盐胁迫对食用型向日葵现蕾期叶片光合性能与冠层结构的影响*
2024-01-20董睿潇王永庆王鑫博李玉义王希全张宏媛宋佳珅逄焕成
董睿潇,王永庆,王鑫博,李玉义,王希全,张宏媛,宋佳珅,于 茹,逄焕成,王 婧
(北方干旱半干旱耕地高效利用全国重点实验室/中国农业科学院农业资源与农业区划研究所 北京 100081)
我国盐渍土分布广泛,总面积约为1300 万hm2,且还在持续增加[1]。盐碱胁迫已经成为影响作物产量和品质的主要障碍因素之一。盐碱胁迫对作物的危害包括离子毒害、渗透胁迫、氧化损伤、高pH伤害、代谢紊乱等,会严重抑制作物生长发育,造成作物光合作用减弱[2-3],并最终影响作物产量。从光合生理生态和株型角度探讨作物对盐碱胁迫的响应特点,对于深入揭示盐碱地栽培作物的耐盐碱机理,降低作物盐害并加速其生态恢复具有重要意义。
食用向日葵(Helianthus annuusL.,以下简称“食葵”)抗旱、耐盐碱,强适应性[4],是黄河上中游地区盐渍土主要种植作物之一。现蕾期是决定食葵结实率和产量的关键生育期,极易受到盐胁迫的影响。前人研究认为,盐胁迫会导致食葵生理和生长特性受到不利影响,其中受影响最严重的是与产量关系密切的光合作用[5]。土壤盐分诱导的渗透胁迫会导致气孔快速关闭以保持水分,造成CO2利用率的降低和碳同化的减少[6]。随着盐分水平的增加和盐胁迫持续时间的延长,离子过量积累会导致过量的活性氧(ROS)产生,引起光合器官的氧化损伤,同时伴随光合电子传递受到抑制[7],最终造成食葵光合能力降低。曾文治[8]研究发现,土壤盐分对向日葵全生育期的净光合速率、蒸腾速率和气孔导度等光合参数有抑制作用,光合参数随盐分水平的增加而降低。凌云鹤[9]研究表明,盐胁迫下,向日葵的叶绿素含量明显下降,盐胁迫阻碍了叶绿素的合成。
冠层结构可决定截获的光合有效辐射能,是影响食葵光合速率和光能有效利用率的重要因素。提高不同冠层中叶面积指数(LAI)和叶角分布的协同效应可以有效提高食葵的光合作用[10]。盐胁迫对食葵的冠层结构参数有不利影响。马韬等[11]研究发现,盐胁迫下,营养生长期食葵的叶面积指数受到抑制,且盐分水平越高,抑制效应越显著。陈泽彬[12]研究表明,现蕾期食葵的叶面积和叶片数随盐胁迫程度的增加而降低,其中叶面积对盐胁迫的响应更敏感。
内蒙古河套灌区是食用向日葵主产区之一,同时其盐渍化耕地面积较大。深入研究不同程度盐胁迫下食葵的光合生理特性,有利于阐明食葵对盐碱环境适应的生态学机制。目前,关于河套灌区盐胁迫下食葵不同叶位叶片光合速率变化以及冠层结构的研究相对缺乏。本研究旨在揭示不同程度盐胁迫下食葵不同叶位叶片光合速率变化的原因及机理,进一步了解盐胁迫下食葵对环境的响应程度和耐盐程度,以期为盐渍化土壤资源合理高效利用提供理论依据。
1 材料与方法
1.1 试验设计
试验地点在内蒙古河套灌区五原县城南乡美丰村食葵种植田内。土壤质地为砂壤土,平均容重为1.46~1.54 g∙cm−3,现蕾期土壤平均水分含量为21.24%,田间整个生育期地下水埋深0.7~2.6 m。供试品种为美国杂交食葵品种‘LD5009’。
试验设置轻度盐胁迫(LS)、中度盐胁迫(MS)和重度盐胁迫(HS) 3 个盐分处理,每个处理重复3 次,共9 个处理小区,小区面积为 36 m2,随机区组排列。3 个处理现蕾期0~20 cm 土层土壤含盐量平均值分别为1.00 g∙kg−1、2.68 g∙kg−1和4.93 g∙kg−1(表1)。食葵于2019 年6 月2 日播种,9 月25 日收获。采用点播器人工点播,穴口覆砂,穴距40 cm,行距60 cm,种植密度约为44 000 株∙hm−2。生育期田间管理措施与当地农户一致。

