黄河首曲湿地植物群落生产力、物种多样性及其与生境的关系
2024-01-17胡凤烨赵锐锋张丽华杨尖拉毛当知才让蒋小玉
胡凤烨, 赵锐锋, 张丽华, 王 宏, 杨尖拉毛, 当知才让, 蒋小玉
1 西北师范大学地理与环境科学学院,兰州 730070 2 甘肃省绿洲资源环境与可持续发展重点实验室,兰州 730070 3 甘肃省玛曲青藏高原土著鱼类自然保护区管理局,玛曲 747300 4 甘肃黄河首曲国家级自然保护区管护中心,玛曲 747300
湿地是水陆相互作用下形成的独特生态系统[1],具有维持生态平衡、调节气候、保护生物多样性的作用,是世界生物多样性重要的储存库,因此,被称为“生物基因库”和“地球之肾”[2]。植物多样性是衡量群落结构与功能复杂性的一种指标,涵盖了物种的丰富程度,单位面积内物种的数量以及物种比例等,能够较好地反映群落结构与生境多样性,对生物多样性的保护具有重要作用[3]。湿地植物多样性受环境因素的影响,包括气候[4]、土壤[5-6]和水环境[7]等,其中土壤作为植物生长的物质基础,对植物分布格局和物种多样性都有重要影响[8]。近年来,随着气候变化和人类活动的增加,导致湿地面积急剧减少,湿地退化问题日益严重[9]。湿地退化引发的土壤退化不仅会导致土壤的基本理化性质发生改变,还会对养分元素生物地球化学循环的变化产生影响,进而影响湿地植物的生长和生产力[10]。湿地生态系统已成为世界上最受威胁的生态系统之一[11]。
近年来,生态学家对持续的生物多样性丧失是否会对生态系统的功能产生影响的问题越来越关注,使得生物多样性与生态系统功能关系的研究是当今生态学研究的主要热点之一[12-13]。生产力作为生态系统功能的综合指标[14],是研究生物多样性与生态系统功能关系的有效途径。目前,人们对植物多样性与生产力之间的关系仍存在争议[15],关于生产力-多样性关系大致分为三类:(1)单峰关系,其中多样性随植物生产力的增加呈现先升高后降低的变化[16];(2)单调关系,多样性与生产力为正或负线性[17]以及(3)不显著关系[18]。关于争议,一方面是由于在研究生物多样性和生产力之间的关系时,将植物多样性的不同组成部分考虑在内,并发现不同类型的多样性指数与生物量之间相关系数可能会产生很大差异[19]。另一方面,在自然生态系统中气候、地理、土壤条件和人类活动等生物与非生物因素的交互作用,使生产力与植物的多样性关系更加复杂[20]。具体而言,降水使非优势物种对资源更有效利用,一定程度上提高生物量,进而影响生产力与物种多样性的关系[21];生产力与多样性的关系受坡向与海拔的影响[22];不同土壤因子可相应调节各种生态过程[23],且有限的养分供应影响着生物量生产;人类活动尤其是放牧,放牧强度和放牧年份改变着多样性-生产力的关系[24]。由于环境扰动,自然群落生物多样性影响比实验中的弱[13]。小规模的实验研究可以控制这些环境因素,并提供对多样性-生态系统功能关系的见解,但对于得出的结论能否广泛地应用于更大尺度地实施保护管理战略中,尚存疑虑[25]。因此,探索湿地植物多样性与生产力之间的关系及影响因子,将有利于湿地植物多样性的保护和恢复,对保障区域的生态平衡和可持续发展具有重要作用。
