黄土丘陵区小流域不同海拔刺槐径向生长对气候的响应差异
2024-01-17刘亚玲信忠保李宗善买尔当克依木
刘亚玲,信忠保,*,李宗善,买尔当·克依木
1 北京林业大学水土保持学院,北京 100083 2 中国科学院生态环境研究中心城市与区域生态国家重点实验室,北京 100085
水热条件变化是影响树木生长的主要外界环境因素,不同海拔高度具有不同的水热条件,进而影响树木的径向生长[1]。大部分研究显示,同一树种在不同海拔梯度水平上受气候变化的影响存在很大区别,一般情况下高海拔树木生长受温度因素限制,低海拔受降水的限制。山西芦芽山中高海拔华北落叶松(Larixprincipis-rupprechtii)生长受低温限制,低海拔生长遭受干旱胁迫[2];滇西高海拔云南松径向生长(Pinusyunnanensis)受夏季高温的限制,而低海拔受生长季降水的影响[3];庐山高海拔日本柳杉(Cryptomeriajaponica)生长受温度的限制,中低海拔径向生长受相对湿度的限制[4];太行山南麓高海拔栓皮栎(Quercusvariabilis)生长受最低温度的限制,低海拔生长受降水影响大[5]。也有研究显示,在不同海拔水平上树木生长对气候因素的响应较为一致[6],玉龙雪山不同海拔长苞冷杉(Abiesgeorgei)均受温度和降水的限制[7],不同海拔云南松(Pinusyunnanensis)对气候因素的响应模式相同[8]。不同气候区不同海拔树木径向生长对气候响应存在较大差异,因此需要开展不同地区不同海拔条件下树木生长对气候因素响应规律研究,为森林应对气候变化的经营管理提供科学依据,也为树木年轮学研究提供有益参考。
黄土高原位于黄河中游,具有世界上面积最大的黄土堆积,由于历史上人类社会对土地资源的加剧利用,该地区天然林和草原破坏严重,土地出现严重的荒漠化问题,成为世界上水土流失最严重的区域之一[9-11]。刺槐是黄土高原主要防护林树种,其根系发达,具根瘤,可固氮,耐瘠薄、耐干旱[12],具有固沟护坡、防风固沙、水文调节、碳固定及增汇等生态系统服务功能,对黄土高原自然环境改善和生态系统服务功能提升具有重要意义[13-14]。近年来,中国学者在黄土高原地区陆续进行了刺槐的树木年轮学研究,韦景树等[15]研究了羊圈沟小流域人工物种(刺槐、柠条)和自然物种(山杏、荆条)径向生长对气候变化的响应,发现人工物种对气候变化敏感,刺槐受水分胁迫严重。Keyimu等[16]研究了半湿润区永寿县和半干旱区神木县刺槐对气候变化的响应差异,发现半湿润地区刺槐对秋季温度响应敏感性增强,对水文要素的相关性减弱,而半干旱地区刺槐对秋季温度响应敏感性减弱,对水文要素的相关性增强。管崇帆等[17]研究了气候和竞争对刺槐径向生长的影响,发现温度和降水是影响刺槐生长的关键因素,高密度刺槐更易受干旱胁迫。以往研究多集中于水平梯度,不能代表海拔梯度上气候变化对刺槐生长的影响,因此需要研究不同海拔刺槐对气候变化的响应差异,以便了解气候变化背景下不同海拔刺槐的生长动态。
