Traba and Pérez-Granados, 2022).In Iberian shrub-steppes, there is a rich community of passerine species,which are sympatric and share morphological and ecological characteristics, including similar dietary habits, breeding ecology or nesting sites (Suárez et al., 2009; Barrero et al., 2023a, 2023b; Zurdo et al.,2023).In these circumstances, niche theory predicts that species will partition space, time or resources in order to reduce interspecific competition thereby allowing coexistence (MacArthur and Levins, 1967;Chesson, 2000; Salsamendi et al., 2012).Indeed, interspecific food resource partitioning has been reported among Iberian shrub-steppe species, although an overall diet niche overlap in the community was observed, probably reflecting enough abundance of shared food resources (Zurdo et al., 2023).Likewise, partial niche segregation among these passerine species was found both for nesting site (Barrero et al.,2023a) and for habitat use (Barrero et al., 2023b).Similarly, prey provisioning to nestlings is expected to differ between sympatric species to limit competition (e.g., Trevelline et al., 2018; Stillman et al., 2022).However, the diet of shrub-steppe passerine nestlings is still poorly understood and limited to a few studies and species (but see Suárez,1987; Herranz et al., 1993; Poulsen et al., 1998; Donald et al., 2001;Ottens et al., 2014).Possible reasons behind the paucity of this type of studies are, on the one hand, the difficulty in finding the nests, which are located on the ground and totally or partially covered by small shrubs or herbaceous vegetation (Barrero et al., 2023a); and on the other hand,the invasive (e.g., neck ligature; Herranz et al., 1993), laborious and time-consuming (visual examination of faecal contents; Pompanon et al., 2012) techniques traditionally applied to determine the prey consumed by nestlings.Recent developments in molecular techniques such as DNA metabarcoding have greatly improved the identification of prey in faecal samples, allowing for higher taxonomic resolution within dietary studies and increased taxonomic accuracy (Taberlet et al., 2012,2018; Alberdi et al., 2019).These recent advances allow for more reliable comparisons of nestlings and adult diet and examining diet overlap and resource partitioning of sympatric species (Trevelline et al., 2018;Davies et al., 2022; Spence et al., 2022; Zurdo et al., 2023).
In this work, we used DNA metabarcoding based on faecal samples collected from nestlings and adults of five sympatric shrub-steppe passerines to achieve multiple objectives.First, we aimed to describe the diet composition of nestlings of the five target species (Dupont’s LarkChersophilus duponti, the Eurasian SkylarkAlauda arvensis, the Greater Short-toed LarkCalandrella brachydactyla, the Tawny PipitAnthus campestris, and the Western Black-eared WheatearOenanthe hispanica) and to investigate the degree of dietary niche partitioning.We expected that nestlings of different species would consume different prey in order to limit interspecific competition for prey resources and facilitate species coexistence.Second, we measured prey availability in the habitat to evaluate the diet preferences in parental provisioning.We hypothesized that parents of all species would select high-quality nutritional prey to feed nestlings.Third, we compared the diet of nestlings and adults of the five species to assess for diet differences in prey richness and taxonomic composition, as well as in nutritional composition.We predicted that nestlings and adults would show diet differences, and that nestlings would be fed on a narrower range of prey, but of higher nutritional quality than adults.This study may provide important insights into the foraging behaviour of adults during nest provisioning, and new knowledge on how passerines exploit and partition prey resources during the breeding season, a period of high energy demands for the entire community (Martin, 1987).Knowledge of diet requirements of nestlings may be essential for designing effective conservation measures to ensure that an abundant supply of key prey is available for the persistence of bird populations, especially for these species, for some of which demographic bottlenecks have been described due to low productivity (García-Antón and Traba, 2021).
2.Materials and methods
2.1.Study area and species
Fieldwork was conducted from April to June 2017–2019 in Soria province (Spain), at the ‘Altos de Barahona’ and ‘Páramo de Layna’Special Areas of Conservation (SAC) and Special Protection Areas (SPA)of the European Union’s Natura 2000 Network (ES4170148 and ES4170120, respectively; Fig.1).These areas are characterised by a plateau landscape (between 1100 and 1200 m a.s.l.), covered by continental basophilic shrublands ofGenista pumila,G.scorpius,Thymusspp.,Satureja intricataandLavandula latifolia, dry perennial grasslands,and mixed grassland-scrublands (Zurdo et al., 2021).Arable crops,ploughed fields and pine afforestations are interspersed in the shrub-steppe matrix.
We collected faecal samples from adults and nestlings of the five target species.Sampling size per specie is presented in Table 1.These are passerines of similar size that inhabit open landscapes with low vegetation cover, coexisting in the Iberian shrub-steppe during the breeding season (Barrero et al., 2023b).All species nest on the ground and lay one or more clutches per year (Perrins, 1987).They are all migratory,excepting Dupont’s Lark, the only resident one (Gómez-Catasús et al.,2016).Clutch size usually varies between three and five eggs, with an incubation time of 11–14 days (Barrero et al., 2023a).Nestlings leave the nest between eight days of age in Dupont’s Lark and 14 days in the Tawny Pipit (Barrero et al., 2023a).Adults are primarily insectivorous during the breeding season (Cramp, 1988; Zurdo et al., 2023), with beetles, grasshoppers and spiders being the most frequently consumed prey (Zurdo et al., 2023).However, dietary differences between adults and nestlings of these species are still unknown.
2.2.Faecal sample collection
Faecal samples from adults and nestlings were collected in eight localities (equivalent in all respects to each other) across the study area during the breeding seasons (April to June) of 2017–2019.Adult individuals were captured using spring-traps baited with Mealworms(Tenebrio molitor) with the aid of a species-specific acoustic record to attract them.All birds were ringed, thus avoiding duplication of samples from the same individual.When captured or handled for ringing, individuals defecated spontaneously, and faecal samples were collected and stored in individual 1.5 mL plastic tubes with 98% ethanol and kept at -20◦C until processing in the laboratory.For a similar methodology see Zurdo et al.(2023).
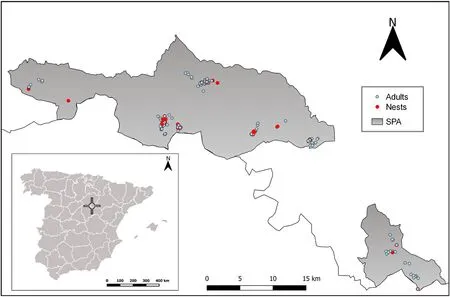
Fig.1.Location of study areas, where faecal samples of adults (blue dots) and nestlings (red dots) of shrub-steppe passerines were collected.Special Areas of Conservation (SAC) and Special Protection Areas (SPA) of the European Union’s Natura 2000 Network are depicted in grey (further north, ‘Altos de Barahona’, and further south, ‘Páramo de Layna’).The inset shows the location of the study area in Spain.(For interpretation of the references to colour in this figure legend, the reader is referred to the Web version of this article.)
