不同造林密度和冠层下杉木幼林叶片光合特性的差异研究
2024-01-20全小强王立冬李小玉胥清利梁海燕王燕茹马祥庆闫小莉
车 豪,全小强,王立冬,李小玉,胥清利,梁海燕,王燕茹,马祥庆,闫小莉*
(1.福建农林大学林学院,福建 福州, 350002;2.福建顺昌埔上国有林场,福建 南平, 353200)
杉木(Cunninghamia lanceolata(Lamb.)Hook.)是我国重要的速生用材树种,长期以来国内外针对杉木开展了大量的研究,初步形成了杉木人工栽培的技术体系,显著地提高了杉木人工林产量。但随着我国木材需求的加大,特别是大径级无节良材的供应无法保证我国木材产业的需求。虽然杉木育种水平提高,高世代良种大量应用,但传统的栽培技术已不能适应这些良种培育大径无节良材的要求[1]。研究表明,杉木人工林培育中不同造林密度对林分的生产力和木材品质具有显著的影响[2-3]。因此,研究和确定适宜的造林密度是有效提升杉木人工林生产力和木材品质关键培育措施,对培育大径无节良材具有重要意义。
杉木大径级无节良材的培育中,通过合理的造林密度和营林措施进行抚育,营建良好的林分结构和有效冠层,可促进林木生长速率和改善树木干形,最终提高杉木人工林产量和质量。研究表明,光合作用为植物生长发育提供物质和能量,树冠中的活枝净光合产物是主干生长的主要来源,但并非所有活枝都有助于主干的生长,部分位于主干下部的活枝其合成的光合产物只能满足自身的生长和呼吸,并没有额外的光合产物提供给主干生长[4]。如当枝叶净光合速率出现负值说明此时光照强度较弱,植株的呼吸速率大于光合速率,该层的枝叶会消耗植株生长所需的能量,应疏除该层枝叶[5]。冠层和枝条部位作为林分中关键的结构性因子,对于林木光合特性具有重要的影响。目前,国内外已有油橄榄(Olea europaeaL.)[6]、欧美杨(Populus×canadensisMoench)[7]、香椿(Cedrela sinensis(A.Juss.)Roem.)[8]、格木(Erythrophleum fordiiOliv.)[9]、栲树(Castanopsis fargesiiFranch.)[10]等多个树种开展不同冠层的光合能力研究,均发现树冠上层的净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr) 和水分利用效率(WUE) 高于树冠下层。因此,在林业集约经营中密度调控和一定程度的修枝是人工林培育中的重要措施。开展不同冠层和枝条部位的林木叶片光合特性的差异研究,可深入了解冠层结构和光照分布对林木光合能力的影响,并有助于优化林分的管理策略。