罗玉沟流域是黄土高原丘陵区第三副区的典型流域,为改善当地生态环境,1980s初该地区在梁顶、沟道等地貌部位上营造了大量的刺槐防护林[18]。2000年初随着退耕还林工程的继续实施,以刺槐为主的沟头防护林得到大规模实施,作为重要的水土保持植物措施在保持水土、涵养水源、调节区域小气候等方面发挥着重要作用[15]。由于早期营造刺槐林受气候变化和人为干扰的影响,该地区刺槐林出现林木生长不良、枯死、防护林防护功能明显下降等退化现象[19-20]。因此迫切需要研究刺槐径向生长过程及其与气候因素的关系,厘定限制不同海拔条件下刺槐生长的主要气候因素。研究以黄土高原丘陵区第三副区典型流域罗玉沟流域为研究区,以人工刺槐林为研究对象,利用树木年轮气候学方法研究不同海拔梯度刺槐径向生长对气候变化的响应,分析影响该地区刺槐生长的主要气候因素,为今后该区域低效刺槐林改造、恢复重建和经营管理提供科学依据。
1 研究区概况
罗玉沟流域位于甘肃省天水市区北部,是黄河水系藉河左岸的一条支流,属于黄土丘陵区第三副区典型流域[18]。罗玉沟流域海拔1200-1900 m,气候为暖温带大陆性季风气候(图1)。该地区降水年内分布不均,春冬干旱少雨,夏秋雨水集中,流域内多年平均降水量504.2 mm(1983-2017年,图2),汛期(6-9月)降水量占年降水量的62.3%;年平均气温11.6 ℃(1983-2017年,图2),无霜期184天,年平均风速1.3 m/s。土壤以山地灰褐土为主,是暖温带半干旱区森林草原气候条件下形成的土壤,占流域面积的91.7%。研究区内刺槐主要为人工纯林,一般选择2年生苗木造林,造林后实行封山育林。罗玉沟流域降水较为充沛,因人工刺槐林规模较大、山梁林业灌溉设施缺乏以及当地林业部门人力跟不上,因此,人工林种植后处于雨养状态,尚未有人工灌溉工作。林下灌木主要有胡枝子(Lespedezabicolor)、狼牙刺(Sophoramoorcroftiana)、刺五加(Eleutherococcussenticosus)等;草本以蔷薇科、菊科、禾本科植物居多。

图1 黄土高原罗玉沟流域刺槐年轮采样点分布图Fig.1 Location map of tree-ring sample site of Robinia pseudoacacia in Luoyugou watershed, Loess Plateau

图2 1983-2017年天水气象站温度和降水的变化趋势Fig.2 Temperature and precipitation changing trend of Tianshui Meteorological Station from 1983 to 2017
2 研究方法
2.1 样品的采集
本研究于2018年7-8月在罗玉沟流域选取受人类干扰较少的刺槐人工林,根据该流域的海拔、地形及刺槐林的空间分布,在北梁梁顶刺槐林中沿海拔布设20×20 m的样地,由低到高分别进行编号,共选取14个样地(图1)。根据海拔梯度将其划分为低海拔(1-4号样地,(1675±38) m)、中海拔(5-10号样地,(1758±23) m)以及高海拔(12-15号样地,(1836±24) m)。样地内刺槐是分批次种植的,存在很多树龄,且相互差异较大,在取样的过程中主要选取长势较好、胸径较大的刺槐5棵,用瑞典haglof 500 mm生长锥(5.15 mm)在胸径(约1.3 m)处沿东西、南北2个垂直方向钻取2-3个树芯,取好后将样芯放到塑料管内封存,同时记录刺槐的树号、胸径、树高、枝下高和冠幅等基本信息,共取树芯167个,其中低海拔采集53个,中海拔采集74个,高海拔采集40个。