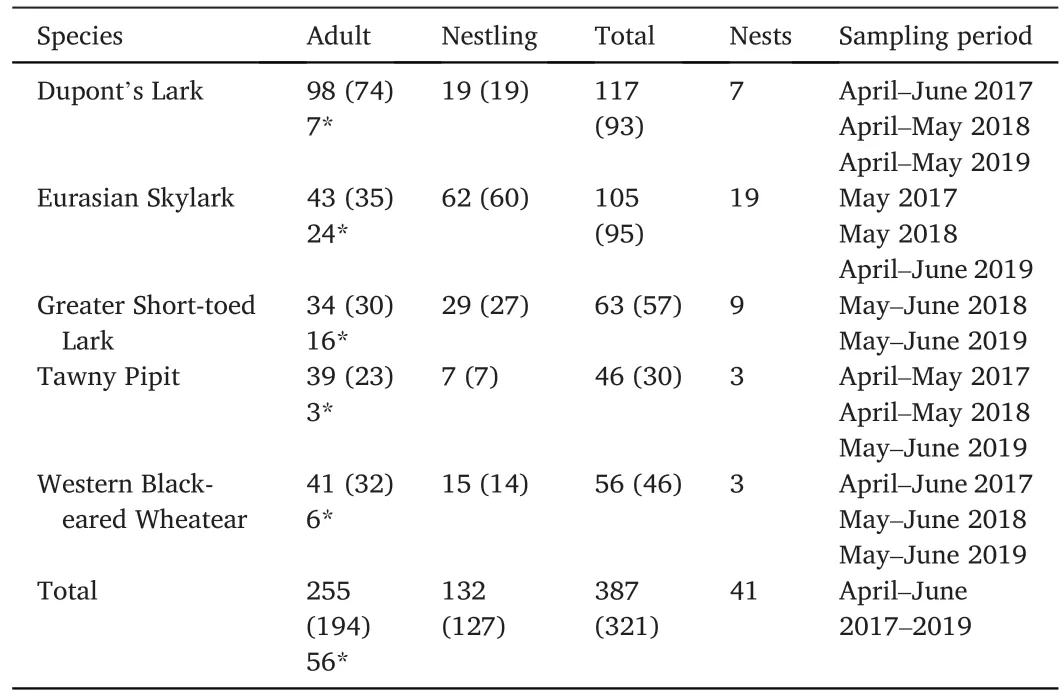
Table 1Number of collected samples per species and age, number of nests per species,and months in which samples were collected.The number of samples that produced dietary data is indicated between brackets.In the column Adult the number of samples from parents of a nest is denoted by an asterisk (*).
As regards nestlings, nests of the different species were first located,either by chance while performing other field tasks, or after an active search following the detection of an adult displaying breeding-related behaviour (carrying nests materials or food) (see Barrero et al.,2023a).The nests were monitored, always avoiding any disturbance,and once the nestlings were of a suitable size for handling (minimum four days old but generally with 7–9 days old, in the last stage of nestling development, which is relatively synchronous across species; Barrero et al., 2023a), they were ringed, and their faecal sacs collected.Faecal sacs were also stored in 1.5 mL tubes with 98% ethanol and opened by shaking to ensure mixing of faecal material and ethanol.Samples were frozen at -20◦C.Whenever possible parents of a nest were also trapped and their faecal material collected at the same date as their nestlings.
2.3.Arthropod sampling
To assess dietary selection of the target species we sampled arthropod abundance and biomass in 78 sampling stations at the same 8 study localities and sampling period of faecal samples collection(April–June of 2017–2019).In each sampling station, terrestrial grounddwelling arthropods were collected using three pitfall traps, placed at 5 m intervals.Pitfall traps consisted of a transparent plastic cup (230 mL,7 cm diameter, 10 cm depth), with holes in the upper part to facilitate rain drainage.Plastic cups were buried and protected by a PVC cylinder to prevent trap collapse and filled with 175 mL of 40% ethylene glycol and a drop of soap to reduce surface tension.After seven days of activity,traps were filtered, and arthropods were stored in plastic tubs with 70%ethanol.Flying arthropods were also sampled when collecting pitfall traps using an entomological sweep net along the longest distance between pitfall traps (10 ×2 m band).The flying trapped individuals were stored in the same plastic tube as ground-dwelling arthropods, and they were considered together.In the laboratory, we identified arthropods to order level, except for Hemiptera, which were discriminated according to the former division of Heteroptera and Homoptera, while Hymenoptera were differentiated between the family Formicidae and all the other Hymenoptera.We determined abundance of each arthropod group, and measured body length (excluding legs, antennas, and other appendices) of specimens using a digital calliper (±0.01 mm) to estimate overall biomass for each arthropod group applying the specific equations from Hódar (1996), which relate weight to body length in several arthropod groups of the Mediterranean region.Same procedure has been used previously in different studies (see, for instance, del Portillo et al.,2021; Gómez-Catasús et al., 2023).Abundance and biomass of arthropod groups per sampling station were calculated as the mean values of the pitfall traps that were active after seven days.
2.4.Molecular analysis
DNA was extracted from faecal samples using the QIAamp® PowerFecal DNA Kit (Qiagen), following the manufacturer’s instructions.Before extraction, ethanol was removed from the samples by decanting following 30 min of centrifugation and dried at 50◦C until the ethanol was vaporized.To analyse the diet of both adults and nestlings, we used two marker sets: a universal eukaryote 18S marker (miniB18S_81;Cabodevilla et al., 2023) and the arthropod marker ZBJ (Zeale et al.,2011) for COI region.Each marker was amplified in an independent PCR reaction, in which a negative PCR control (DNA-free) was included.Products were subjected to a second PCR to perform indexing and attach Illumina sequencing adaptors.PCR reactions were conducted following Cabodevilla et al.(2023) and Zeale et al.(2011) for each marker,respectively.PCR products were purified using AMPure XP beads(Beckman Coulter) and checked in Bioanalyzer before pooling per marker in equimolar amounts.The final two libraries were sequenced in an Illumina MiSeq NGS platform using a v3 MiSeq Reagent kit, following the manufacturer’s instructions.DNA extraction was performed by the Genomics and NGS Core Facility at the Centro de Biología Molecular Severo Ochoa (CBMSO, CSIC-UAM, Spain), and amplification, library preparation and sequencing by the Genomics Unit of the Fundación Parque Científico de Madrid (Spain).