表1 不同处理土壤的盐分含量Table 1 Salt content of soil in different treatments
1.2 测定内容和方法
本研究于食葵现蕾期(2019 年7 月23 日)在各小区采用五点取样法取0~20 cm 土层土壤样品,用四分法留取混合样,自然风干并除去根系、植物残体等,研磨过2 mm 筛,用电导率仪和pH 计分别测定土壤pH 和EC,再根据公式S=EC1:5×0.064×5×10/1000[13](EC1∶5表示利用土水比1∶ 5 浸提液测定的电导率),计算土壤含盐量。
采用美国LI-COR 公司生产的LI-6400 型便携式光合测定系统在食葵现蕾期内(2019 年7 月23 日),选择晴朗无风的天气于9:00—11:00 进行光合参数的测定。每个处理小区选取长势相对一致的植株5 棵,每株叶片由上向下依次编号测定,每片叶读取数据5 次,取平均结果。测定参数为: 净光合速率(Pn,μmol∙m−2∙s−1)、光合有效辐射(PAR,μmol∙m−2∙s−1)、叶片温度(TL,℃)、蒸腾速率(Tr,mmol∙m−2∙s−1)、细胞间隙CO2浓度(Ci,mol∙mol−1)和气孔导度(Gs,mol∙m−2∙s−1)。光合作用的气孔限制值(Ls)按Berry等[14]的公式计算。
叶绿素采用SPAD-502 型叶绿素仪测定。叶面积(LA)按长宽系数法测定。叶倾角(LI)通过直尺测量叶片自然状态下的水平长度和垂直长度,根据三角函数求得。
顶叶是植株冠层顶部出现的第一片完全功能叶。采用美国LI-COR 公司生产的LI-6400 型便携式光合测定系统于8:00、10:00、12:00、14:00、16:00、18:00 分6 次测定不同盐分水平下现蕾期食葵顶叶的光合日变化及当天各环境因子的变化。每个处理小区选取长势相对一致的植株5 棵,对顶叶进行挂牌标记用于测定,每片顶叶读取数据5 次,取平均结果。
1.3 数据处理
初始数据在Excel 2019 中进行处理和绘图,采用SPSS 23.0 对数据进行统计分析,利用单因素方差分析(ANOVA)进行显著性检验,采用Duncan 法进行多重比较(P<0.05)。
2 结果与分析
2.1 盐胁迫程度对不同叶位叶片光合特性的影响
2.1.1 对不同叶位叶片净光合速率(Pn)的影响
由图1 可知,现蕾期食葵单株叶片的Pn总体随叶位(从上往下,下同)呈先升后降的趋势,不同程度盐胁迫下,食葵现蕾期最大光合功能叶均在中上位。LS 处理峰值出现在从上往下第5 位叶片,且第4~16位叶片始终保持较高的光合速率;MS 处理峰值出现在第7 位叶片,第2~7 位叶片光合速率较高;HS 处理峰值出现在第6 片叶,第4~7 位叶片表现出高光合速率,其中MS 和HS 处理的最大净光合速率比LS 处理分别降低5.09%和38.69%。LS 处理除上部嫩叶外,成熟叶片的光合速率均高于MS 处理,但差异不显著,二者均极显著高于HS 处理(P<0.01)。这说明盐胁迫会降低食葵叶片的净光合速率,并影响高光合功能叶的数量,盐胁迫越重,食葵的高光合功能叶越少。其中,重度盐碱对食葵叶片光合能力的负面效应极显著高于轻中度盐碱,这可能与食葵的耐盐能力密切相关。

图1 不同盐分水平下食葵现蕾期不同叶位叶片的净光合速率Fig.1 Net photosynthetic rate of sunflower leaves at different positions at budding stage under different salinity levels