青藏高原(QTP)是泛第三极的主体,平均海拔>4000 m,素有“世界屋脊”和“亚洲水塔”之称,作为世界重要的生态区,是高寒生态系统物种及遗传基因最丰富和最集中的地区之一,是研究物种多样性与生产力关系的理想场所[26]。由于该地区的脆弱性、对气候变化的敏感性以及不断加剧的人类活动,面临严峻的退化演替形势[27],生物多样性受到威胁。高寒湿地作为一种特殊的湿地类型,具有蓄水调节、生物多样性保护和生态环境改善等重要功能[28]。黄河首曲湿地作为青藏高原最具代表性的内陆高寒湿地,是黄河上游重要的水源涵养区和补给区,素有黄河“蓄水池”、“高原水塔”之称[29]。因此,研究高寒湿地植物多样性与生产力之间的关系可为维持生态系统服务,特别是植物生产提供理论依据。然而物种多样性与生产力的关系并不总是一致的,高寒湿地退化导致的物种丧失如何影响高寒生态系统功能的内在机制并不完全清楚。在此背景下,本文以黄河首曲高寒湿地为研究对象,探讨以下3个主要问题:(1)黄河首曲高寒湿地植物多样性和生物量的地理梯度变化遵循何种规律?(2)物种多样性指数与植物生物量生产之间的关系;(3)高寒生态系统植物多样性、生产力与土壤因子关系。以期为我国高寒地区的植被多样性与生物量空间分布模式与影响因子的研究提供典型范例的同时,为高寒湿地植物多样性保护和生态系统稳定性维持提供理论参考。
1 材料与方法
1.1 研究区概况
研究区黄河首曲湿地国家级自然保护区(101°54′12″-102°28′45″ E,33°20′01″-33°56′31″ N)是典型的青藏高原高寒湿地,在行政区上划为甘南藏族自治州玛曲县,地处青藏高原东部边缘,素有黄河“蓄水池”和“中华水塔”之称,是黄河的重要水源涵养区和补给区[30]。黄河自青海流入玛曲县,经县境西、南、东、北环流全县,形成一个流程长达 433.7 km 的 U 形大拐弯,谓之“黄河首曲”。海拔高度3391-4145 m,地势西高东低,由西北向东南倾斜。低气温、短生长季、大风、寒冷与大风所致的土壤冻结和生理干旱是该区最为显著的气候特点。该区属高寒半湿润气候,年平均气温为1.1℃,年降水量为672 mm,降雨集中在7-8月份。年日照时数约为2580 h,年无霜期平均少于100 d。土壤类型为高山草甸土,境内湿地集中的曼日玛、采日玛镇多发育沼泽土和泥炭土。该地区天然植被类型为典型高寒草甸,主要以禾本科、莎草科、菊科、毛茛科等多年生草本植物为优势种,如禾本科的垂穗披碱草(Elymusnutans)和早熟禾(Poaannua);莎草科的线叶蒿草(Kobresiacapillifolia)、华扁穗草(Blysmussinocompressus);菊科的长毛凤毛菊(Saussureahieracioides)、黄帚橐吾(Ligulariavirgaurea);毛莨科的唐松草(Thalictrumaquilegifolium)和条叶银莲花(Anemoneobtusiloba)等,并伴有其它双子叶类杂草[31]。