2.5.Bioinformatic analysis
Bioinformatic processing of sequencing reads was done using MJOLNIR pipeline (Metabarcoding Joining Obitools and Linkage Networks In R; pipeline steps in Appendix S1), with a separate analysis for each molecular marker.We first used OBITools (Boyer et al., 2016) to quality filter and align paired-end Illumina sequences, and then we implemented VSEARCH (Rognes et al., 2016) to remove chimaeras.We clustered the sequences into molecular operational taxonomic units(MOTUs) using swarm (Mahé et al., 2015), which is based on an iterative aggregation of sequences that differ less than a given distance.For the taxonomic assignment, we first created a reference database in ecoPCR format for each molecular marker, obtained from the download of all 18S and COI sequences from NCBI database.We then used ecotag from OBITools to match the MOTUs generated to the reference sequences.We finally removed pseudogenes using LULU (Frøslev et al., 2017).MOTUs were identified with the most resolved taxonomic assignment possible,and those identified to species or genus were manually confirmed using the BLAST algorithm (NCBI), as well as their presence in Spain.We removed every taxa not belonging to Animal kingdom, as well as mammals (human), birds and internal parasites (phyla Nematoda and Platyhelminthes).We also excluded taxa not considered as potential prey items (mites, ticks, springtails; da Silva et al., 2019; Zurdo et al.,2023).We finally removed MOTUs representing less than 1% of the total number of dietary reads (Drake et al., 2021) to avoid incorporating false positives resulting from tag-jumping events, and samples with less than 100 dietary reads as they were considered to have failed (negative PCR controls and some samples of different bird species).
Finally, for each sample, the dietary information derived from the two molecular markers was combined using a python 3.0 script (da Silva et al., 2019).The script takes into account the differences in taxonomic resolution provided by the different markers, assuming that a dietary component obtained at a lower taxonomic resolution (e.g., order or family) by one of the markers is the same as a component of the same taxonomic group obtained at a higher resolution by the other marker (e.g., genus or species).For example, if in a given sample the 18S detected Orthoptera and the ZBJ detected a species belonging to the family Acrididae, it was assumed that the two markers detected the same taxon,so it was merged into the taxon with a higher taxonomic resolution (in this example, the acridid species).
2.6.Data analysis
All analyses were performed using R version 4.3.0 (R Core Team,2023).A significance level ofα= 0.05 was considered.Diet analyses were performed based on presence-absence data at order and family levels, which reduces biases affecting the quantitative performance of metabarcoding (Cuff et al., 2022).We also calculated the frequency of occurrence (FOO) of each prey taxon for each bird species or age class at family and order levels, defined as the number of faecal samples in which a family or order was detected divided by the total number of samples per bird species or age class.
Prior to further analysis, we tested whether there was a ‘nest effect’in nestling’s diet, i.e., a possible non-independence of data between nestmates due to shared parents.Following Stillman et al.(2022), we calculated the dissimilarity matrix at order level for all sample pairs within each bird species, using the Jaccard index implemented in the function vegdist of the R packagevegan(Oksanen et al., 2022).These matrices were decomposed to represent two dissimilarity vectors: beta diversity among nestmates and beta diversity among non-nestmates(Stillman et al., 2022); and then compared using Wilcoxon rank-sum tests with the function wilcox.test of the package stats (R Core Team,2023).Results showed no significant differences between diversity measures for any bird species (see Appendix S2, Table I–S2), indicating that similar amounts of dissimilarity were found among nestmates and among non-nestmates (Stillman et al., 2022).We therefore determined that no ‘nest effect’ was evident for any bird species, and that each nestling could be considered as an independent observation regardless of the nest it belonged to.Thus, we did not account for this effect in subsequent diet analyses.
Generalized linear mixed models (GLMMs) were carried out to test for differences in prey average richness at both the order and family levels between nestlings of the different bird species, and between adults and nestlings.For that, we used the glmer function of the package lme4(Bates et al., 2015) considering the number of orders or families identified as response variable, using a Poisson error distribution, and with the variable locality as a random factor to account for a possible geographical effect.We fitted two independent GLMMs incorporating the following predictor variables: (i) species for differences between nestling species (only nestling samples); and (ii) age, species, and the interaction between them for differences between adults and nestlings(all adult and nestling samples).The variables of all GLMMs were checked for significance using ANOVA tests with function Anova of the package car (Fox and Weisberg, 2019).
To explore whether dietary composition differed between nestlings of different species, and between adults and nestlings, we fitted multivariate generalized linear models (MGLMs) using the function manyglm within the package mvabund (Wang et al., 2022).This function fitted the presence-absence data of prey taxa at order and family levels to binomial family generalized linear models.Two independent models were fitted incorporating the following predictor variables: (i) species for nestling dietary composition comparison (only nestling samples);and (ii) age, species, and the interaction between them for dietary composition comparison between adults and nestlings (both adult and nestling samples).The function anova.manyglm was used to perform significance tests for each predictor variable within the model, with the pairwise.comp argument to perform pairwise comparisons.The p.uni =adjusted argument was finally implemented in order to applied univariate tests, which identify any significant relationships between predictor variables and specific prey items within the diet matrix.P-values were estimated using parametric bootstrap (Monte Carlo) resampling,recommended for presence-absence data (Wang et al., 2012), with 999 bootstrap iterations.Non-metric multidimensional scaling (NMDS)based on Jaccard distance of prey families was used to visualize the differences in diet between nestlings of the different species and between adults and nestlings using the function metaMDS of the package vegan (Oksanen et al., 2022).
To quantify overlap in observed nestling diets between species, we used the package EcoSimR (Gotelli et al., 2015) to calculate Pianka’s index of dietary overlap (Pianka, 1973) between pairs of nestlings of different species.This index quantifies the degree of similarity between two diets and ranges from 0 to 1, where 1 indicates complete overlap.In EcoSimR, a null model simulation, based on randomization of dietary data (here based on FOO) is generated and used in a statistical comparison to test whether the observed niche overlap differs from what would be expected by chance.We used the randomization algorithm 3(RA3), which is a permutation test recommended for niche overlap studies that reshuffles zero and non-zero values (Lawlor, 1980), and 9999 repetitions for the simulation.
To test for prey preferences during parental provisioning, we used null modelling with the package econullnetr (Vaughan et al., 2018).It uses a null-modelling approach where the observed frequency of occurrence of the different prey taxa in the diet (at the order level,except for Hemiptera and Hymenoptera; see arthropod sampling section) is compared to the frequency expected based on the null model (i.e., their availability recorded in the field).Using the function generate_null_net, we ran two null models based on the diet data for each individual of each nestling species (presence-absence data), indicating the month and the year in which the faecal sample was collected (i.e.,sampling period: April, May or June 2017, 2018 or 2019), and on the availability data of each arthropod taxa identified on all sampling stations combined by sampling period; one model used abundance data and the other used biomass data.Models were run for 9999 iterations.Prey taxa present in the diet with a frequency significantly higher than expected by the null model indicates preferred prey, while the opposite indicates avoided prey (Vaughan et al., 2018).Results were visualized using the functions plot_preferences of the same package.