但是,杉木幼林在不同造林密度和不同冠层下的光合特性差异研究鲜有报道,特别是对杉木幼林的有效冠的高度未进行科学判定。故开展不同密度和修枝强度对杉木人工林冠层特征的影响研究,探讨有利于改善林分冠层特征的最佳经营技术组合,合理制定杉木人工林的培育措施具有重要的现实意义[11]。鉴于此,本研究在野外大田试验条件下,在5 个不同造林密度的杉木幼龄林中通过搭建6 m高的光合指标测定固定塔,通过测定不同冠层、不同方向和不同枝条部位叶片的光合特性指标,比较和分析不同造林密度和冠层下杉木幼林叶片光合特性的差异,对5 年生未修枝的杉木幼林的有效冠的高度进行科学判定,明确不同造林密度下杉木幼林合理的修枝高度,从而为杉木人工林的高效和优质培育提供科学依据。
1 材料与方法
1.1 试验地概况
试验地位于福建省南平市埔上林场(117°47′5″ E、26°56′8″ N)。属亚热带海洋性季风气候;海拔200~400 m,坡度20°~30°,年平均气温18.5 ℃,年平均降雨量1 880 mm,年平均日照时数1 699 h,无霜期260 d 以上,土壤以红壤为主,土层深厚,理化性能良好。
1.2 实验材料
选择5 年生杉木幼林及5 种造林密度,每个密度设置3 个20 m × 20 m 的重复样地,即每个密度造林面积为1 200 m2。于2022 年6 月和8 月在每个样地中心位置选择3 株长势良好且均一的标准木,在3 株标准木之间搭建测定不同冠层光合特性的固定梯架,共计5 个高度为6 m 的固定检测塔。
1.3 研究方法
如表1 所示,于每个造林密度样地选择3 株标准木,测定样树树高和胸径。以1 m 为一层,用喷漆进行画圈分层,分为5 个不同冠层(距离树体基部1、2、3、4、5 m 处)、枝条不同朝向(东西和南北)、枝条叶片不同部位(尖端和末梢)。在实际测定中发现距树体基部1 m 处树冠的枝条已腐烂、枯死无法测量叶片的光合值,故不同冠层从距离树体基部2 m 处开始测定和研究。

表1 不同造林密度下杉木幼龄林生长和土壤养分基本数据表Table 1 Basic data table of growth and soil nutrients of C.lanceolata under different planting densities
于2022 年8 月20—25 日上午9:00—12:00 的晴朗无云天,温度为31.3~40.1 °C、相对湿度为29.65%~63.46%、VPD 为1.31~5.58 k·Pa 的环境,采用LI-6400XT 便携式光合仪对5 个密度林的样树进行光合特征指标测定,包括净光合速率(Pn)、蒸腾速率(Tr)和气孔导度(Gs),并计算水分利用效率(WUE)。测定时将光量子通量密度统一设置为1 000 μmol·m-2·s-1,叶室温度设置在30 °C,CO2浓度设置在400 μmol·m-2·s-1,湿度控制在50%~75%[12]。同时用纸板画出与光合仪叶室大小的方框(10 cm × 10 cm),将纸板置于测定的部位底部进行拍照,每个测定部位采集8~10 片叶子,将采集的叶片带回实验室进行扫描、分析、计算叶面积和换算实际的光合特征值。