1.2 植被调查、采样与样品分析
依据地上生物量波动规律,选择于2021 年 7 月末至 8 月初在植物生长的旺盛期进行野外调查取样。为全面客观地反映自然保护区植被与土壤特征,沿保护区边界大致以5 km为间距设置样地,使得样地尽可能均匀分布,共调查52个样方,每个样方均用GPS定位,并记录经纬度、海拔高度、周围环境特征、距河道的距离及人类干扰等。现场鉴定样方中的植物物种,并将个别现场无法识别的物种采集标本带回实验室鉴定。记录个体数、物种数及高度,样方总盖度及不同种植物的高度和分盖度,将样方内所有植物按物种贴着地面从基部用剪刀剪取植物地上部分,装入信封袋带回实验室,经105℃杀青处理,置入65℃烘箱中加热烘干至恒重即得到地上生物量。用土钻取0-10 cm、10-20 cm、20-40 cm的土壤样品,一部分装入铝盒用于土壤含水量的测定,另取一份均匀混合好的土样每层各约1 kg装袋,及时带回实验室在阴凉通风处自然风干,进行土壤理化特性指标分析。
将装入铝盒的土壤样品使用校对后的电子天平测量鲜重,然后将土壤样品置于已预热至(105±5)℃的烘箱中烘烤6 h左右,取出样品待冷却至室温迅速称重,重复操作,继续烘烤0.5-1 h,再称重,直至恒重(前后两次质量差不超过1%)。测定重复2-5次,取平均值。土壤含水量计算公式如下:

将装袋的土壤样品带回实验室自然风干,剔除较大的根系、砾石等,研磨过筛至1 mm和0.15 mm粒径后,用常规法测定各层土壤理化性质并取平均值。土壤 pH 值由pHS-3C型酸度计测定(水土比2.5∶1),电导率(Electrical conductivity,EC)由电导率仪测定(水土比5∶1);土壤有机碳(Soil organic carbon,SOC)含量采用重铬酸钾-外加热法测定;土壤全氮(Total nitrogen,TN)采用半微量凯氏定氮法测定。以上实验均在西北师范大学土壤地理实验室完成。
1.3 植物多样性的测定
基于重要值计算以下4 种指数测度植物多样性:
重要值=(相对高度+相对盖度+相对多度)/3
选择了四个物种多样性指数,Patrick 丰富度指数(R)、Shannon-Weiner 多样性指数(H)、Pielou 均匀度指数(J)和 Simpson 优势度指数(D),被用来描述高寒湿地植物物种多样性。具体计算公式如下[32]:
Patrick 丰富度指数:
R=S
Shannon-Weiner多样性指数:
Pielou均匀度指数:
Simpson优势度指数:
式中,S为群落中的物种数,Pi为相对重要值。
1.4 数据处理