Finally, we tested whether adults and nestlings differed in the nutritional composition of the diet, in terms of protein, fat, carbohydrates and calcium.To do this, we extracted from the literature (Robel et al., 1995; Brodmann and Reyer, 1999; Rumpold and Schlüter, 2013;Razeng and Watson, 2015; López et al., 2016; Aradis et al., 2019; Reeves et al., 2021) the nutritional values associated to the most frequently consumed arthropod orders (Appendix S3).Means were calculated to obtain an averaged nutritional value for each arthropod order (percentage of dry mass for protein, fat and carbohydrates; mg/g for calcium).Then, we assigned to each averaged value a score between 0 and 5 as follows: a score of 5 was assigned to the arthropod order with the highest value of each nutrient, and the scores for the remaining orders were relativized to that value (Appendix S3, Table I–S3).The resulting scores for each order replaced the presences in the presence-absence matrix at the order level for each adult and nestling sample as the total nutritional value for each sample.We then calculated the sum of the scores of each nutrient for each faecal sample and performed a multivariate analysis of variance (MANOVA) using the R’s base function manova and with nutrient variables as dependent variables, and age,species, and the interaction between them as predictor variables.To understand which nutrients mostly contributed to differences, we further performed univariate tests using the R’s base function aov.
3.Results
A total of 387 faecal samples of 5 shrub-steppe passerine species were collected, including 255 adult samples and 132 nestling samples from 41 nests (see samples per species and age in Table 1).From those,321 samples yielded dietary data, giving an overall success rate of 83%,which ranged from 65% for the Tawny Pipit to 90% for the Western Black-eared Wheatear and the Eurasian Skylark (Table 1).Sequences obtained from the ZBJ marker were clustered into 246 dietary MOTUs,while those from the 18S marker were clustered into 148 dietary MOTUs.When integrating information from both molecular markers,the initial 394 dietary MOTUs were reduced to 339 (Appendix S4),which indicates that less than 15% of dietary information obtained by the individual markers was redundant.With the bi-marker approach we identified in nestling and adult samples 121 prey species from 84 families and 15 orders of the classes Insecta, Arachnida and Diplopoda of the phylum Arthropoda.One MOTU was identified to a Gastropoda clade(Panpulmonata) from an adult sample.Seventy-eight families were detected from adult samples and 45 from nestlings.Faecal samples yielded a mean of 4.1 (±2.7 SD, range = 1–12) unique prey taxa.Lepidoptera was the most frequently detected order in adult samples,present in 64.9% of samples, while the most frequently consumed order by nestlings was Orthoptera (FOO: 87.4%) (Table 2).Other important orders were Coleoptera (FOO: 50.5%) and Orthoptera (36.6%) for adults, and Lepidoptera (42.5%) and Julida (33.1%) for nestlings(Table 2).The orders Mantodea and Pseudoscorpiones were only consumed by adults, whereas Thysanoptera was only detected in nestling samples.Noctuidae and Acrididae were the families most frequently detected in adult and nestling samples, respectively (Appendix S5).
3.1.Diet comparison between nestlings and dietary overlap
We found no difference in the number of orders (LR Chisq =2.93, df=4,P=0.569) or families (LR Chisq =6.35, df =4,P=0.174) detected per faecal sample between nestling species (Table 2; Appendix S5).However, we found differences, at the order level, in nestling’s diet composition between species (deviance =63.80, residual df =122,P=0.008).Pairwise comparison and univariate tests revealed that differences in prey composition between species were only due to differences in the frequency of Lepidoptera consumption between nestlings of the Eurasian Skylark and the Greater Short-toed Lark (Appendix S6,Table I–S6), being higher in the first one (55% vs 25.93% in the second species, Table 2).Similarly, differences in prey composition at the family level were found between species (deviance =201.86, residual df =122,P= 0.001).Pairwise comparison and univariate tests (Appendix S6,Table I–S6) indicated that differences were driven by a higher frequency of consumption of the family Julidae by nestlings of Greater Short-toed lark in comparison to Dupont’s Lark nestlings (55.56% vs 10.53%,Appendix S5), and also driven by differences in the occurrence of families Nymphalidae and Geometridae in the diet of Eurasian Skylark and Greater Short-toed Lark nestlings (Appendix S6, Table I–S6), being higher in the first species for both lepidopteran families (Appendix S5).
Pianka’s overlap index for nestlings as a whole was 0.87 (P<0.001)and close to 1 in all pairwise combinations, showing significantly greater dietary overlap than predicted by the null model (Table 3).The NMDS plot based on family level showed a high degree of similarity in consumption of prey families among shrub-steppe nestlings (Fig.2, stress =0.152,k= 2).
3.2.Prey choice in nestlings
Theeconullnetranalysis characterizing dietary preferences of nestlings of shrub-steppe species indicated that 33% of consumptions were stronger or weaker than expected from the null model, providing evidence of dietary selection (Fig.3; Appendix S7, Table I–S7).Orthopterans were consumed more than expected from their biomass by nestlings of all species, as well as Lepidoptera, except for Tawny Pipit nestlings.Julida were consumed less than expected by Dupont’s Lark nestlings, but more than expected by nestlings of the Eurasian Skylark,the Greater short-toed Lark and the Wester Black-eared Wheatear.Nestlings of all species, except Dupont’s Lark, showed significantly fewer detections of ants (Formicidae) than expected from their biomass in the habitat (no ants were detected in any nestling faecal sample,Appendix S7).Coleoptera were also consumed less than expected by nestlings of all species except Tawny Pipit.Nestlings of Dupont’s Lark and Greater Short-toed Lark consumed Heteroptera more than expected from their biomass in the foraging habitat.See Table II–S7 and Fig.I–S7 in Appendix S7 for results using prey abundance data.
3.3.Diet comparison between adults and nestlings
We found that only age was significant in predicting prey richness differences at order level (LR Chisq =4.22, df =1,P=0.039) and family level (LR Chisq =7.30, df =1,P=0.007), being both order and family richness higher in adult samples (Table 2; Appendix S5).However,species and the interaction term between age and species were notsignificant predictors of family and order richness differences, suggesting that differences in adult and nestling diet remained constant across species (test statistics available in Appendix S8, Table I–S8).

Table 3Pairwise Pianka’s index values among nestlings of the five shrub-steppe species based on the frequency of arthropod families consumed.All values indicate statistically significant niche overlap (i.e., greater than expected by chance based on comparison with 9999 null models; **P=0.01–0.001, ***P<0.001).