1.4 数据统计与分析
数据通过Microsoft Excel 2022 进行整理,通过SPSS24.0 软件进行数据分析,采用单因素ANOVA 检验分别对不同造林密度、不同冠层、不同方向、枝条不同方位4 个因子在不同处理间的差异进行分析,采用多重比较4 个因子间的差异,以P<0.05 作为显著性判断标准,利用Origin2022绘图。
2 结果与分析
2.1 不同造林密度和冠层下杉木叶片净光合速率的差异分析
由图1 可知,杉木东西、南北向尖端叶片和末梢叶片的净光合速率Pn值在5 种造林密度下均表现为距离树体基部5 m 处 > 4 m 处> 3 m 处 >2 m处。在5 种造林密度下,冠层5 m 处的Pn值大多显著高出冠层4、3、2 m 处(P< 0.05)(图1a~d)。其中,在M5密度下冠层2 和3 m 处的Pn值出现负值(图1a~d),在M4密度下冠层2 m处的Pn值出现负值(图1b、d)。
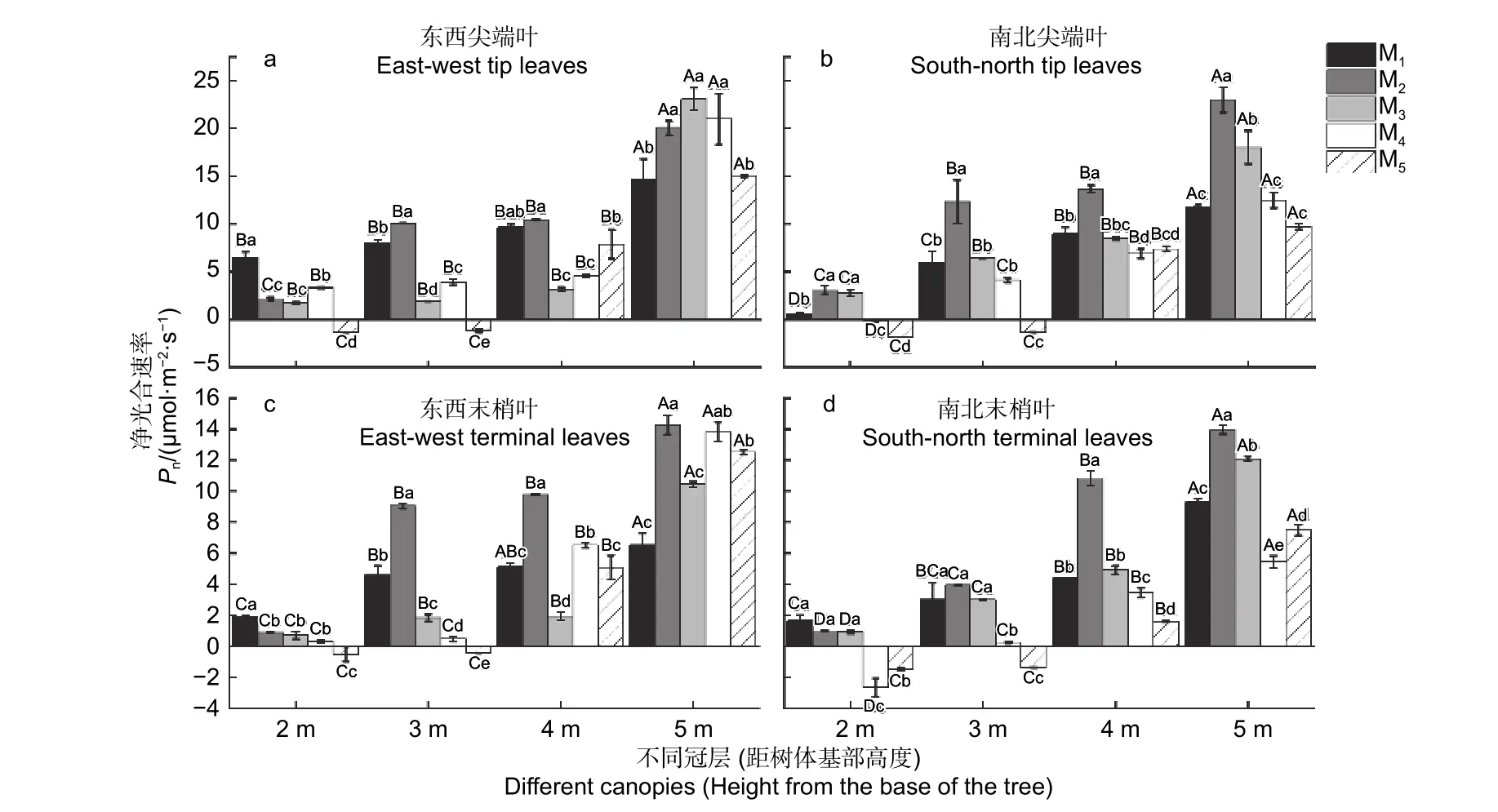
图1 不同造林密度冠层下杉木叶片的净光合速率差异分析Fig.1 Analysis on the difference of net photosynthetic rate of C.lanceolata leaves under different afforestation densities and canopy
在不同冠层中,冠层2 m 处的东西向尖端叶、东西和南北向末梢叶的Pn均在M1密度下最大(图1a、c、d),而南北向尖端叶Pn是M2密度最大(图1b)。冠层3、4 m 处,杉木东西、南北向尖端叶和末梢叶的Pn均在M2密度最大(图1a ~d)。冠层5 m 处,东西向尖端叶的Pn在M3密度下最大(图1a),而南北向尖端叶、东西和南北向末梢叶的Pn均在M2密度下最大(图1b~d)(P< 0.05)。
2.2 不同造林密度和冠层下杉木叶片气孔导度的差异分析
由图2 可知,杉木东西、南北向尖端叶和末梢叶的气孔导度Gs值在5 种造林密度下整体均表现为距树体基部下层向上层递增的趋势。在5 种造林密度下,东西和南北向尖端叶在冠层5 m 处的Gs值最大(图2a~b)。但东西和南北向末梢叶的Gs值在M1、M3、M4、M5密度下均是冠层5 m处最大,在M2密度下分别为冠层3 m 处以及4 m处为最大(图2c~d)。