采用 Microsoft Excel 2010录入所有原始数据,首先采用描述性统计分析,包括最大值、最小值、平均值、标准差(SD)和变异系数(CV)来识别植物群落特征变化趋势;采用 Origin 2021 绘制植物生物量、物种多样性、盖度和土壤因子的三维(经度、纬度和海拔)空间分布图和线性回归图;用 SPSS 26.0统计分析软件对土壤元素、地上生物量与物种多样性之间的关系进行Pearson相关分析。图中和表中每个指标的表现形式均为平均值加减标准误差。
2 结果
2.1 湿地植物科属种组成
样方调查统计结果显示,研究区52个样方中共出现植物25科70属94种(表1),以多年生草本为主体,蕨类植物 1 科 1 属 2 种,单子叶植物5科13属18种,双子叶植物19科56属74种。植物科内属、种的组成差异较大,其中以菊科种类最多,为16 种,占种组成的17.02%;其次为禾本科,共12种,占种组成的12.77%;然后依次为毛茛科(10种)、龙胆科(9种)、莎草科(6 种)、豆科(6 种)等。各科植物中有10科为1科1属1种,占总科数的 40%,说明研究区湿地植被不是以某一个或几个科植物占主导。

表1 湿地植物物种组成Table 1 Species composition of wetland plants
2.2 湿地植物物种多样性与生产力的关系
研究结果表明,Patrick丰富度指数、地上生物量随空间维度(经度、纬度和海拔)变化较大,变化范围分别为5.2-111.2 g/m2和3-21,变异系数为46.42%、54.75%;植被盖度和Shannon-Weiner多样性指数变异系数居中,分别为25.97%、31.92%,数值变化范围分别从0.49到2.9、20%到90%。均匀度指数和Simpson优势度指数空间变异性较小,且数值相对较高,均值分别为0.8、0.75(表2)。

表2 植物群落特征Table 2 Characteristics of plant community
从图1空间分布格局来看,Shannon-Weiner多样性指数和Patrick丰富度指数峰值均出现在保护区东部(靠近黄河支流)地区;且保护区东部比西部(海拔较高)地区具有较高植被盖度;植物群落地上生物量和物种多样性在经纬度和海拔上均不存在线性分布。植物生物量、物种多样性的峰值均出现在海拔3400-3500 m的保护区东北部。
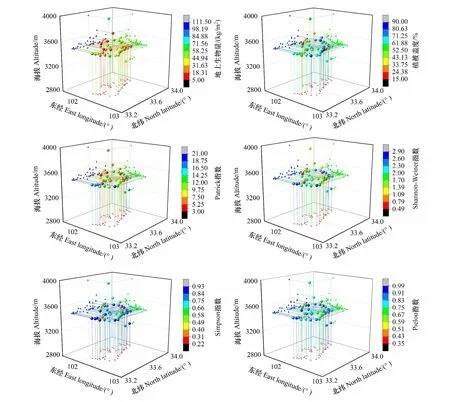
图1 植被生物量、盖度和物种多样性指数的三维空间分布格局Fig.1 The three-dimensional spatial distribution pattern of vegetation biomass, coverage and species diversity index