2.1.2 对不同叶位叶片叶绿素含量的影响
不同盐分处理下,现蕾期单株食葵上部叶片叶绿素含量低,中部叶片含量高,变化相对平稳(图2)。中部叶片是食葵进行光合作用、形成光合产量的重要部位。LS 处理中上位叶片叶绿素含量由上往下逐渐上升,最大值出现在第23 位叶片;MS 处理与LS处理类似,其叶绿素含量最大值出现在从上往下第17 位叶片;HS 处理上位和下位叶片叶绿素含量较低,中位叶片含量较高,最大值出现在从上往下第10位叶片。MS 与HS 处理的中位叶片叶绿素含量差异不显著,却均高于LS 处理,其中MS 和HS 处理的叶绿素最大值较LS 处理分别提高6.84%和11.11%。盐胁迫提高了食葵叶片的叶绿素含量,这可能是食葵受到盐胁迫而形成的应对策略。
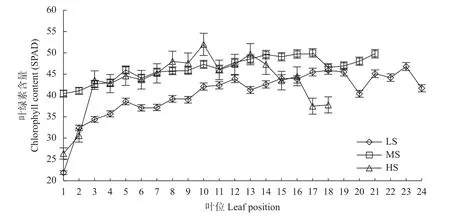
图2 不同盐分水平下食葵现蕾期不同叶位叶片的叶绿素含量Fig.2 Chlorophyll content of sunflower leaves at different positions at budding stage under different salinity levels
2.1.3 对不同叶位叶片光合参数的影响
由图3 可知,不同盐分处理下,现蕾期食葵单株叶片的气孔导度(Gs)随叶位的增加均呈先升后降的趋势。不同点在LS 处理上位叶较高,后波动下降,而MS 和HS 处理中位叶较高,后逐渐降低。不同盐分处理下,食葵不同叶位叶片胞间CO2浓度(Ci)均表现为中位叶相对稳定,上下位叶降低。不同叶位叶片蒸腾速率(Tr)的变化在各盐分处理之间存在差异,具体表现为: LS 处理上位叶较高,中下位叶逐渐降低,MS 处理上中位叶变化平缓,下位叶较低,HS 处理呈先升后降的趋势,中位叶较高。不同叶位叶片气孔限制值(Ls)变化为: 各处理均为上位叶和下位叶较高,中位叶较低。另外,由于相互遮阴,各处理均为上位叶光合有效辐射(PAR)总体较大,下位相邻叶片光合有效辐射差异极大,这也说明各处理相邻叶片各因子存在差异,应与叶片遮阴关系密切。
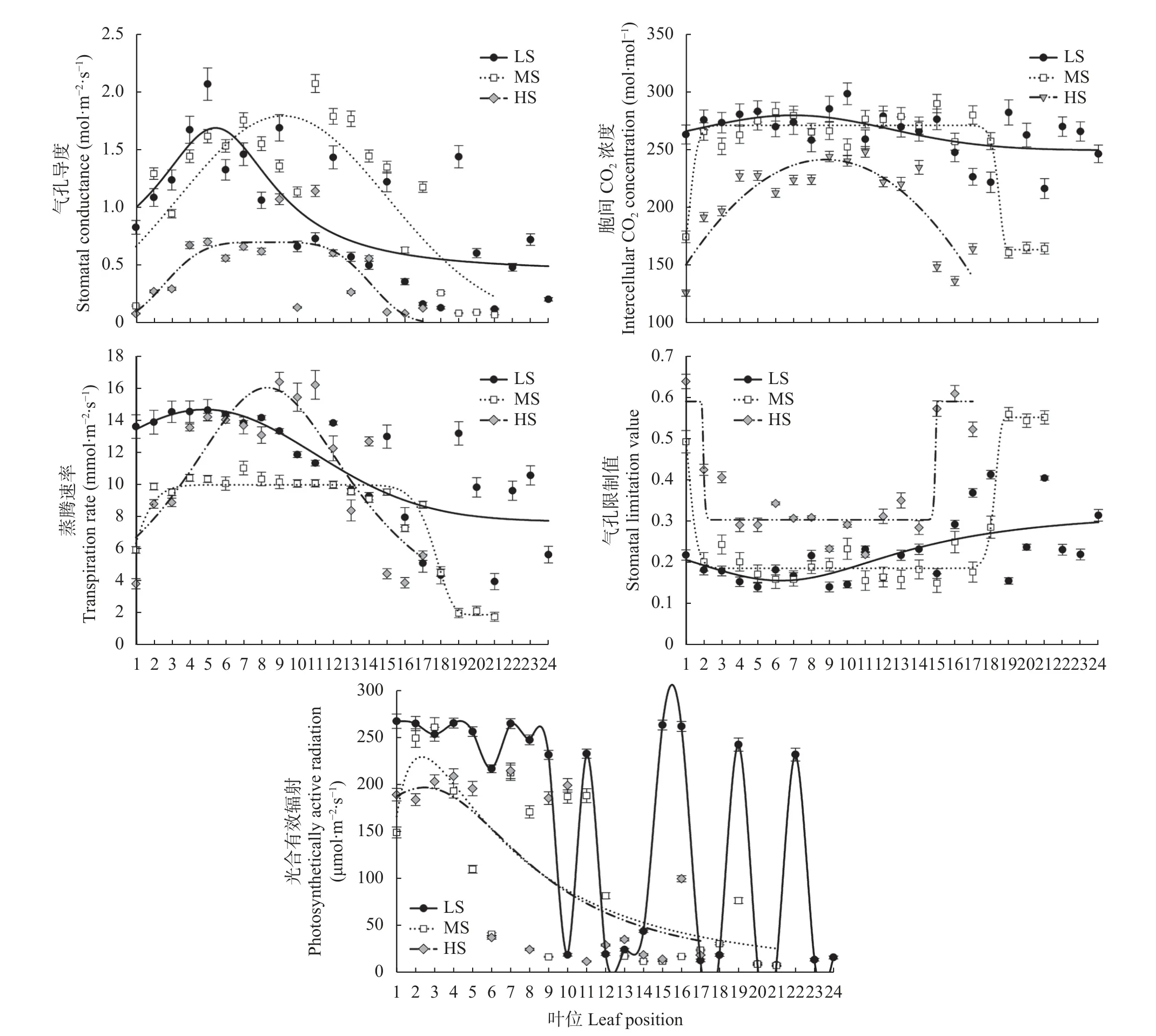
图3 不同盐分水平下食葵现蕾期不同叶位叶片光合速率影响因子变化Fig.3 Changes in the influencing factors of photosynthetic rate of sunflower leaves at different positions at budding stage under different salinity levels
2.1.4 不同叶位叶片净光合速率(Pn)与各因子的相关性分析
在不同盐分水平下,影响现蕾期食葵不同叶位叶片Pn的因素不同。由表2 可知,各盐分处理不同叶位叶片净光合速率与Gs、Ci、Tr、PAR、LA 呈正相关,与TL、Ls呈负相关,但其相关程度和相关系数并不相同。
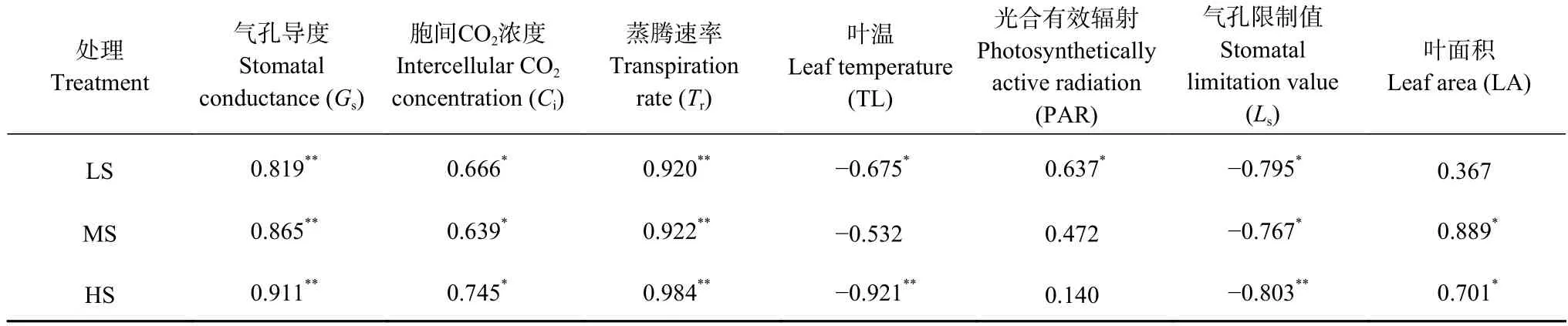
表2 不同盐分水平下食葵现蕾期叶片净光合速率与各因素之间的相关系数Table 2 Correlation coefficients between net photosynthetic rate and its affecting factors of sunflower leaves at budding stage under different salinity levels