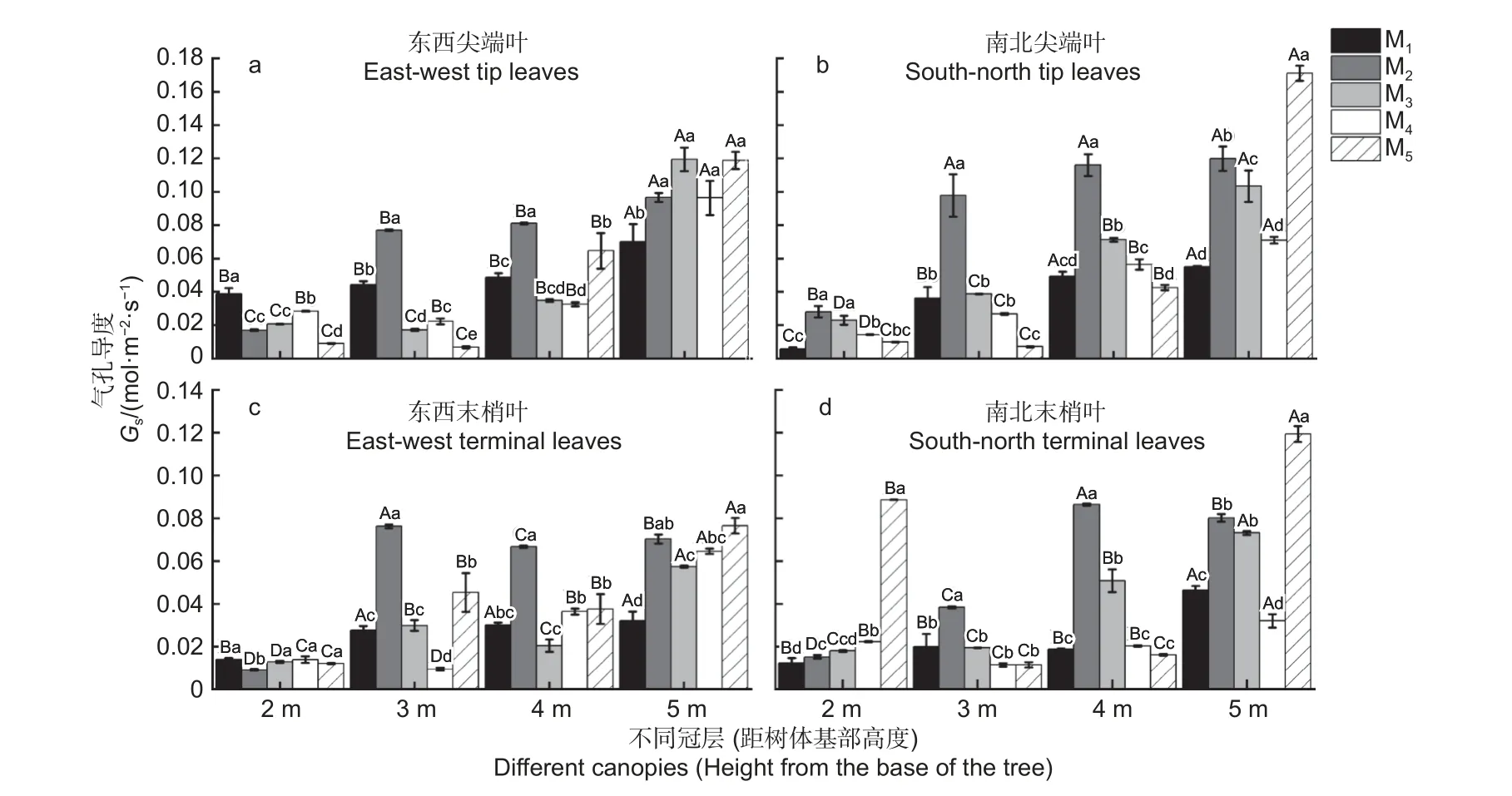
图2 不同造林密度和不同冠层下杉木叶片的气孔导度差异分析Fig.2 Analysis of stomatal conductance difference of C.lanceolata leaves under different planting densities and canopy
在不同冠层下,冠层2 m 处东西向尖端叶和末梢叶的Gs均在M1密度下最大(图2a、c),而南北尖端叶和末梢叶的Gs分别在M2和M5密度下最大(图2b、d)。在冠层3、4 m 处,东西、南北向尖端叶和末梢叶的Gs均在M2密度下最大(图2a~d)。在冠层5 m 处,东西向尖端叶的Gs在M3密度下最大(图2a),而南北向尖端叶、东西和南北向末梢叶的Gs均在M5密度下最大。
2.3 不同造林密度和冠层下杉木叶片蒸腾速率的差异分析
5 个造林密度中,杉木东西向尖端叶的Tr值在M1、M3、M4、M5密度下整体为冠层5 m 处最大,而在M2密度下整体为冠层4 m 处的Tr值最大(图3a)。南北向尖端叶Tr值在M1、M5密度下为冠层5 m 处的最大,但在M2、M3、M4密度下为冠层4 m 处的Tr值最大(图3b)。东西向末梢叶Tr值在M1密度下为冠层4 m 处最大,在M2密度下为冠层3 m 处最大,在M3、M4、M5密度下为冠层5 m 处的最大(图3c)。南北向末梢叶Tr值在M1密度下为冠层5 m 处的最大,在M2、M3密度下为冠层4 m 处的最大,在M4、M5密度下为冠层2 m 处的最大(图3d)。
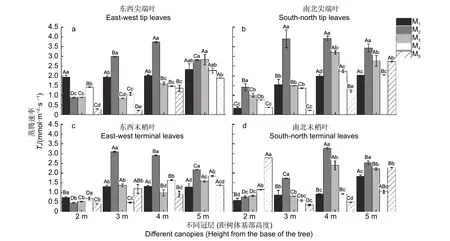
图3 不同造林密度和冠层下杉木叶片的蒸腾速率差异分析Fig.3 Analysis of transpiration rate difference of C.lanceolata leaves under different afforestation densities and canopy
在不同冠层下,东西和南北向的尖端叶与末梢叶的Tr值在冠层2 m 处分别为M1、M2、M1、M5密度下最大,且大多显著高于其他4 种密度;在冠层3、4 m 处均为M2密度的最大,且均显著高于其他4 种密度(图3a~d)。在冠层5 m 处,东西向尖端叶Tr为M3密度的最大(图3a),但南北尖端叶、东西和南北末梢叶Tr均在M2密度下最大,且均显著高于其他4 种密度(图3b~d)(P<0.05)。
2.4 不同造林密度和冠层下杉木叶片水分利用效率的差异分析