由表3相关性分析得出,多样性指数之间均呈现极显著正相关(P<0.01),因为4种多样性指数反应的群落特征具有一致性;Patrick丰富度指数、Shannon-Weiner多样性指数与地上生物量存在显著的正相关关系,表明地上生物量随植物物种多样性的增加而显著增加。植物盖度与4种多样性指数之间均呈显著正相关(P<0.05),与Simpson优势度指数相关性最高,其相关系数为0.26;其次为Shannon-Weiner多样性指数和Patrick丰富度指数,相关系数分别为0.25、0.22(图2)。

图2 植物多样性指数与地上生物量、盖度的关系Fig.2 The relationship between plant diversity index and aboveground biomass, coverage
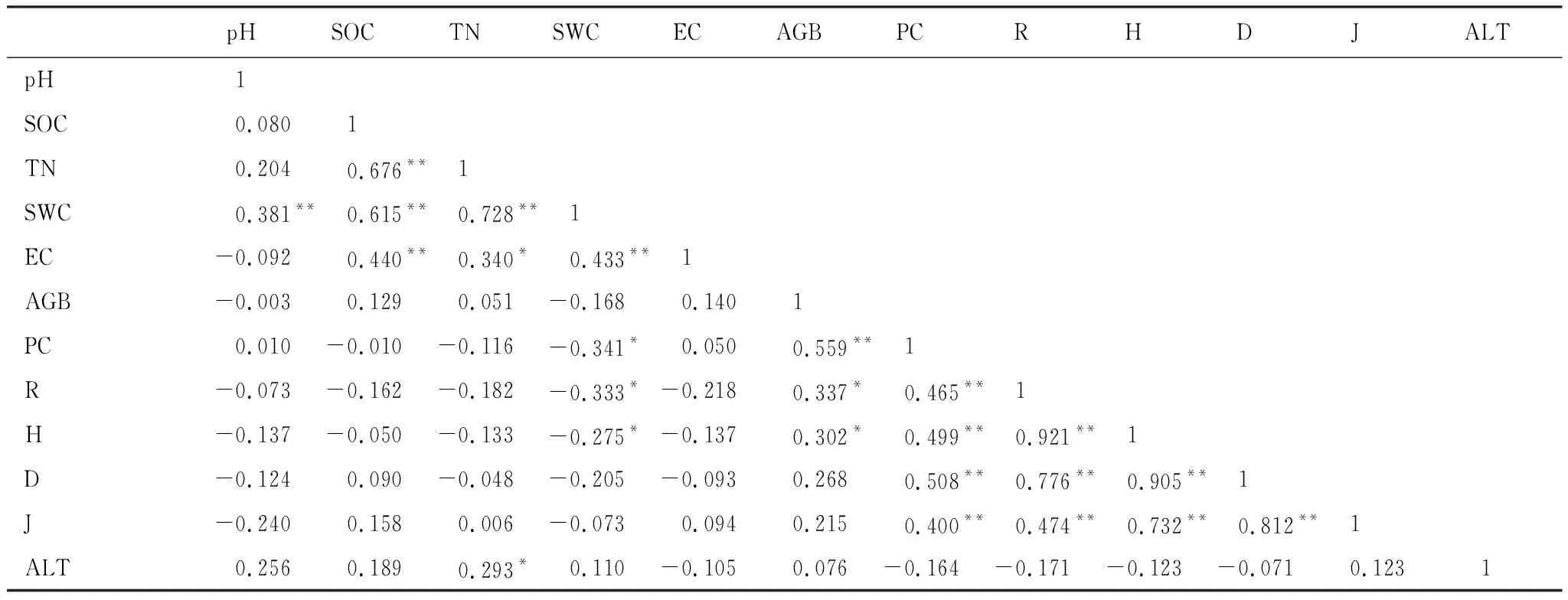
表3 高寒湿地群落土壤理化性质与物种多样性和生物量间的相关性Table 3 Correlation coefficients among soil physicochemical properties,species diversity and biomass of alpine wetland community
2.3 土壤因子对物种多样性、生产力的影响
对于土壤因子,保护区全氮含量呈现出由北向南增加的趋势,并随海拔升高而增加,全氮与海拔呈显著正相关(表3),表明土壤全氮具有一定的空间相关性,变化范围为0.18-7.10 g/kg;pH、电导率表现为保护区北部高于南部,变化范围分别为5.3-8.68、203.5-59 μS/cm;土壤有机碳和含水量变化幅度较大,分别约为11.00-116.50 g/kg、5.00%-224.00%,峰值均出现在海拔3500-3600 m的保护区西南部,空间分异规律不明显(图3)。

图3 土壤因子的三维空间分布格局Fig.3 Three-dimensional spatial distribution pattern of soil factors
pH与土壤有机碳、全氮之间存在极显著负相关(P<0.01)(表3);如图4所示,土壤含水量与植被盖度、Patrick丰富度和Shannon-Weiner多样性指数呈显著负相关关系,说明随土壤含水量的升高,群落的多样性将呈现出线性减少的趋势,进而抑制湿地植物的生长;其他土壤因子与植物物种多样性相关性不显著。
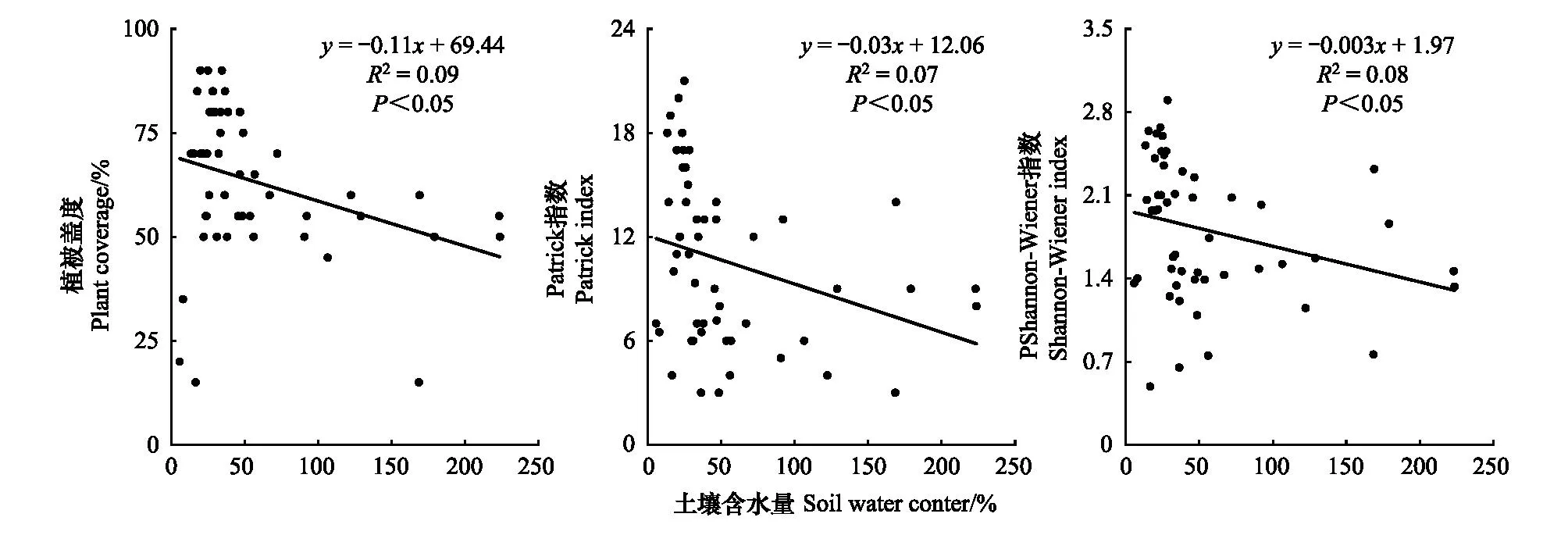
图4 土壤含水量与植被盖度、丰富度和多样性指数的关系Fig.4 The relationship between soil water content and plant coverage, richness index and diversity index
3 讨论