LS 处理与Tr、Gs、Ls、TL、Ci、PAR 呈极显著或显著相关,其中与TL、Ls呈显著负相关,说明轻度盐胁迫下,不同叶位叶片光合速率主要受气孔因素与非气孔因素协同作用;MS 处理与Tr、LA、Gs、Ls、Ci呈极显著(P<0.01)或显著(P<0.05)相关,说明中度盐胁迫下,不同叶位叶片光合速率主要受非气孔因素作用;HS 处理与Tr、TL、Gs、Ls、LA、Ci呈极显著(P<0.01)或显著(P<0.05)相关,说明重度盐胁迫下,不同叶位叶片光合速率主要是非气孔因素主导。
从各因子与Pn的相关性可以看出,盐胁迫对食葵光合的影响具体表现在3 个方面: 1)盐胁迫造成食葵根部吸水困难,致使蒸腾速率下降,各叶片光合作用随之下降;2)还会造成食葵叶面积下降,尤其是中度和重度盐胁迫下,食葵光合叶面积减小,光合能力下降;3)盐胁迫越重,气孔因素在光合中的主导作用越低。
2.2 盐胁迫程度对顶叶光合特性的影响
2.2.1 不同盐胁迫下顶叶净光合速率(Pn)及各影响因子日变化
顶叶不存在遮蔽现象,能够充分接受太阳光的辐射,光合功能完全。另外,顶叶较为幼嫩,对盐胁迫的响应更为敏感,研究顶叶Pn的日变化有助于揭示不同程度盐胁迫下食葵的光合生理特性。图4 为不同盐分处理下现蕾期食葵顶叶净光合速率及各因子的日变化特征。由图4a 可知,LS 处理的Pn日变化为单峰曲线,主要呈先升后降的趋势;MS、HS 处理的Pn日变化为双峰曲线,说明中度、重度盐胁迫对食葵的光合能力产生较大影响,加重了食葵“光合午睡”现象。此外,盐胁迫也会推迟Pn最高值出现的时间,LS、MS 处理的Pn最高值出现在上午10:00,而HS 处理的Pn最高值出现在中午12:00。另外,随着盐胁迫的加重,多数时段食葵顶叶的Pn值显著降低,其中,与LS 相比,MS、HS 处理的食葵顶叶Pn最大值分别降低8.08%、14.66%。
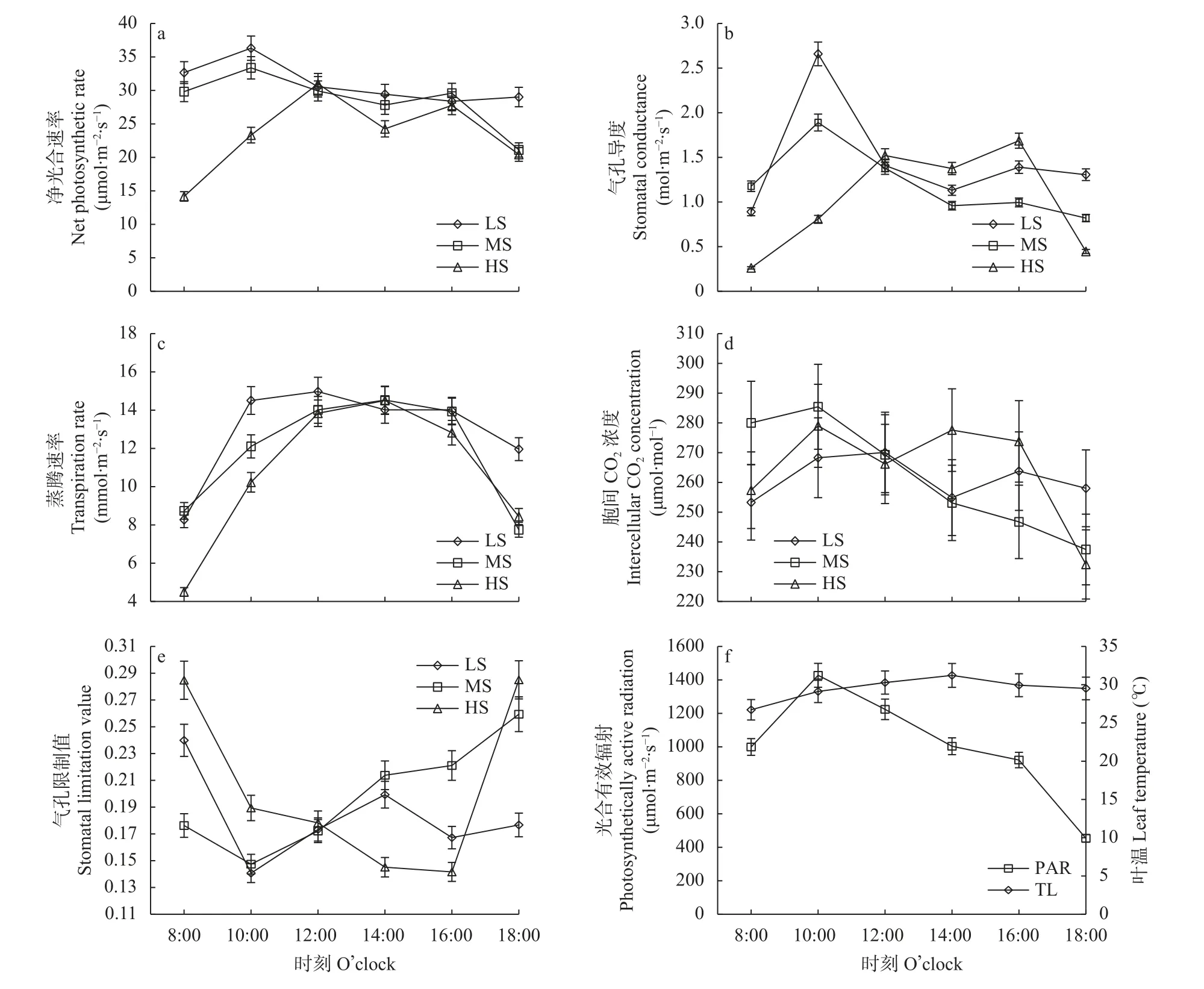
图4 不同盐分水平下食葵现蕾期顶叶净光合速率及各因子的日变化特征Fig.4 Daily changes of photosynthetic rate and its affecting factors of the top leaves of sunflower at budding stage under different salinity levels
气孔是水分、氧气和二氧化碳进出叶片的主要通道,控制叶片与大气之间的气体交换,对植物光合、呼吸和蒸腾作用都起到一定的调节作用[15]。气孔导度反映了气孔张开程度的大小。由图4b 可知,Gs日变化特征随盐胁迫程度的不同而不同。LS 处理的Gs与Pn变化趋势相似,在10:00 达到峰值,其后在14:00 降至谷底值,16:00 后又缓慢上升,这可能是由于12:00—14:00 时间段内的光照强度与外界温度较高,蒸腾速率加快,植株为防止水分散失过多而关闭气孔。MS 处理的Gs峰值也出现在10:00,但显著低于LS 处理(P<0.05),正午及之后逐渐降低。HS 处理的Gs值从上午开始逐渐增长,到中午时达到一定的高度,在16:00 达到峰值,之后急剧下降。从以上处理的日变化可知,不同盐分水平对气孔开度产生的影响不同,最终导致其对光合速率的影响不同。
由图4c 可知,不同盐分处理Tr日变化均为先递增后递减的单峰曲线,处理间差异较为显著(P<0.05)。LS 处理的Tr最大值出现在12:00,MS 和HS处理的Tr最大值出现在14:00。LS、HS 处理早上数值低于晚上,MS 处理早上数值高于晚上。除8:00和14:00 外,LS 处理的Tr值均高于MS 和HS 处理,除18:00 外,MS 处理的Tr值均高于HS 处理。这说明盐胁迫降低了食葵根系的吸水能力,盐胁迫加重使得不同时段叶片的蒸腾量逐渐减小。但正午时,由于温度较高,3 个处理之间差异不显著。