由图4 可知,杉木东西、南北向尖端叶和末梢叶的WUE值在5 种造林密度下整体表现为距树体基部下层向上层递增的趋势。不同造林密度下,冠层5 m 处的WUE值大多显著高于冠层4、3、2 m 处的WUE(P< 0.05),但在M5密度下冠层2 和3 m 处的WUE值、M4密度下冠层2 m处WUE值均出现负值(图4a~d)。
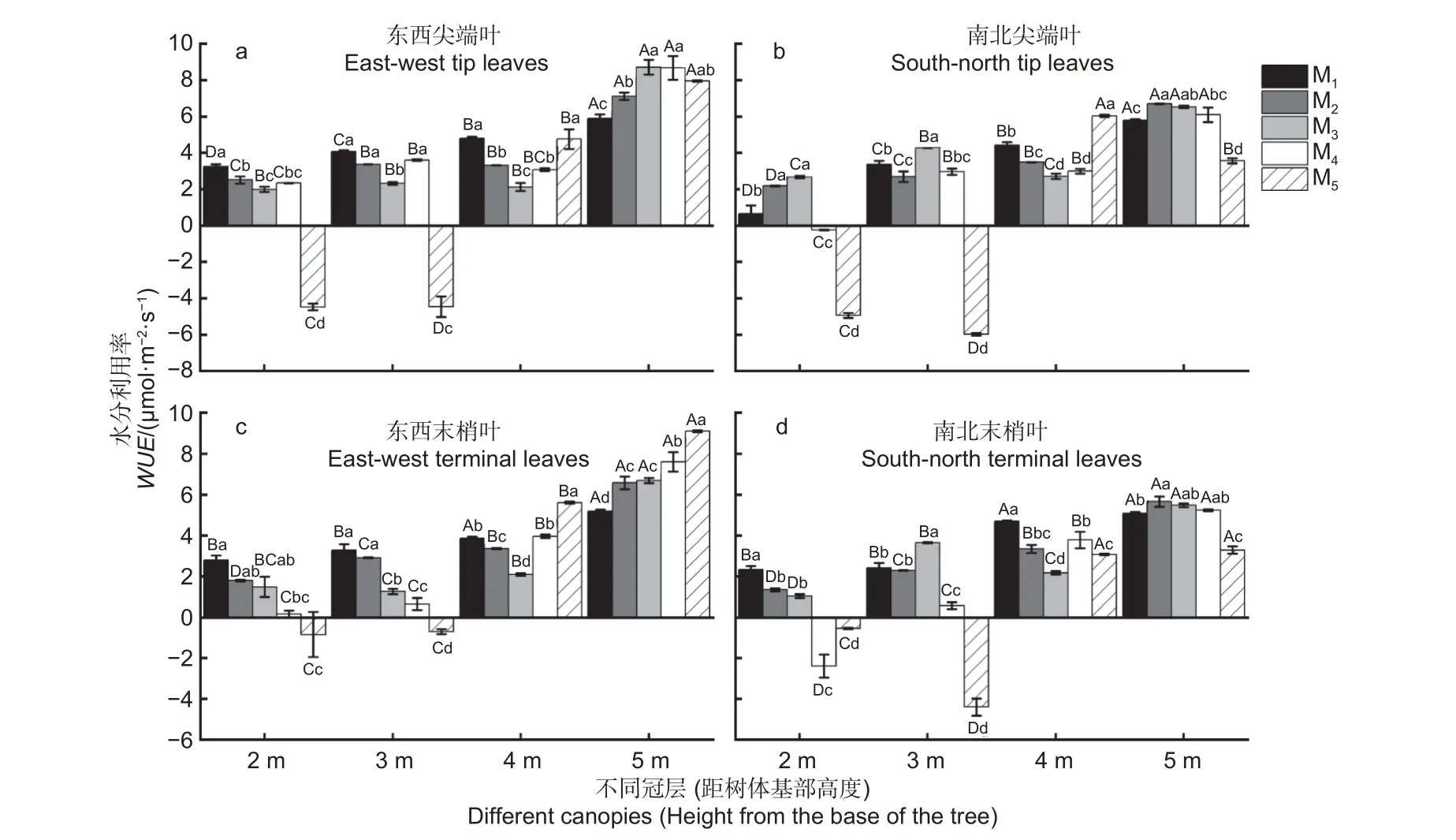
图4 不同造林密度和冠层下杉木叶片的水分利用效率差异分析Fig.4 Difference analysis of water use efficiency of C.lanceolata leaves under different planting densities and canopy
在不同冠层下,冠层2 m 处东西向尖端叶、东西和南北向末梢叶的WUE均是M1密度下最大(图4a、c、d),但南北向尖端叶WUE在M3密度下最大(图4b)。在冠层3 m 处,东西向尖端和末梢叶的WUE均在M1密度最大(图4a、c),而南北向尖端和末梢叶WUE均在M3密度下最大(图4b、d)。在冠层4 m 处,东西向尖端叶和南北向末梢叶的WUE值均在M1密度下最大(图4a、d),而南北尖端叶和东西末梢叶的WUE均在M5密度下最大(图4b、c)。在冠层5 m 处,东西向尖端和末梢叶的WUE分别在M3和M5密度下最大(图4a、c),而南北向尖端和末梢叶WUE均在M2密度下最大(图4b、d)。
2.5 不同造林密度和冠层下杉木叶片光合特性在不同方向和枝条部位间的差异分析
由表2 可知,在树冠2 m 和 3 m 处,造林密度M1、M2、M3、M4下杉木叶片在不同枝条部位上的Pn、WUE大多表现为尖端叶>末梢叶,但M5造林密度下大多表现为末梢叶>尖端叶。在5 个造林密度中,M1密度在树冠2 m 和3 m 处东西向尖端叶的Pn、WUE均为最大,分别是其他方向和枝条部位最小值的10.6、5.1 倍和2.6、1.7倍;在M2密度下,树冠2 m 和3 m 处南北向尖端叶的Pn值均最大,树冠 2 m 和3 m 处东西向尖端叶的WUE均最大。在M3密度下,树冠2 m 和3 m 处南北向尖端叶的Pn、WUE均为最大。在M4密度下,树冠2 m 处东西向尖端叶的Pn、WUE均为最大;树冠 3 m 处南北向尖端叶的Pn最大,东西向尖端叶WUE最大。在M5密度下,树冠2 m 处东西向末梢叶的Pn最大,南北向末梢叶的WUE最大,树冠3 m 东西向末梢叶的Pn、WUE均为最大。
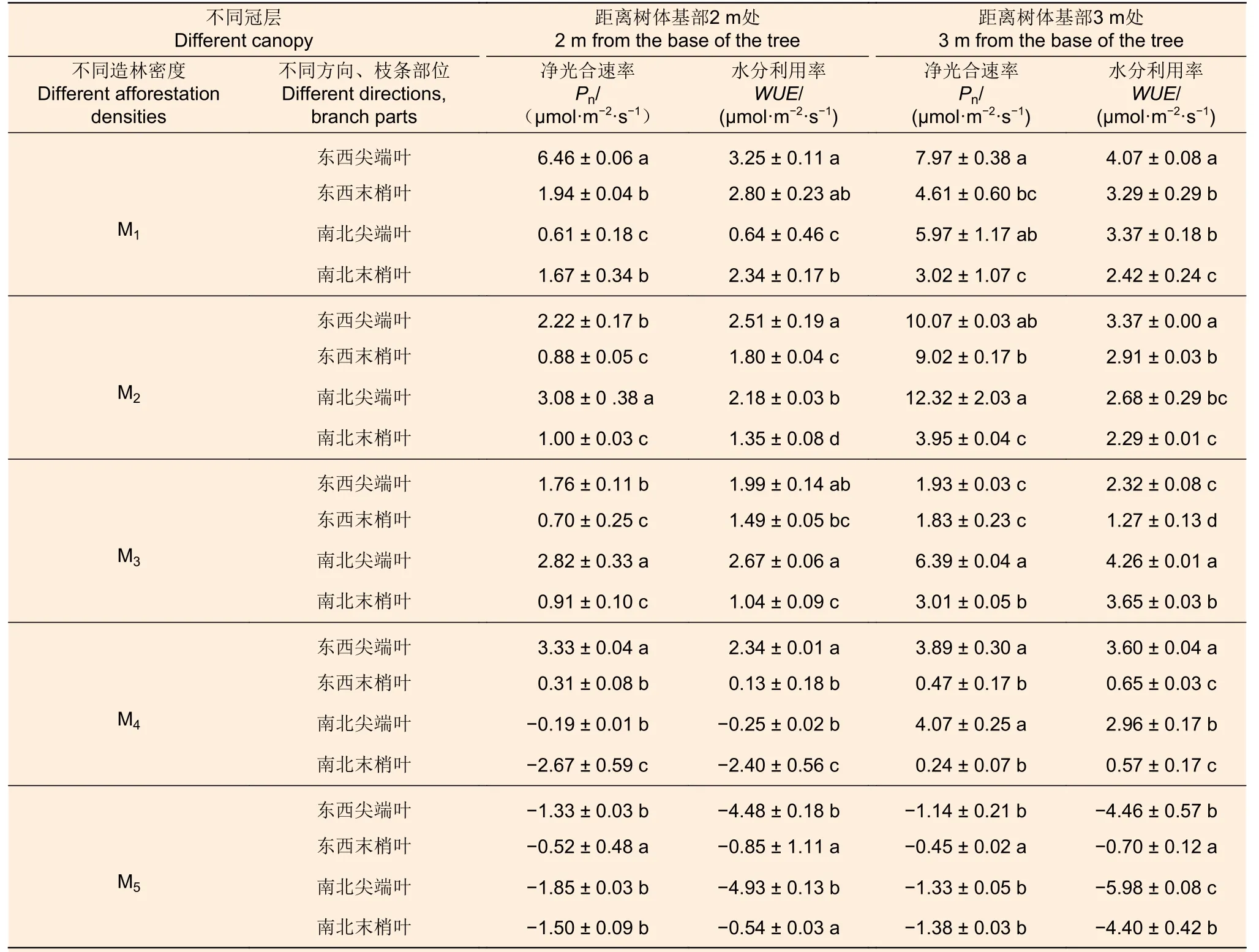