2.2 样品的处理及年表建立
将树芯带回室内后,按Stokes等的方法对刺槐样芯进行预处理,风干后用白乳胶固定在特制的木槽内,由于树芯两端可能会翘起,还需用绳子加固[21]。待乳胶晾干后,将其切割成单个样芯,用400目、800目和1200目的干砂纸依次打磨,直至在显微镜下能清楚看到刺槐的年轮界线。利用精度为0.001 mm树木年轮测量分析仪Win DENDROTM(Regent instrument inc. Canada)进行树轮宽度参数的获取,用COFECHA程序[22]对定年和测量结果进行检验和校准,剔除与主序列相关性差和重复的样芯,最终通过交叉定年的样芯有115个,高中低海拔树芯分别为20个、55个和40个。
为去除树木随年龄增长和其他非气候因素导致的生长趋势,使用ARSTAN程序建立年表[23-24]。选择负指数函数方法进行不同海拔刺槐的生长拟合,去趋势后得到标准化年表(Standard chronology,STD)、自回归年表(Arstan chronology,ARS)和差值年表(Residual chronology,RES)。综合各年表的统计参数特征,本研究选用标准年表进行树木生长与气候因素间的相关分析。胸高断面积增量(Basal area increment,BAI)可更加准确的反映研究区内刺槐的生长量变化,具体公式如下:
式中,Rn是n年的树木半径;Rn-1是n-1年的树木半径。
2.3 气象数据来源
由于研究区气象站器测资料不完整,平均温度、最低温度、最高温度、降水量和相对湿度(1983-2017年)数据来源于中国气象数据网(http://data.cma.cn/),从距离取样点最近的天水气象站(105.75°E,34.58°N,海拔1141.6 m,距采样点4 km)获取。为更好的反映刺槐生长与水分的关系,研究亦选用了全球气候数据库(CRU TS 3.22 Global Climate Database,http://www.cru.uea.ac.uk/)中的帕默尔干旱指数(Palmer drought index,PDSI)[25]。从距采样点最近的CRU网格点(105.50°-106.00°E,34.50°-35.00°N)选取1983-2017年的PDSI格点数据。
2.4 数据处理及图表制作
通过计算不同海拔刺槐年表的统计特征值来评估年表的可靠性,使用Dendroclim 2002国际树木年轮程序对不同海拔树轮宽度指数与各气候数据进行相关分析[26]。考虑到树木生长可能受到前一年气候的影响,选用上一年6月到当年10月(共17个月)的气候数据进行相关性分析[27]。为进一步评估树木径向生长对主要气候因素的响应随时间变化的稳定性,使用DendroClim 2002程序中的Evolutionary and Moving Response and Correlation模块进行滑动相关分析,滑动窗口为20 a。使用Excel 2016和Origin 2018进行各图表的绘制。