由图4d 可知,LS 处理的Ci日变化呈双峰曲线,在12:00 达最大值,在14:00 达最小值。MS 处理Ci的变化趋势为先升后降,在10:00 达最大值,在18:00达最小值。HS 处理Ci的变化趋势为先增后减,再增再减,在10:00 达最大值,在18:00 达最小值。MS 处理的Ci值在上午8:00—12:00 高于其他盐分处理,HS 处理在下午14:00—16:00 高于其他盐分处理。
图4e 中Ls的趋势是先降后升,LS、MS 处理均在上午10: 00 降至谷底,之后开始上升,LS 处理在14:00 后又开始下降,MS 处理则持续上升。HS 处理的Ls值在上午高于其他处理,其后缓慢下降,至16:00 达最小值,在14:00 至16:00 时段低于其他处理,随后迅速上升,18:00 又高于其他处理。
在图4f 中,PAR 和TL 呈先升后降的单峰曲线,PAR 在10:00 左右达最大值,随后逐渐下降。TL 在午后14:00 达最大值,随后缓慢下降。从Gs、Ci、Ls等因子的日变化对Pn的影响来看,在不同时段不同盐分处理下,Pn的主要限制因子不同。
由以上分析可以得出,各盐分处理Pn日变化的不同可能是由Pn对温度和光合有效辐射响应的不同造成的。
2.2.2 顶叶净光合速率(Pn)与各影响因子的相关性分析
不同盐分处理下影响现蕾期食葵顶叶净光合速率日变化的因素不同。基于SPSS 软件,利用皮尔逊(Pearson)简单相关系数分析各因素与Pn变化趋势。由表3 可知,不同盐分处理的顶叶Pn在多数时候与Gs、Tr、Ls呈正相关,多数时候与Ci、TL、PAR 呈负相关,但其相关程度及时间并不相同。此外,Pn与TL、PAR、Ls的相关性在不同盐分处理之间有较大差异,而且因盐分水平不同其相关性甚至相反。出现这种现象的原因是食葵受到盐胁迫而形成自发调节的适应性。
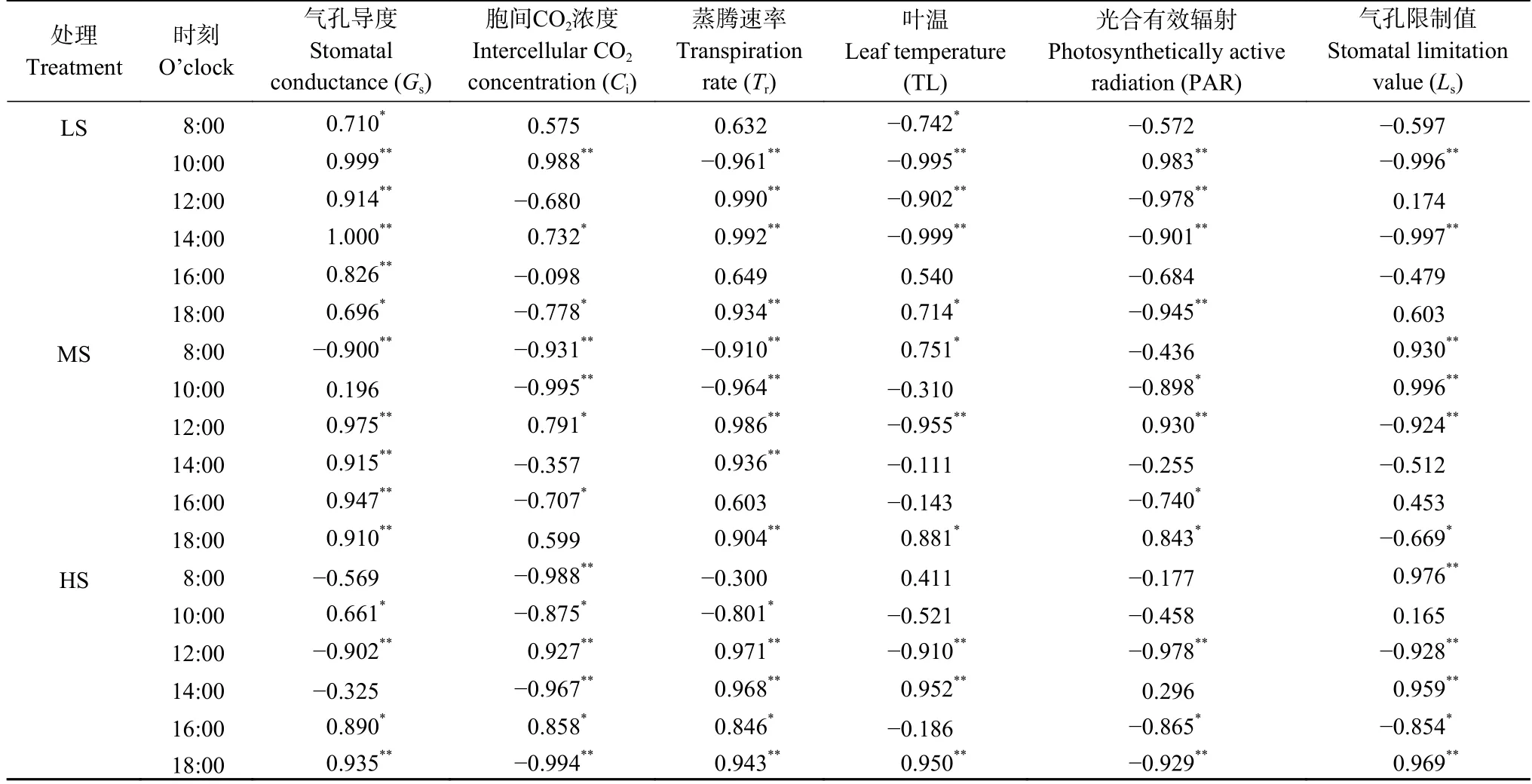
表3 不同盐分水平下食葵现蕾期顶叶净光合速率(Pn)与各因素相关系数比较Table 3 Comparison of correlation coefficients between net photosynthetic rate (Pn) and various factors in the top leaves of sunflower at budding stage under different salinity levels
另外,从各处理Pn与PAR 和TL 的相关关系来看,LS 处理的Pn仅在10:00 与PAR 呈正相关,在8:00—14:00 与TL 呈负相关,这说明LS 处理的Pn在强光和低温条件下会变高。MS 处理的Pn在12:00 和18:00 与PAR 呈正相关,在10:00—16:00 与TL 呈负相关,这说明MS 处理的Pn在强光和高温条件下降低。HS 处理几乎全天与PAR 呈负相关,在10:00—12:00 与TL 呈负相关,这说明HS 处理的Pn在高温和强光、弱光条件下都会降低。
进一步用相关系数来分析不同盐分处理下各时段食葵光合作用主要受到哪些因素的影响(表4)。结果表明,LS 处理下,各时段Pn变化气孔因素占主导地位,个别时段各因素协同作用;MS 处理下,14:00 时段,气孔因素占主导地位,其余时段气孔因素与非气孔因素协同作用;HS 处理下,气孔和非气孔因素协同作用。
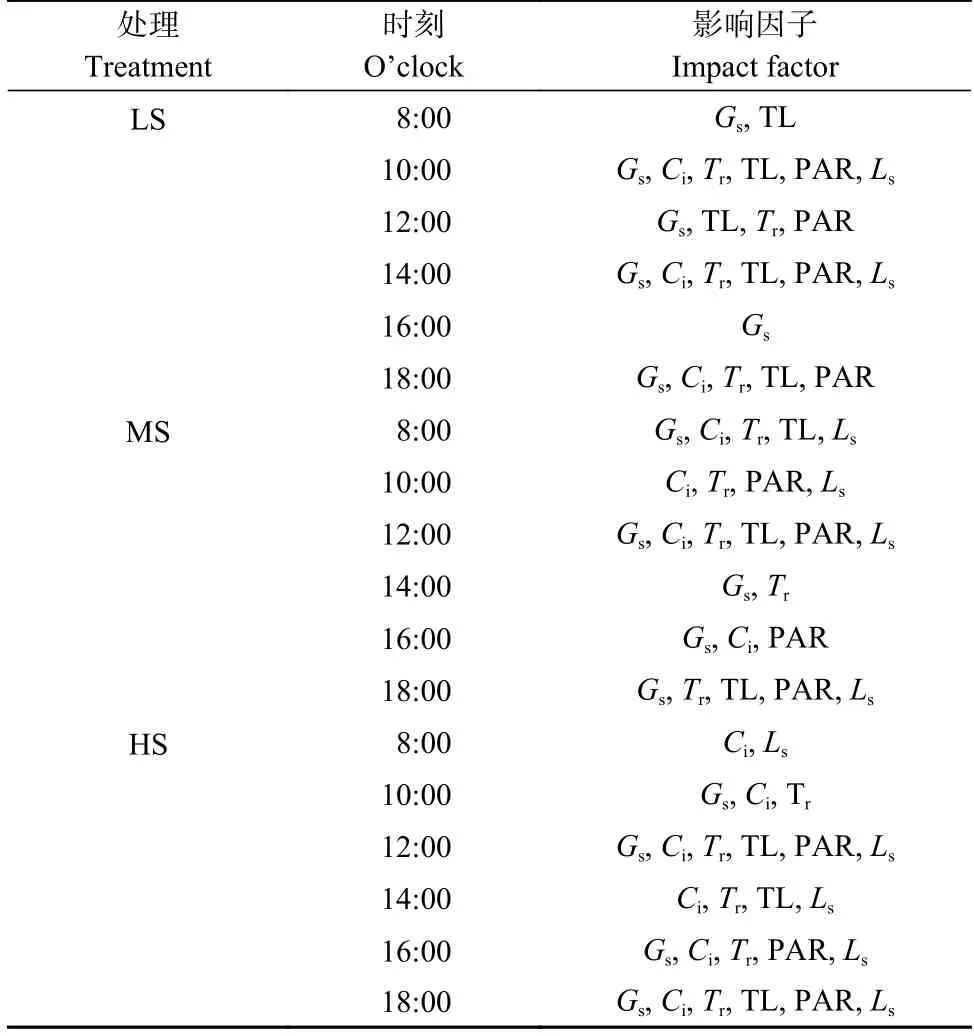