表2 不同造林密度和冠层(距离树体基部2 m 和3 m 处)杉木叶片光合特性在不同方向和枝条部位间的差异分析Table 2 Analysis of photosynthetic characteristics of C.lanceolata leaves in different directions and branches in different forest densities and canopy layers (2 m and 3 m away from the base of the tree)
由表3 可知,树冠4 m 和5 m 处,5 个密度下杉木不同枝条部位上叶片的Pn、WUE总体表现为尖端叶>末梢叶。在M1 密度下,树冠4 m 和5 m 处东西向尖端叶的Pn、WUE为最大,分别是其他方向和枝条部位最小值的2.2、1.2、2.2、1.2 倍。在M2 密度下,树冠4 m 和5 m 处南北向尖端叶的Pn为最大,但WUE的差异不显著。在M3 密度下,树冠4 m 处南北向尖端叶的Pn、WUE均最大,树冠5 m 处东西向尖端叶的Pn、WUE均最大。在M4 密度下,树冠4 m 处南北向尖端叶的Pn最大,树冠5 m 处东西向尖端叶的Pn、WUE最大,分别是南北末梢叶(最小值)的3.8、1.7 倍。在M5 密度下,树冠4 m 和5 m 处东西向尖端叶的Pn最大。

表3 不同造林密度和冠层(距离树体基部4 m 和5 m 处)杉木叶片光合特性在不同方向和不同枝条部位间的差异分析Table 3 Analysis of photosynthetic characteristics of C.lanceolata leaves in different directions and branches in different forest densities and canopy layers (4 m and 5 m away from the base of the tree)
由表4 可见,不同密度,不同冠层,不同方向,不同枝条部位及其交互作用均对杉木叶片的光合特性指标有极显著影响(P<0.01),但不同方向和不同枝条部位的交互作用对杉木的光合特性指标均无显著影响(P>0.05)。
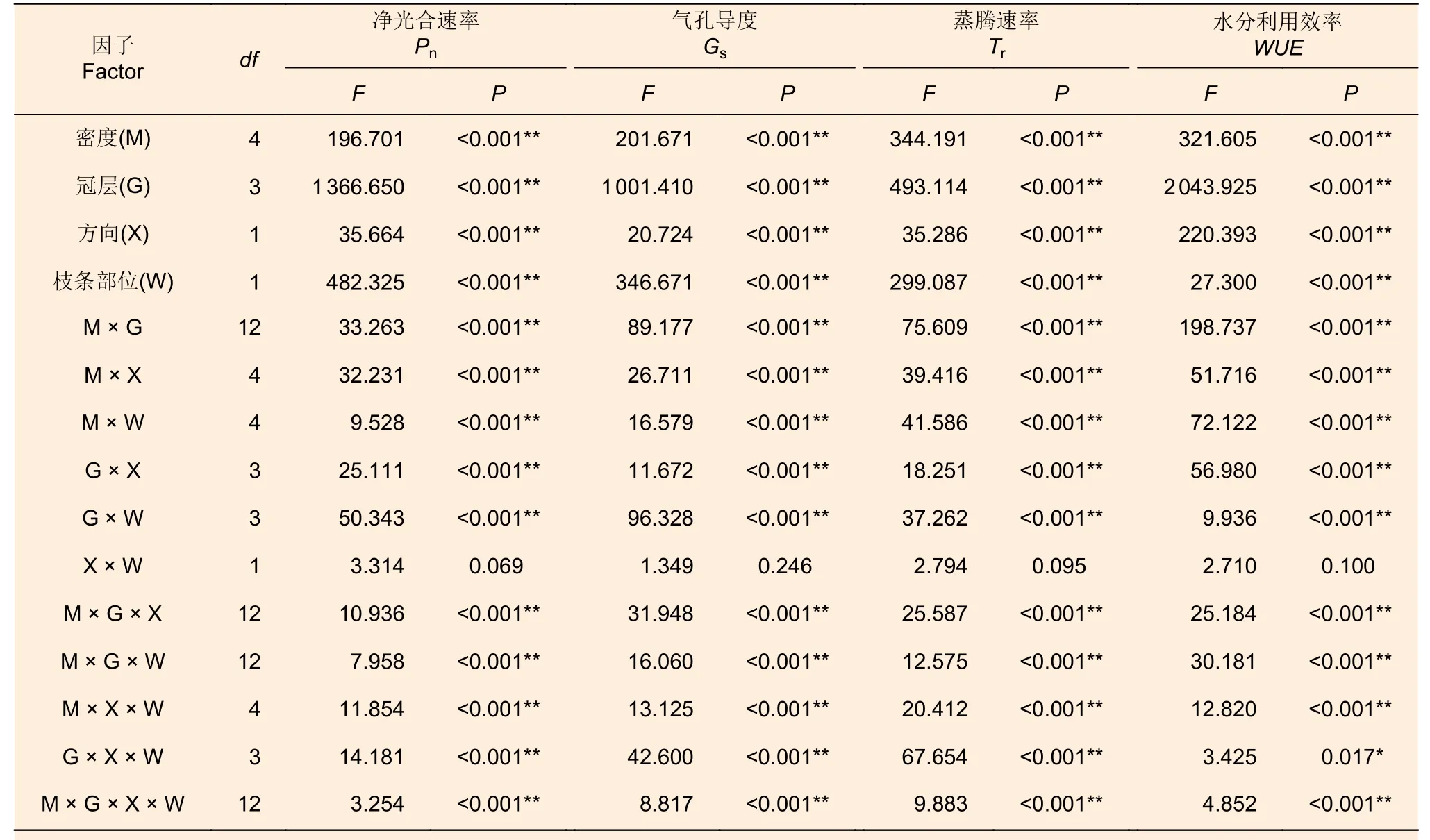
表4 不同密度、冠层、方向和枝条部位对杉木光合指标和水分利用效率影响的多因素方差Table 4 Multivariate variance of the effects of different densities, canopy layers, directions and branch positions on photosynthetic indexes and water use efficiency of C.lanceolata
3 讨论
3.1 不同造林密度对杉木叶片光合特性的影响