3 研究结果
3.1 年表统计特征
由不同海拔刺槐标准年表的统计特征可知(表1),3个标准年表的平均敏感度在0.217-0.237之间,标准差在0.263-0.273之间,说明年表包含较多的气候信息。序列间相关性越高,说明刺槐受到的共同限制性越强[23],高海拔地区树间相关系数为0.390,中海拔为0.316,低海拔为0.404。高中低海拔的一阶自相关系数分别为0.525、0.487和0.512,说明上一年气候对树木生长有较大影响。标准年表的信噪比为6.402-7.446,表明各年表保留了丰富的气候信息。3个年表的样本总体代表性较高,超过样本总体代表性可接受的临界阈值0.85,说明各年表序列对总体的代表性高,适宜于树木年轮气候学分析[28]。

表1 罗玉沟流域采样点信息及标准年表统计特征Table 1 Site information, chronology statistics and results of common interval span analysis of standard tree-ring chronologies from the Luoyugou Watershed in the Loess Plateau of China
不同海拔标准年表的整体变化趋势有所差异(图3),中、低海拔刺槐分别在2000年和1997年出现最低值,2012年后出现下降趋势,而高海拔刺槐在2012年后出现升高趋势。低海拔和中海拔标准年表的相关性最高(0.576),中海拔和高海拔年表的相关性最低(-0.108)。

图3 罗玉沟流域不同海拔刺槐树轮宽度标准年表Fig.3 Ring-width standard chronologies of black locust at different elevations in Luoyugou Watershed of China
3.2 不同海拔刺槐树轮宽度和BAI年际变化特征
由图4可以看出高海拔刺槐树轮宽度年际变化波动较小,而低海拔刺槐树轮宽度年际变化波动最大。低海拔刺槐树轮宽度在1990年达到最高值(4.80 mm),1998年树轮宽度值最小(1.68 mm);中海拔刺槐树轮宽度在1992年达到最高值(3.95 mm),2002年树轮宽度值最低为2.17 mm;高海拔刺槐同样在1992年达到最高值(3.17 mm),1997年时最低值为1.76 mm。2000年后随着第二批刺槐林的加入树轮宽度值有一个高峰,低海拔刺槐在2007年达到最高值4.61 mm,中海拔刺槐在2007年为最高值4.94 mm,高海拔刺槐在2006年达到最高值4.61 mm。2013年之后,中低海拔刺槐树轮宽度呈下降趋势,而高海拔刺槐呈上升趋势。
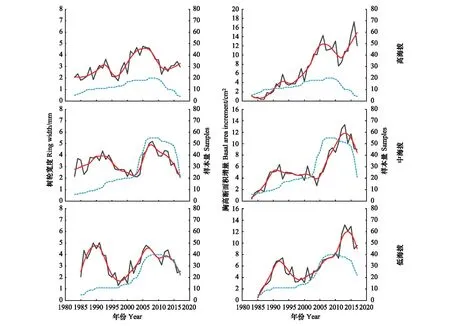