表4 食葵现蕾期不同盐分水平下不同时段光合作用的主要影响因子Table 4 The major impact factors of photosynthesis of sunflower at different periods under different salinity levels at budding stage
2.3 盐胁迫程度对株型冠层结构的影响
2.3.1 对不同叶位叶片叶面积的影响
由图5 可知,不同盐分处理下,现蕾期食葵单株不同叶位叶片的叶面积主要呈先升后降的趋势。LS处理峰值在第13 位叶,上位叶和下位叶面积均较小,中位叶面积较大,呈宝塔型冠层结构;MS 处理峰值在第6 位叶,除顶部嫩叶外,上部位叶面积较大,中下部叶面积明显减小,下部叶面积直线下降,呈平展型冠层结构;HS 处理峰值在第7 位叶,各叶之间差异较小,呈圆柱体型冠层结构。HS 处理的总叶面积较LS、MS 处理分别降低56.03%、47.74%。此外,盐胁迫降低了食葵单叶面积和主要功能叶面积。除上部叶片外,HS 处理的单叶面积均显著低于LS 和MS 处理,其中HS 处理的最大单叶面积较LS、MS处理分别降低38.71%、49.46%。由此可见,盐胁迫降低了食葵总叶面积和光合功能叶面积,使食葵冠层结构由伸展向收缩变化,导致食葵光合能力下降。
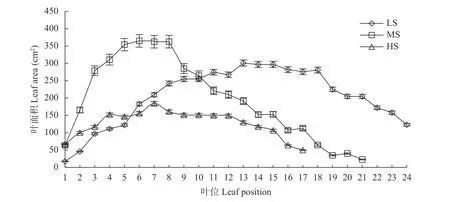
图5 不同盐分水平下食葵现蕾期不同叶位叶片的叶面积Fig.5 Leaf area of sunflower leaves at different positions at budding stage under different salinity levels
2.3.2 对不同叶位叶片叶倾角的影响
不同盐分处理下,现蕾期食葵单株不同叶位叶片叶倾角主要呈先降后升的趋势,上部叶片和下部叶片的叶倾角大,中部叶片叶倾角小(图6)。LS、MS、HS 处理都是从上往下第1 位叶的叶倾角最大,不同盐分处理之间存在差异,LS 处理第1 位与最后1 位叶片叶倾角差异不显著。MS 和HS 处理第1 位与最后1 位叶片叶倾角差异显著。平均叶簇倾斜角即叶片与水平面的夹角,值越大,叶片越直立,值越小,叶片越平伸。3 种处理下上位叶和下位叶的叶片较直立,而中位叶的叶片较为平伸。除第14 位叶片外,HS 处理的叶倾角均高于LS 处理,除第14~17 位叶片外,MS 处理的叶倾角均高于LS 处理。HS 处理的叶倾角最大值与LS、MS 处理相比分别提高30.92%、14.59%。由此可见,盐胁迫增大了食葵叶片的叶倾角,从而降低了对光的截获能力和光能利用效率。

图6 不同盐分水平下食葵现蕾期不同叶位叶片的叶倾角Fig.6 Leaf inclination angle of sunflower leaves at different positions at budding stage under different salinity levels
3 讨论
3.1 盐分对不同叶位叶片功能发挥的影响
光合作用是植物受盐胁迫影响的主要生理过程之一。其中,净光合速率是表征光合系统功能的直接指标[16]。试验结果表明,随着盐胁迫的加重,现蕾期食葵不同叶位叶片的净光合速率总体呈下降趋势,这与张俊莲等[17]研究结果相一致。本研究发现,不同盐分处理下食葵现蕾期叶片的净光合速率(Pn)随叶位均呈先升后降的趋势,最高值通常出现在中部叶片,上部和下部叶片Pn值较低。这与徐惠风等[18]的研究结果相一致,主要是因为上部为新生叶,发育不充实,光合功能未建成,下部叶片叶龄长,趋于衰老,光合能力有所下降。不同节位叶片发育的不平衡性造成Pn及其光合生理指标如蒸腾速率、气孔导度等均呈波动性单峰变化,在植物生长进程中具有普遍性。