植物群落的物种组成作为反映其结构变化的重要指示因子,是了解群落的基础和群落性质的关键[33]。在本研究调查的52个样方共出现植物94种,隶属于25科70属,可见黄河首曲湿地植物组成比较丰富。相较于在湿润、半湿润地区的平陆黄河湿地植物67 科213 属[34],该湿地的植物种类组成较少;但略高于在内陆干旱区的塔里木河中下游湿地和艾里克湖湿地植物15 科35 属[35]。土壤类型与湿地类型决定了该区植物群落组成[36],湿地植物种类主要集中在菊科、禾本科、豆科、莎草科,其他科的物种相对少且分散,其中菊科、禾本科物种数较多,分别为16和12种,莎草科和豆科物种数相对较少,均为6种,该结果也从侧面反映出湿地植物群落种类组成的变化,呈现逆向演替方向,即杂类草比例增加,而莎草和禾草比例减少[37],这与其他高寒生态系统在退化过程中的演替规律相一致[38],可能是由于放牧作为保护区最主要的人为干扰方式,显著影响植物多样性和植物群落的覆盖模式。
随着植物多样性指数类型不同,物种多样性与生产力之间的关系存在差异。本研究检测出两种关系,即正向和非线性关系。Patrick丰富度、Shannon-Weiner多样性指数与地上生物量呈显著正相关,这与早期研究结果一致[39],这表明在小尺度研究上,地形虽然引起了土壤水热因子的变化,但物种多样性变化更为明显,初级生产力受生物群落内部调节的作用较大,群落内部对物种组成结构的调节更为积极,更多的物种可以利用微小的环境异质性产生的资源差异而获得生存;另一方面放牧作为保护区最主要的人类活动方式,“牲畜的选择性采食”[40]使得占据群落上层的禾本科植物高度降低,遮阴效果减弱,增加了杂类草接受光照的机会,从而造成大量的毒杂草(特别是菊科)的入侵,在一定程度上增加了地上生物量和物种多样性[41],提高植物的气孔导度、光和水分利用效率,从而提高植物的光合净速率和初级生产力[42];而过度放牧显著降低了优势种的高度,使得根系较深的杂类草在环境变恶劣时更好的适应环境的变化,进而实现植物优势种的转变,通过降低植被生存状态来降低生产力[43]。Simpson优势度指数、Pielou均匀度指数与地上生物量呈非线性关系,这与陈宁和牛钰杰所得出群落生产力和物种多样性的非线性响应的研究结果相似[18, 22]。出现这一结果,究其原因,可能来自两方面。第一,Pielou均匀度指数表示物种间丰富度的相似程度,相对独立于生境的资源水平,更多地受制于群落本身动态的影响[44];第二,气候暖干化下的湿地退化,引起植物群落类型发生变化,群落优势种重要值的逐渐降低,其在群落内绝对支配地位逐渐削弱,而毒杂草获取资源能力则会相对增加,在群落中表现为比例升高,使得优势度指数和均匀度指数出现波动[45]。
在自然生态系统中,地形、气候、光照、温度等环境因子与取样方式的综合作用,使得植物多样性与生产力之间的关系更为复杂。植物群落是相互联系的单元,以往的研究显示,由于纬度和海拔的改变,植物种类的多样性发生了显著地改变。更确切地说,在高海拔高寒生态系统方面,有学者认为,高寒生态系统植物的物种多样性随着纬度和经度的增加而增加,随着海拔的升高而降低[46],或者是在中等海拔的“膨胀现象”[47]。