植物光合作用对林木生长发育具有重要的影响,特别是林木树冠光合作用的空间异质性研究可为林木光合能力进行精确评估提供科学依据[13]。同时,确定造林密度是人工林培育环节中的一个重要环节,合理的造林密度是林木获取养分资源和保证生长发育的前提[14-15]。本研究通过对杉木幼林不同造林密度、冠层、方向、枝条部位叶片进行系统的光合特征指标的测定[16]。研究发现,冠层2 m 处M1密度的Pn、WUE反而更高,说明低造林密度以及下冠层对弱光的利用率要分别大于高造林密度以及上冠层,这与骆丹等[17]对西南桦(Betula alnoidesD.Don)、李勇[18]等对油松(Pinus tabuliformisCarrière)、刘强[19]等对长白落叶松(Larix olgensisA.Henry)等研究的结果基本一致,可能是杉木低冠层长期处于弱光环境,且低造林密度的林分具有较长的树冠及较大的冠幅,上冠层枝叶的遮挡使光辐射透过率较低,下冠层光环境更弱。在冠层3~5 m 处,Pn、Tr、Gs、WUE值随造林密度增大总体呈现出先升后降的趋势,这与赵连春等[20]人对怪柳(Tamarix chinensisLour.)光合特性的研究结果基本一致,M2密度达到最大值说明不同造林密度对杉木光合特性有显著的影响,合理的造林密度对于培育高效优质的杉木人工林有很关键作用[21]。此外,在树冠2、3 m 处Pn、WUE出现负值,这可能是与M4、M5密度较大,使杉木叶片之间相互遮阳逐渐形成不利于叶片光合作用的环境[22],表明较高的造林密度会导致光照在林分中的分布变得不均匀。在高密度林地中,上层树冠相互竞争光照资源,使下层枝叶接受到较弱的光照,从而降低下层枝条的光合作用效率[3],故可考虑进行适当营林措施优化树冠结构,以促进杉木的生长和发育。
3.2 不同冠层对杉木叶片光合特性的影响
研究表明杉木叶片光合特性的影响因素很多,但多数学者认为光环境变化是造成不同部位叶片光合能力差异的主要原因[23-25],Baldochhi 等认为,光在林木冠层中的传递和分配方式适用于比尔定律[13],即光照强度从树冠2~5 m 呈指数函数递增,周围的空气温度和湿度也随之变化,使不同冠层形成不同的微气候环境。在本研究中,杉木各光合指标在4 个冠层上有明显的差异,随冠层升高其光合指标Pn、Gs、Tr、WUE显著增大,这与郑元[16]和孟陈[10]等人研究结果一致,造成这种差异可能与光照强度、叶片中的叶绿素和叶氮含量在冠层垂直分布的差异有关[26-28]。因此,依据不同冠层枝条叶片的光合特性的差异,对下冠层光合能力较差的枝条进行营林措施处理,可提高其光合能力。本研究发现树冠3 ~4 m 差别较小,而树冠2 和5 m 差异显著,产生这种差异主要与其所处的光环境有关,树冠5 m 处的枝叶光照条件充足,而树冠2 m 的枝叶获得的透射光或散射光较少,且强度较弱[17]。