目前,关于盐胁迫下食葵气孔和非气孔限制对光合作用影响的研究结果并不一致[19],这可能与盐胁迫的强度、持续时间和土壤基本性质等因素有关。本试验发现,盐胁迫下多数叶位叶片的胞间CO2浓度相对平稳,受气孔导度和气孔限制值变化的影响较小,这说明气孔限制不是影响食葵光合速率的唯一因素。另外,本试验还发现气孔与非气孔限制对光合作用的影响与盐胁迫水平有关,轻度盐胁迫下各叶位叶片的光合速率主要受气孔因素和非气孔因素的协同作用;中、重度盐胁迫下,主要受非气孔因素作用,这与郑国琦等[20]的研究结果类似,这可能是因为轻度盐胁迫下,低浓度盐分可能通过渗透胁迫或者刺激植物形成化学信号引起气孔导度降低而使CO2供应减少,进而导致净光合速率下降[21]。盐胁迫加重使气孔扩散的阻抗增加,核酮糖-1,5-二磷酸羧化酶(Rubisco)活性和底物1,5-二磷酸核酮糖(RuBP)再生能力降低,活性氧过度积累等破坏光合系统的结构和功能,非气孔限制占据主导地位,进而对光合速率产生不利影响[20]。
叶绿素是影响植物光合作用的关键因素之一。本研究发现,在盐胁迫加重的情况下,现蕾期食葵不同叶位叶片的叶绿素含量提高,这与Santos[22]研究结果相一致,可能是高盐胁迫诱导叶绿素降解酶的活性降低所致,但与Manivannan 等[23]、Foyer[24]研究结果不一致。这可能与食葵的耐盐性有关,通过提高叶绿素含量减轻盐胁迫对光合速率的不利影响,是一种保护策略,同时这也说明叶绿素并不是盐胁迫造成食葵光合速率下降的主要因素。此外,食葵在盐胁迫下,是否通过降低叶面积以增加叶绿素含量,还有待进一步论证。
3.2 盐分对顶叶光合功能的影响
顶叶是食葵冠层顶部第一片完全发育、功能健全的叶片,也是食葵主要光合功能叶。研究结果显示,盐胁迫使同一时段食葵顶叶净光合速率降低,这与孔东等[25]的研究结果一致。本研究发现,LS 处理的顶叶Pn日变化为单峰曲线,而MS、HS 处理的Pn日变化为双峰曲线,这说明中、重度盐胁迫下会加重食葵的“光合午睡”现象,但中、重度盐胁迫出现午睡现象的原因不同。试验结果显示,中度盐胁迫下,午间温度过高,空气湿度降低,蒸腾作用强烈,叶片出现缺水现象导致气孔部分关闭,造成CO2供给不足进而降低净光合速率[26];重度盐胁迫下,食葵的气孔导度在午间降低,但胞间CO2浓度却升高并出现峰值,可能是由于光合酶活性受到高温抑制进而限制光合功能[27]。
现蕾期食葵顶叶的蒸腾速率、气孔导度、胞间CO2浓度等的日变化总体上与光合速率相似,表现出盐胁迫下净光合速率主要受气孔限制影响的规律,这符合Farquhar 等[28]的气孔限制理论结果。试验结果表明,LS 在10:00—12:00 时段胞间CO2浓度并没有随气孔导度下降而降低,而是受到非气孔限制的影响;MS 在此时间段Pn下降是由于气孔限制,这可能由于LS 处理对光照和温度的敏感性更强烈,由于光合有效辐射增强和温度升高对光合系统的活性产生抑制作用,产生非气孔抑制现象[29]。
HS 在12:00—14:00 时段内的光合速率和气孔导度逐渐降低,而胞间CO2浓度却升高,应是高温导致叶片光合酶的活性受抑制,合成过程受阻,造成胞间CO2积累,导致非气孔限制并降低光合速率。而16:00—18:00 时段,光合有效辐射减弱,光合消耗CO2量减少,气孔开度随之降低,导致气孔限制。这一研究结果与韩瑞锋等[30]研究结果相似。总之,盐胁迫下气孔限制是影响不同时段叶片光合速率的主要因素,而非气孔限制是影响不同叶位叶片光合速率的主导因素。