与之相比,本研究结果表明植物多样性和生物量在经纬度和海拔等方面未表现出显著差异,空间变异性、生境差异和资源供给程度可能是造成植物群落结构和组成变化的主要原因[22]。一些对于土壤因子与物种多样性和生产力关系的研究中得出,有机碳、全氮等土壤养分与物种多样性、生产力呈显著正相关[23, 46, 48],而本研究中并未表现出很强的依赖性,这与Bi等人研究一致[49]。之前研究表明,土壤养分随着植被盖度和地上生物量的增加而增加[50],也有研究发现二者没有相关性或呈现负相关[51-52]。出现不同结果的原因可能是由于首曲湿地泥炭地土壤中丰富的有机质,削弱了有机碳对植被覆盖和物种多样性的影响[23];另一个原因是鼠害啃食牧草,破坏了土壤结构和原有植被,加剧湿地退化,从而影响高寒湿地植物物种分布以及促进群落进一步演替[44]。湿地的水文动态是决定植物群落发展和植物分布模式的主要因素。一般来说,土壤含水量一方面通过影响养分的可利用性影响植物分布,进而影响物种多样性,物种多样性的增加导致土壤碳输入和氮矿化速率增加[53],有机化合物更加多样化,微生物分解加强,产生更多的细胞外酶[54],从而提高了土壤养分含量,增加群落的生产力;另一方面土壤水分的更高可用性一定程度上可增加植物生物量,带来更大的生长潜力[55]。而本研究中,土壤含水量与植被盖度、Shannon-Wiener多样性指数和物种丰富度指数呈显著负相关,这与前人研究结果一致[44, 56],一方面是因为植物和土壤呈现出“异步性”,随着优势种逐渐减少,生物量较多的杂类草会弥补,尽管物种多样性和生物量没有较大的变化,但群落结构和盖度的变化使土壤物理化学性质发生明显变化;另一方面黄河首曲高寒湿地长期积水区土壤的通气条件较差,只有一部分耐水淹胁迫种处于优势,倾向于形成较单一的植物群落[44]。在光捕获以及水和氮利用方面的物种互补性可能是自然组装群落中生物多样性-生产力关系的基础[57],植物多样性对生产力的影响在复杂的自然生态系统中比在实验中要弱[13, 58]。土壤因子的复杂性使得与植物的生物量、生物多样性之间的相关关系受到了很大的挑战,还需要通过控制实验进行长期观察和探讨。
4 结论
综上所述,植被物种多样性、群落生产力和环境因子之间相互作用、相互影响,共同影响植物群落的分布格局。黄河首曲高寒湿地植物物种多样性和生物量未表现出明显的纬度、经度和海拔梯度变化;在研究植物多样性对生物量生产的影响时,因多样性指数的不同而呈现不同的关系模式,植物生物量与丰富度指数和Shannon-Weiner 多样性指数呈显著正相关,与Simpson优势度指数和Pielou均匀度相关性不显著;在自然植物群落中植物多样性与生物量生产之间的关系更加复杂,土壤含水量显著影响着高寒湿地的植被覆盖和物种多样性。文章结果表明,对于小尺度研究区域,地形虽然引起了土壤水热因子的变化,但物种多样性比土壤因子造成生产力差异的影响更为明显。因此,在实际高寒湿地的保护和恢复中需重点考虑植物多样性和土壤含水量对高寒生态系统功能的影响。