Multivariate analysis revealed that age (deviance =191.88, residual df =319,P=0.001), species (deviance =123.33, residual df =315,P=0.001), and the interaction between age and species (deviance =73.03,residual df = 311,P= 0.001) were all significant predictors of steppe passerines prey composition at order level.Coleoptera, Orthoptera,Lepidoptera, Hymenoptera, Hemiptera, Diptera and Julida significantly influenced diet differences between adults and nestlings (Appendix S6,Table II–S6).Nestling diet was associated with higher frequency of occurrence of Orthoptera (87.40% vs 36.60% in adults) and Julida(33.07% vs 19.59 in adults), while adult diet had higher incidence of Lepidoptera (64.95% vs 42.52% in nestlings), Coleoptera (50.52% vs 22.05 in nestlings), Diptera (27.32% vs 13.39 in nestlings), Hemiptera(24.23% vs 10.24% in nestlings), and Hymenoptera (15.98% vs 0.79%in nestlings) (Table 2).Hymenoptera and Orthoptera were the main drivers of the differences between species, and Julida was the unique prey taxa affecting the age-species interaction (Appendix S6,Table II–S6), due to its higher frequency of occurrence in the diet of Dupont’s Lark adults than in the diet of Dupont’s lark nestlings, in contrast to all other species (Table 2).

Fig.2.Non-metric multidimensional scaling (NMDS) plot of the prey families detected in faecal samples of shrub-steppe nestlings, calculated using the Jaccard index and visualized in two-dimensional space (k= 2, stress = 0.152).Lines represent the joint of individual nestling to the mean centroid (great dots)of its species.Proximity of individual nestlings to the name of an arthropod family indicates the likelihood of individuals to have consumed that family.
At the family level, also age (deviance =305.50, residual df =319,P=0.001), species (deviance =455.40, residual df =315,P=0.001), and the interaction between age and species (deviance =154.40, residual df=311,P=0.001) were significant predictors of the diet composition of birds.The occurrences of the prey families Acrididae, Curculionidae,Formicidae, Noctuidae, Pamphagidae and Tenebrionidae were significantly influenced by age class (Appendix S6, Table II–S6), with Acrididae and Pamphagidae being more frequently consumed by nestlings than by adults, whereas the other prey families were more prominent in adult diet compared to that of nestlings (Appendix S5).Prey differences among species were mainly due to the families Acrididae and Cydnidae,and Julidae was the driver of the age-species interaction effect(Appendix S6, Table II–S6).The NMDS plot based on family level indicated a degree of separation in consumption of prey families among adults and nestlings (Fig.4, stress =0.155,k=2), the latter segregated towards the families Acrididae and Pamphagidae.

Fig.3.Dietary preference plots for the nestlings of the five shrub-steppe passerines, showing the observed interaction frequencies (dots) and the predictions from the null model with 95% confidence limits (horizontal black lines).White dots denote arthropod prey groups consumed in proportion to their biomass in the field, orange dots indicate prey groups consumed more frequently than expected, and blue dots represent arthropods preyed less frequently than expected.(For interpretation of the references to colour in this figure legend, the reader is referred to the Web version of this article.)
We found significant differences in the nutritional composition of the diet between adults and nestlings (MANOVA: Pillai’s trace =0.30,F1,311= 32.78,P< 0.001), and these age-related differences varied across species (interaction term in MANOVA: Pillai’s trace = 0.09,F4,311=1.90,P= 0.017).However, there were no significant differences between species (MANOVA: Pillai’s trace =0.06,F4,311=1.16,P=0.29).Univariate tests showed that age was significant in predicting the nutritional composition of birds in terms of fat (F1,311= 19.13,P<0.001), and calcium (F1,311=7.05,P=0.008), as well as the interaction between age and species in terms of calcium (F4,311=4.82,P<0.001).Adults showed higher fat content in diet than nestlings, while the opposite occurred regarding calcium, except for Dupont’s Lark (Fig.5).No significant differences were found between adults and nestlings in terms of protein and carbohydrates (see Table II–S3 of Appendix S3 for complete MANOVA statistics).
4.Discussion
This study describes for the first time the diet of nestlings of five shrub-steppe passerine species using DNA metabarcoding, as well as the differences in diet richness and composition between nestlings and adults.In addition, a high degree of trophic niche overlap between nestlings of the different species has been found, which is a relevant contribution to the knowledge of the food ecology and interspecific relationships of sympatric species.In short, all the results of this study seem to converge on the same principle: nestlings are fed by receiving easily digestible, large, and nutritious prey (mainly Acrididae, Julidae,Lycosidae and Lepidoptera).
4.1.Nestling diet composition
In our study, parents of all the species fed their offspring with a high supply of orthopterans, especially from the family Acrididae, which were present in all the nestling samples analysed from the Greater Shorttoed Lark and the Tawny Pipit, and in more than 90% of the nestling samples from the Western Black-eared Wheatear.These orthopterans have already been reported as an important food source for insectivorous and omnivorous birds (Sullins et al., 2018; Hebda et al., 2019) and to be the bulk of nestling diet in some lark species (Cramp, 1988;Mainjargal et al., 2013).Furthermore, a previous study describing the diet of nestlings of two larks in the shrub steppes of southeastern Spain(the Thekla’s Lark,Galerida theklae, and the Mediterranean Short-toed Lark,Calandrella rufescens) also highlighted a great importance of these arthropods in their diets (Herranz et al., 1997).These findings suggest that Acrididae are of extreme importance for the development of nestlings of these species and, therefore, could be a good indicator of habitat quality to define key areas for the reproduction of shrub-steppe passerines.