此外,提供给光合作用的物质能量和水分主要来源于蒸腾作用,光环境不仅对光照条件有直接影响,且对水分、温湿度等也有间接影响,进而对蒸腾速率和水分利用效率产生影响[18],本研究发现不同冠层叶片所处的光照条件的差异[29-30],不仅影响各冠层的光合速率,还对蒸腾和水分利用效率均产生了显著的影响。
3.3 不同方向和不同枝条部位对杉木叶片光合特性的影响
一般认为,气孔是植物进行CO2和水汽交换的一个重要通道,反映这种交换能力的一个关键指标就是Gs[31],而Tr和Gs之间有着非常紧密的联系,二者都是影响光合速率的重要生理指标[32],在本研究结果中杉木叶片Gs、Tr对Pn的影响表现出较大的差异。另外,不同造林密度和冠层的杉木光合特征在不同方向上差异均显著,这与郑元[16]对刺槐(Robinia pseudoacaciaL.)的研究结果不一致,可能是郑元等人将树冠分为东、西、南、北4 个方位进行测定,而本实验采用东西向与南北向两个方位测定;也可能是所选用林木的龄级不同使本实验结果与郑元等人研究不一致。
在自然条件下,人工林中林木同一枝条不同部位针叶的净光合率差异显著[23]。本研究发现,Pn、WUE在不同枝条部位上均表现为尖端叶>末梢叶,这与郭继善[33]和肖文发[23]等的研究结果基本一致。由于不同部位的枝叶对稳态和动态光照的反应与其对所处光环境的长期适应有一定关系,处于枝条末梢的叶片靠近树干,冠层浓密,光照强度较弱,而处于枝条尖端的叶片光环境相对优越[24-25]。但本研究发现在M5较高密度下树冠2~3 m 处的光合指标表现为末梢叶大于尖端叶,这可能是因为造林密度过大,冠层上部和周围林冠的长期遮阳使光线因子失去光合时的主导性[7]。
4 结论
整体上,5 年生杉木叶片在冠层2 m 处的Pn、WUE随造林密度的增加而减小,在冠层3~5 m 处,Pn、Gs、Tr、WUE随造林密度的增加呈现先升后降的趋势。Pn、Gs、Tr、WUE均随冠层的升高而显著增大,树冠5 m 处的枝叶光照条件充足,光合作用较强。5 个造林密度中M2密度(1 667株·hm-2)的杉木叶片在不同冠层和枝条方向上的光合能力整体上更强,在4 个冠层中各造林密度下均为树冠5 m 处的杉木叶片表现出更大的光合潜力,尖端叶的光合能力强于末梢叶,东西向枝条叶片光合能力强于南北向枝条叶片。在M5造林密度(6 667株·hm-2)的冠层2 m 和3 m 处、M4密度造林(3 333株·hm-2)的冠层2 m 处的枝条叶片净光合速率出现负值,即密度的增加会对森林内光照资源产生影响,特别是位于低冠层的枝叶,且密度越大,冠层受到的影响也越明显。