图4 罗玉沟流域不同海拔刺槐树轮宽度和BAI年际变化Fig.4 Interannual variation of black locust ring width and BAI at different elevations in Luoyugou Watershed红色线条为11年滑动平均序列,蓝色虚线为样本量
高中低海拔刺槐胸高断面积增量(BAI)总体均呈上升趋势(图4)。低海拔刺槐在1985-1992年期间呈上升趋势(0.80 cm2/a),1993-1999年呈下降趋势(-0.42 cm2/a),2000-2014年一直呈上升趋势(0.55 cm2/a),2015年后BAI呈下降趋势。中海拔刺槐在1983-1991年期间呈上升趋势(0.54 cm2/a),1992-2006年期间生长平稳,平均生长速率为(4.64±0.88) cm2/a,2007-2013年呈上升趋势(0.72 cm2/a),2014年后BAI呈下降趋势。高海拔刺槐在1983-2006年期间呈上升趋势(0.47 cm2/a),2007-2011年期间生长略有下降,2012年后呈上升趋势(0.96 cm2/a)。
3.3 不同海拔刺槐径向生长与气候因素的相关性
中低海拔刺槐径向生长与温度因素多为负相关关系,而高海拔刺槐径向生长与温度因素基本呈正相关(图5)。中低海拔刺槐径向生长均与上一年生长季和当年生长季温度呈显著负相关,而高海拔刺槐主要与上一年生长季、休眠期和当年生长季温度呈显著正相关(图5)。具体来看:(1)低海拔刺槐与上一年7-9月以及当年6-7月均温、最高温和最低温呈显著负相关,其中对均温的响应更显著;(2)中海拔刺槐与上一年6月、当年1、2、7、9月均温和最高温呈显著负相关,其中对最高温的响应更显著;(3)高海拔刺槐与上一年6月、当年3、4月和6月均温、最高温和最低温呈显著正相关,还与当年2、9月最低温呈显著正相关,其中最低温限制作用最大。
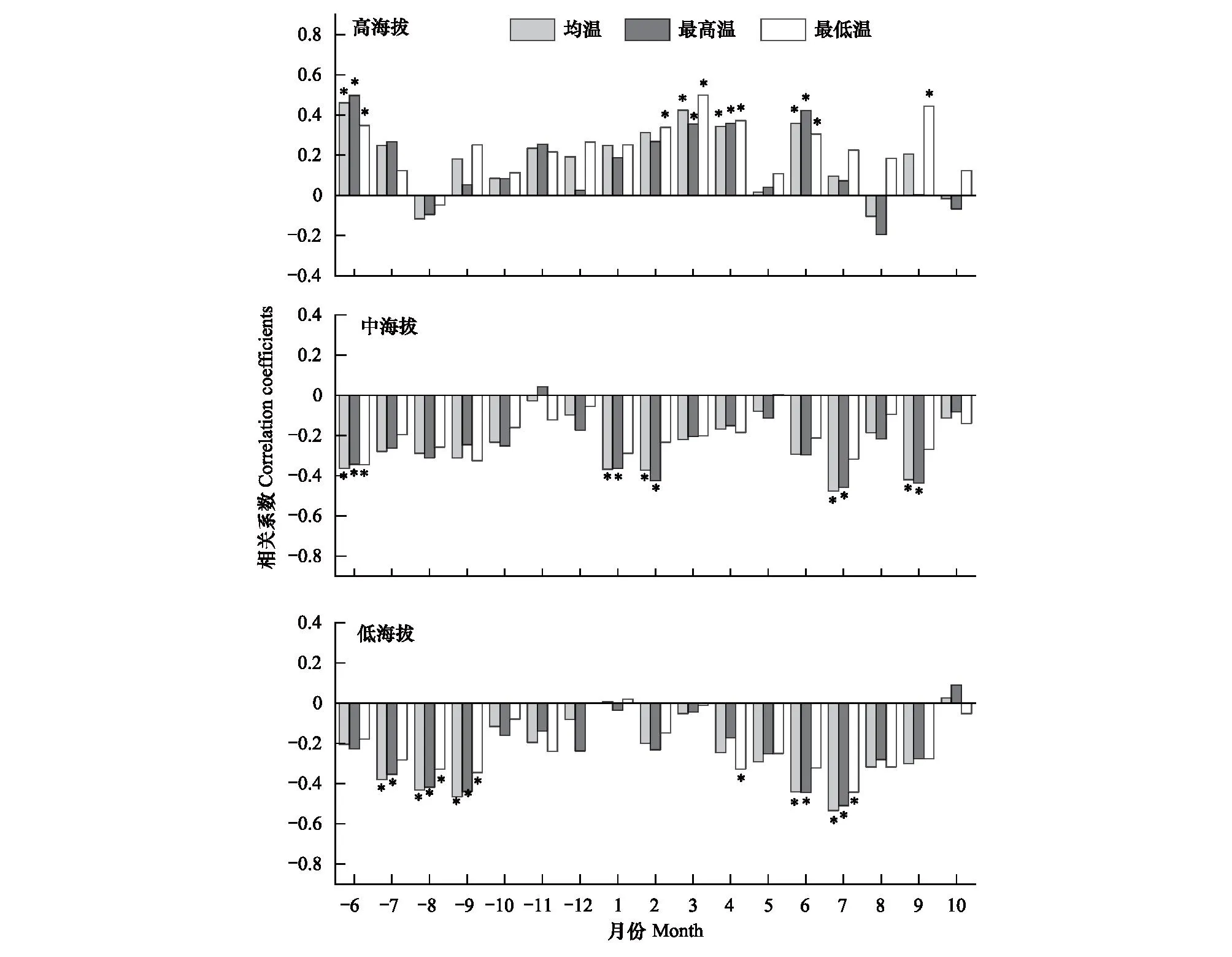
图5 不同海拔刺槐标准年表与温度因素的相关关系Fig.5 Correlation between standard chronology and temperature factors of black locust at different elevations*代表P<0.05,月份前的“-”表示上一年,如-6表示上一年6月份
中低海拔刺槐径向生长与降水和相对湿度多呈正相关关系,而高海拔刺槐径向生长与相对湿度的负相关关系较大(图6)。低海拔刺槐生长与上一年生长季和当年生长季降水、相对湿度呈显著正相关,中海拔刺槐与当年生长季降水、相对湿度呈显著正相关,而高海拔刺槐生长与上一年生长季相对湿度呈显著负相关(图6)。具体来看:(1)低海拔刺槐生长与上一年8月、当年6、7月降水和相对湿度呈显著正相关,还与上一年7、9月、当年8、9月相对湿度呈显著正相关;(2)中海拔刺槐与当年6月降水呈显著正相关,与当年2、7-9月相对湿度呈显著正相关;(3)高海拔刺槐仅与上一年6、7月相对湿度呈显著负相关,与降水相关性较小。