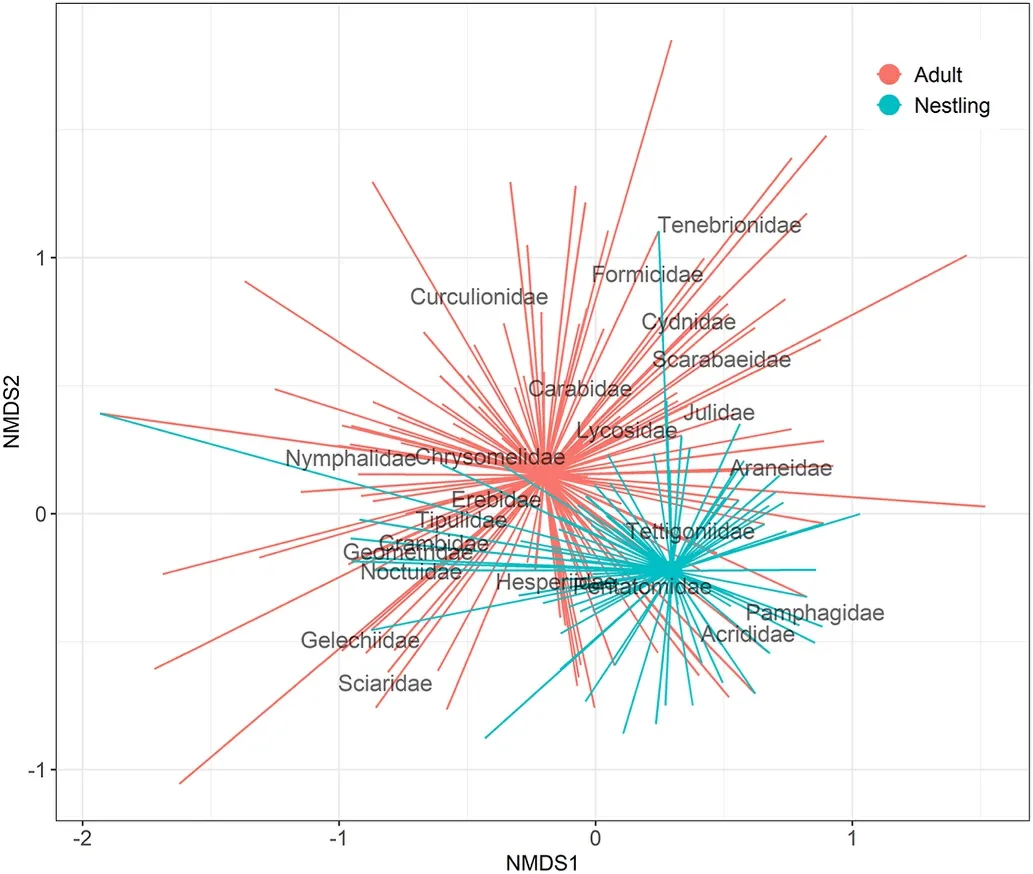
Fig.4.Non-metric multidimensional scaling (NMDS) plot of the prey families detected in faecal samples of shrub-steppe adults and nestlings, calculated using the Jaccard index and visualized in two-dimensional space (k= 2, stress =0.155).Lines represent the joint of individual bird to the mean centroid (great dots) of its age group.Proximity of individual nestlings or adults to the name of an arthropod family indicates the likelihood of individuals to have consumed that family.
Our results also showed that other important prey for nestlings of the study species were lepidopterans (mainly of the family Noctuidae),millipedes of the family Julidae, and wolf spiders (Lycosidae).Beetles were also consumed with high frequency by nestlings of most species,primarily dung beetles (Scarabaeidae), leaf beetles (Chrysomelidae) and ground beetles (Carabidae).Hemipterans of the family Cydnidae and dipterans of the family Tipulidae (probably in the larval stage) were also frequent prey in the diet of nestlings of Dupont’s Lark, the Eurasian Skylark and the Greater Short-toed Lark.For the latter species, to the best of our knowledge, this work provides the first data on the food ecology of nestlings.For Tawny Pipit nestlings, the only related information available was provided by Smetana and Gusewa (1981), who indicated that older nestlings received greater number of feeds than younger ones.In this study, only four arthropod orders and seven families were observed in their diet (Acrididae, Tettigonidae, Julidae, Curculionidae, Tenebrionidae, Geometridae and Nymphalidae).However,it should be noted that the number of samples from nestlings of this species was the lowest (n= 7), which may explain the lower total richness of prey consumed and urges for further sampling effort.On the other hand, our findings regarding the diet of nestlings of the Western Black-eared Wheatear are broadly consistent with those obtained by Suárez (1987) in Central Spain, who reported Lepidoptera, Orthoptera,Coleoptera and Diptera as their main prey.In addition, our results of the diet of Dupont’s Lark nestlings were in line with those described by Herranz et al.(1993) using cervical ligation, who also highlighted that nestlings were mainly fed on prey such as Acrididae, Lepidoptera,Lycosidae and Coleoptera.In respect to the diet of Eurasian Skylark nestlings, our results contrast with those obtained in other studies conducted in England (Poulsen et al., 1998; Donald et al., 2001) and The Netherlands (Ottens et al., 2014), which indicated a greater importance of the orders Coleoptera, Diptera and Hymenoptera.The different environmental contexts of those studies may partly explain their divergence with our results.No previous studies mention the remarkable consumption of millipedes by nestlings of all species obtained in our results.
4.2.Diet comparison between nestlings,dietary overlap and prey selection
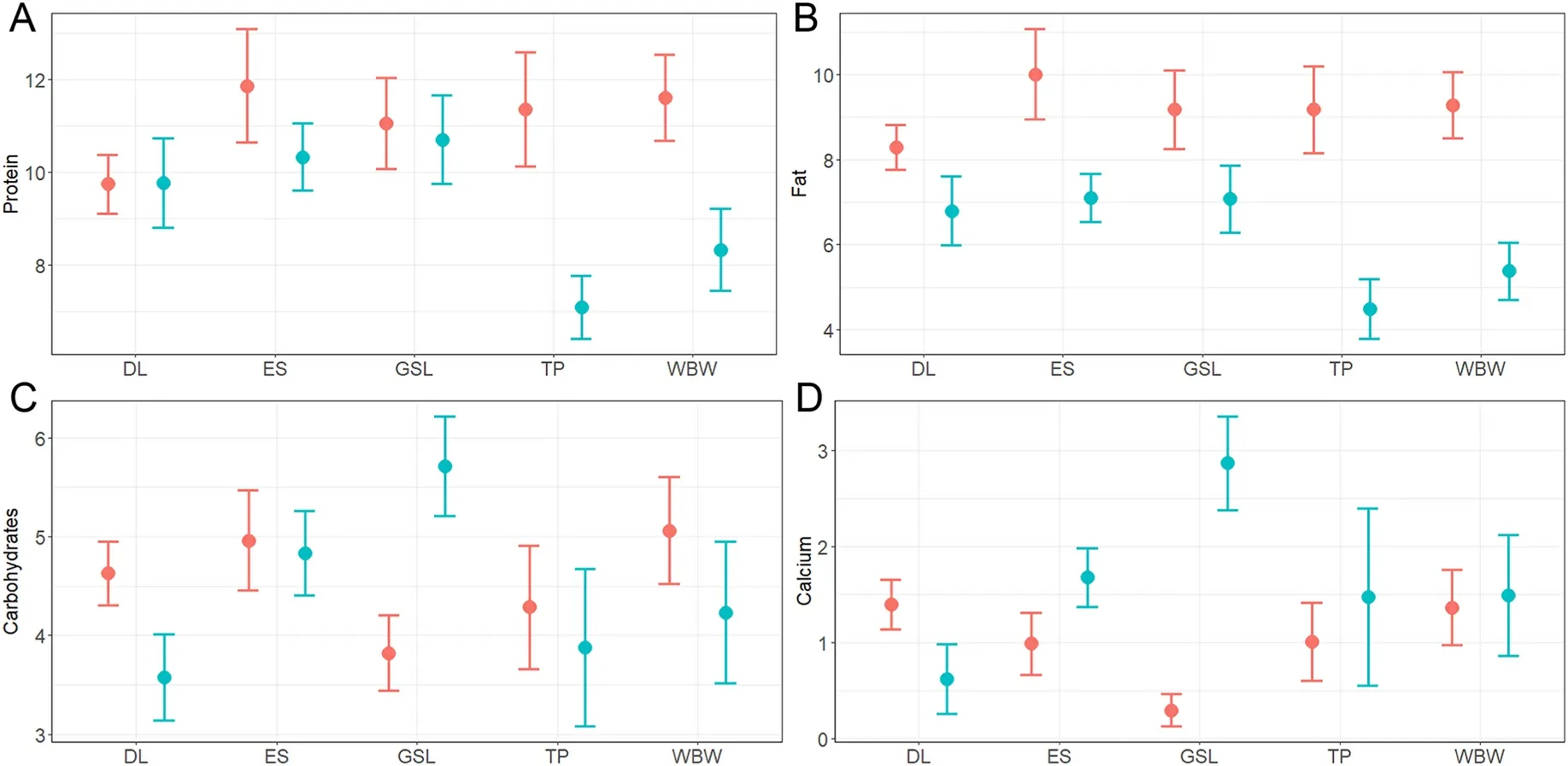
Fig.5.Nutritional values (mean ±standard error) of A) protein; B) fat; C) carbohydrates; and D) calcium of the diet of adults (red) and nestlings (blue) of five shrubsteppe passerines (DL: Dupont’s Lark; ES: Eurasian Skylark; GSL: Greater Short-toed Lark; TP: Tawny Pipit; WBW: Western Black-eared Wheatear).Significant differences were found between adults and nestlings in terms of fat and calcium.(For interpretation of the references to colour in this figure legend, the reader is referred to the Web version of this article.)