3.3 盐分对株型结构的影响
冠层结构影响光在植株不同层次的分布情况、不同叶位叶片对光的截获能力以及相应的光合有效面积[31],直接影响光合作用。盐胁迫会影响根系吸水能力和叶片生长,最终改变植株冠层结构。叶面积是表征植物冠层结构最常用的参数之一,影响叶片对光能的捕获量和冠层光能利用效率。试验结果表明,盐胁迫会减小食葵总叶面积和主要功能叶面积,这与马韬等[11]研究结果一致,一方面可能是盐胁迫造成生理干旱,叶片缺水,在形态上表现为叶面积的下降;另一方面是盐胁迫减少了光合产物向叶的分配,影响叶片伸展速度,降低了光合叶面积[32]。此外,也可能是食葵的一种抗盐策略,通过减小叶面积来降低蒸腾,减少由质流引起的盐分离子积累,叶片形态变化是植物应对土壤盐胁迫的响应[5]。
研究发现,盐胁迫下食葵不同叶位叶片的叶面积均先升后降,这可能与植物适应盐胁迫并实现光合作用最大化表达有关,通过调节不同叶位叶片的生长速率,促使主要光合功能叶形成较大的叶面积,更有效利用光能[33]。
叶倾角也是表征食葵冠层结构的一个重要参数,是影响光截留和冠层光合作用的重要因素[34]。本研究发现,不同盐分水平下现蕾期食葵不同叶位的叶倾角分布呈现出一致的变化规律,上部和下部叶倾角较大,中部叶倾角较小,这与徐昭[35]发现的盐渍土食葵现蕾期上挺下垂的冠层结构一致。上部叶片叶倾角大,叶片较直立,利于更多光线到达中下部冠层[31],这种变化使得冠层内的光分布更加均匀,提高光拦截能力[36]。中部叶片叶倾角小,叶片较平展,对光的截获能力较强,从而有效提高食葵对光的利用率和光合能力。此外,研究发现,重度盐胁迫导致食葵叶片倾角增大,这可能是由于盐胁迫加剧了食葵的光抑制,食葵为减少潜在损害而调整叶片角度[37]。
研究还发现,LS 处理现蕾期食葵呈宝塔型冠层结构,MS 处理呈平展型冠层结构,HS 处理呈圆柱体型冠层结构,盐胁迫使食葵冠层结构由伸展向收缩变化,光合功能叶面积减小,轻度、中度盐胁迫下食葵叶倾角数值相对较小,其冠层结构更为合理,光能截获效率相对较高,群体光合势相对较大,叶片光合能力相对较强[38]。
4 结论
1)盐胁迫降低了现蕾期食葵各叶位叶片的净光合速率,其中中度、重度盐胁迫处理的净光合速率最大值较轻度盐胁迫分别降低5.09%、38.69%;同时,盐胁迫也减少了高净光合速率单叶的数量,即减少了高光合功能叶片。轻度盐胁迫处理下,不同叶位叶片的光合速率主要受气孔因素和非气孔因素协同作用;中度和重度盐胁迫处理下,主要受非气孔因素影响。随着盐胁迫程度的加重,影响光合作用的因素由气孔限制逐步转向非气孔限制,但叶绿素不是导致光合速率下降的主要因素。
2)盐胁迫降低了现蕾期食葵顶叶的净光合速率。中度、重度盐胁迫处理的食葵顶叶Pn最大值较轻度盐胁迫分别降低8.08%、14.66%。盐胁迫加重了食葵的“光合午睡”现象,光合能力受到抑制。轻度盐胁迫处理下,各时段顶叶Pn变化主要受气孔因素主导;中度、重度盐胁迫处理下,主要受气孔因素与非气孔因素协同作用。
3)轻度盐胁迫处理食葵呈宝塔型冠层结构,中度盐胁迫处理呈平展型冠层结构,重度盐胁迫处理呈圆柱体型冠层结构,盐胁迫使食葵冠层结构由伸展向收缩变化。盐胁迫会减小食葵总叶面积和主要功能叶面积,其中重度盐胁迫处理的总叶面积较轻度、中度盐胁迫处理分别降低56.03%、47.74%,重度盐胁迫处理的最大单叶面积较轻度、中度盐胁迫处理分别降低38.71%、49.46%;同时重度盐胁迫处理的叶倾角最大值与轻度、中度盐胁迫处理相比分别提高30.92%、14.59%。