We expected that prey provisioning would differ between nestlings of the studied species in order to limit interspecific competition (e.g.,Trevelline et al., 2018).Despite we found differences in the diet composition of nestlings, these were subtle, and showed a high trophic niche overlap, for both the overall assemblage and for each pair of species.It is important to note that the few differences observed between species could be influenced by the moment of the breeding season when samples were collected, as nestling diet may vary due to seasonal changes in food availability, as has been observed in other passerine species (e.g., Davies et al., 2022).Also, the prey selection analysis indicated that parents of all bird species selected and avoided the same type of prey for their offspring, with some exceptions.For instance, prey such as Orthoptera, Lepidoptera and Julida were selected during parental provisioning for most species.These are groups of arthropods that provide an important source of crucial nutrients for nestling growth, particularly Orthoptera and Lepidoptera in terms of protein(Ueckert et al., 1972; Appendix S3, Table I–S3), and Julida in terms of calcium (BureŠ and Weidinger, 2003; Hames et al., 2006; Aradis et al.,2019) and carbohydrates (López et al., 2016).On the contrary, ants(Formicidae) were avoided during parental provisioning by most species, probably due to their small size and high chitin content, which make them a nutritionally less profitable prey (Razeng and Watson,2015; Reeves et al., 2021).Also, beetles were consumed by nestlings below their availability, probably due to the lower digestibility of adult beetles which usually contain high chitin proportions (Yang et al., 2013;Reeves et al., 2021).Beetle larvae, however, should be a more profitable prey for nestlings, but DNA metabarcoding does not allow to differentiate life stages, so it would be interesting to use additional techniques in future research (see below).Finally, the lower intake of Julida by Dupont’s lark in relation to all other species calls for further research.
The observed high dietary overlap, and hence, the resource sharing by nestlings of all species, although it could eventually lead to competition between them (Barrero et al., 2023b), suggests a non-limiting supply of resources for which they would theoretically compete(Soberón, 2007; Chase and Leibold, 2009), as the breeding season coincides with the peak of arthropod activity.This suggest that interspecific competition is not relevant in this context.However, the general decline of arthropod communities recorded worldwide (Hallmann et al.,2017; Cardoso et al., 2020; Wagner et al., 2021) and specifically in Spain(Galante et al., 2015), raises the alarm about possible resource limitation, which could eventually intensify interspecific competition (Schoener, 1982).This would certainly affect nestling survival (Grames et al.,2023), and thus impact on species’ viability (Bowler et al., 2019), as well as have consequences in the structure of the community assemblage(Chesson, 2000).Therefore, both the long-term monitoring of arthropod communities and the adoption of effective conservation measures for steppe habitats that guarantee an adequate availability and variety of food resources preferred by steppe bird nestlings should be priority actions.In Iberian steppes, crops have been shown to have negative effects on the availability of some arthropod groups and on the space use of steppe passerines, even beyond their limits (Reverter et al., 2021),suggesting that effective conservation strategies should be directed at improving the habitat quality (food availability and vegetation structure) within the steppe habitat patches.
4.3.Diet comparison between adults and nestlings
According to our predictions, we found important age-related diet differences both in terms of richness and composition.Adults of all species showed higher diet richness than nestlings, which also indicates that nestlings are selectively fed, receiving a smaller variety of arthropod groups (Orłowski et al., 2014).Adults consumed Coleoptera,Hymenoptera, Diptera, Lepidoptera and Hemiptera more frequently than nestlings, while nestlings were fed more frequently on Orthoptera and Julidae, prey groups of higher nutritional quality in terms of protein and calcium, respectively.Higher diet quality allows faster nestling growth (Costantini, 2010; Honarmand et al., 2010), which can accelerate fledging (Johnston, 1993).These shrub-steppe species nest on the ground, which makes them suffer high nest predation rates(Pérez-Granados et al., 2017; Gómez-Catasús et al., 2021), so that nestling growth needs to be fast (they remain in the nest for 8–14 days;Barrero et al., 2023a), and early fledging reduces the period during which nestlings are vulnerable to nest predation.
Despite the high consumption of protein-rich prey such as orthopterans and spiders (Appendix S3, Table I–S3) by nestlings, we found no differences between adults and nestlings in the protein composition of the diet, as adults also frequently incorporated these and other proteinrich groups such as Coleoptera and Diptera into their diet.Thus, our results suggest that adults provided their nestlings prey as rich in protein as those they used to feed themselves, but probably reserving in many occasions larger prey (grasshoppers and wolf spiders) for their offspring.Parents favouring larger prey for nestlings have been observed in other insect-eating passerines, such as Water Pipits (Anthus spinoletta; Brodmann and Reyer, 1999), Great and Blue Tits (Parus majorandCyanistes caeruleus; Naef-Daenzer and Keller, 1999), House Sparrows (Passer domesticus; Schwagmeyer and Mock, 2008) or Eurasian Reed Warblers(Davies et al., 2022).We also found no differences between the diet composition of adults and that of nestlings regarding carbohydrates,which are macronutrients also essential for adult birds (Barboza et al.,2008), and can be obtained from Lepidoptera, Hymenoptera or Orthoptera, in addition to millipedes (Rumpold and Schlüter, 2013).
Our findings indicated that nestlings consumed calcium-rich prey with higher frequency than adults, except in the case of Dupont’s Lark.Birds require increased calcium intake during nestling growth, as is involved in skeletal mineralisation (BureŠ and Weidinger, 2003; Heiss et al., 2009).Thus, parents need to search for calcium-rich items to provide their offspring, such as millipedes.In the case of Dupont’s Lark,calcium rich-prey were consumed with higher frequency by adults than by nestlings, which could be related to the breeding phenology of the species.This resident species is the first to breed in the study area (from mid-March; Pérez-Granados et al., 2017; Barrero et al., 2023a), when weather conditions are still severe.This may influence adults, and specifically females, to increase the consumption of calcium-rich prey to produce thicker shells to better isolate their clutches from temperature changes (Attard and Portugal, 2022).This would imply that Dupont’s Lark nestlings might have higher calcium content at hatching than those of other species, since this micronutrient can be resorbed from the shell into the egg interior during embryo formation (Orłowski et al., 2016).Thus, a high intake of calcium-rich prey in the nestling stage might not be required for their adequate growth.Further research would be necessary to better understand our findings.
Regarding fat consumption, adult diet had a significantly greater content of this nutrient than that of nestlings, due to the higher consumption of fat-rich prey, mainly Hemiptera, but also Coleoptera and Hymenoptera (Appendix S3, Table I–S3).Fat, together with protein, is a major macronutrient in avian reproduction (Reynolds and Perrins,2010), so it is reasonable that adults of these species consume with high frequency fat- and protein-rich prey during the breeding season, to provide them a store of nutrients to meet the demands of egg production and incubation (Hails and Turner, 1985).However, these fat-rich prey usually contain high chitin proportions (Reeves et al., 2021), making them more difficult to digest by nestlings.
4.4.Metabarcoding limitations
In this study, we used two different molecular markers and integrated the information provided by each, thereby probably overcoming the problems associated with primer bias and providing a more comprehensive picture of diet composition than with either marker used separately.For instance, the ZBJ failed to detect three entire prey orders that 18S detected (Blattodea, Phasamtodea and Thysanoptera), and probably overestimated the dietary importance of Lepidoptera and Diptera (da Silva et al., 2019; Tournayre et al., 2020), while 18S failed to detect Mantodea, and appeared less biased, but tended to provide a lower taxonomic resolution of prey items.Nevertheless, the continuous development of improved molecular markers for insectivorous diets (e.g., fwh, Vamos et al., 2017; ANML, Jusino et al., 2019) may decrease the need to combine multiple markers, also reducing costs.The combined use of metabarcoding with traditional techniques, such as visual examination of faecal contents, may be also relevant to identify unexpected biases and failures of molecular approaches, e.g., secondary detection or contamination (da Silva et al., 2019), or to differentiate between life stages of prey consumed (Bravo et al., 2017).In addition,quantification of dietary metabarcoding data to understand the contribution of each arthropod taxa to adult and nestling diets might have provided a valuable extra information on the structure of bird-prey interactions (Cuff et al., 2022).However, there is little consensus on the extent to which proportions of reads generated correspond to the original proportions of prey items (Deagle et al., 2013; Pi˜nol et al., 2018;Lamb et al., 2019), mostly due to differential degradation rates of tissues in the guts of consumers, differences of DNA density between consumed tissues, PCR primer bias and random sampling during sequencing(Murray et al., 2011; Leray and Knowlton, 2017).With the use of aggregated diet data within groups (bird species or age classes) to obtain frequencies of occurrence per group, we have partially overcome this lack of quantification (Cuff et al., 2022).Nevertheless, future work should assess the quantitative potential of DNA metabarcoding to draw more robust conclusions and broaden the picture of the food ecology of insectivorous passerines.
Ethics statement
This work was approved by the Local Ethical Committee for Animal Experiments of the Universidad Autónoma de Madrid (CEI80-1468-A229).
CRediT authorship contribution statement
Julia Zurdo:Writing – review & editing, Writing – original draft,Visualization, Methodology, Investigation, Formal analysis, Data curation, Conceptualization.Paula G´omez-L´opez:Writing – review &editing, Formal analysis, Data curation.Adri´an Barrero:Writing – review & editing, Investigation, Conceptualization.Daniel Bustillo-de la
Rosa:Writing – review & editing, Investigation, Conceptualization.
Julia G´omez-Catasús:Writing – review & editing, Investigation,Conceptualization.Margarita Reverter:Writing – review & editing,Investigation, Data curation, Conceptualization.Cristian P´erez-Gran
ados:Writing – review & editing, Investigation, Conceptualization.
Manuel B.Morales:Writing – review & editing, Supervision, Project administration, Methodology, Conceptualization.Juan Traba:Writing– review & editing, Supervision, Project administration, Methodology,Funding acquisition, Conceptualization.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgments
The authors wish to thank Israel Hervás, Miguel Mu˜noz, Ana Santos,Inmaculada Abril-Colón, Filipo Colla, álvaro Ortega and Adaia Cid for their help during field and/or lab work.We also thank Vanessa A.Mata and Luís P.da Silva for their inestimable help with bioinformatic analysis and to two anonymous reviewers for their constructive comments.This study was supported by the European Commission LIFE Ricotí(LIFE15-NAT-ES-000802) and LIFE Connect Ricotí (LIFE20-NAT-ES-000133) projects.This is a contribution to the Excellence Network Remedinal TE-CM (S2013/MAE2719).JG-C is funded by a Margarita Salas postdoctoral fellowship (CA4/RSUE/2022–00205) provided by the Spanish Ministry of Universities and Universidad Autónoma de Madrid (Spain).CPG acknowledges support from Ministerio de Educación y Formación Profesional through the Beatriz Galindo Fellowship(Beatriz Galindo-Convocatoria 2020) and JZ acknowledges support from Ministerio de Universidades through the predoctoral FPU fellowship program.
Appendix A.Supplementary data
Supplementary data to this article can be found online at https://doi.org/10.1016/j.avrs.2023.100151.
杂志排行

Avian Research的其它文章
- Morphology and morphometry of two hybridizing buntings at their hybrid zone in northern Iran reveal intermediate and transgressive morphotypes
- Quiet in the nest: The nest environment attenuates song in a grassland songbird
- Characteristics of cross transmission of gut fungal pathogens between wintering Hooded Cranes and sympatric Domestic Geese
- Fecal DNA metabarcoding reveals the dietary composition of wintering Red-crowned Cranes (Grus japonensis)
- Short-term night lighting disrupts lipid and glucose metabolism in Zebra Finches: Implication for urban stopover birds
- Variaiton in the composition of small molecule compounds in the egg yolks of Asian Short-toed Larks between early and late broods