图6 不同海拔刺槐标准年表与降水、相对湿度和PDSI的相关关系Fig.6 Correlation between standard chronology, precipitation, relative humidity and PDSI of black locust at different elevations*代表P<0.05,月份前的“-”表示上一年,如-6表示上一年6月份
中低海拔刺槐径向生长与PDSI多呈正相关关系,而高海拔刺槐径向生长与PDSI多呈负相关(图6)。低海拔刺槐径向生长与上一年6-当年10月PDSI均呈显著正相关,中海拔刺槐与上一年6月和当年2-10月PDSI呈显著正相关,而高海拔刺槐径向生长与上一年6、7月PDSI呈显著负相关。
3.4 不同海拔刺槐径向生长对气候响应的稳定性评价
不同海拔刺槐径向生长对气候变化均产生不同程度的分异现象(图7)。低海拔刺槐生长与当年6-9月平均温度、最高温度、相对湿度和PDSI的响应一直保持稳定的显著相关水平,未发生分异现象;与当年6-9月最低温度出现负相关明显减弱趋势(P<0.05),发生分异现象。中海拔刺槐生长与当年6-9月平均温度、最高温度的响应保持稳定的负相关,与当年6-9月降水量的响应保持稳定的正相关;而与当年6-9月最低温度出现负相关明显减弱趋势(P<0.05),1990年后对相对湿度和PDSI的敏感性逐渐减弱。高海拔刺槐生长与当年6-9月PDSI维持较为稳定的关系,1991年后对温度要素的敏感性略有减弱,1994年后对降水量和相对湿度的敏感性明显减弱,出现分异响应。

图7 不同海拔刺槐树轮宽度与当年6-9月气象因子的滑动相关分析(20 a)Fig.7 The 20-year window moving correlations of black locust chronologies and climate factors from current June to September at different elevations
4 讨论
4.1 不同海拔刺槐年表特征及径向生长差异
年表特征值可以反映不同海拔刺槐径向生长与气候因素的相关程度。低海拔刺槐年表的标准差、平均敏感度和树间相关系数较中、高海拔高,说明低海拔年表质量最好,对气候响应的敏感性较高。本研究中随海拔升高,信噪比和样本总体代表性随之减小,说明刺槐径向生长对气候的响应敏感性降低,此结果与白天军等[4]研究结果一致。罗玉沟流域属于半干旱地区,流域内树木生长受水分限制作用强,随着海拔升高,降水量增加,高海拔地区降雨总量大于低海拔地区,促使高海拔地区刺槐生长受水分限制作用降低,高海拔地区对气候敏感性也随之降低[29]。低海拔刺槐径向生长主要受限于土壤水分可利用程度(图6)。
不同海拔刺槐树轮宽度均在1992年和2007年左右出现最高值,在1998年左右出现最低值(图4),说明大尺度气候变化对不同海拔刺槐径向生长影响大,在极端年份对树木的生长起决定作用。2013年后,中、低海拔刺槐径向生长呈下降趋势,而高海拔刺槐生长呈上升趋势。在气候暖干化背景下,中低海拔区域将不再适合刺槐的生长,若干旱时间和强度持续上升,会导致该区域生态服务功能下降,提高火灾和病虫害的发生机率,后期可能出现枯树梢或枯死现象,导致人工刺槐林大面积衰退,生产力下降。气温升高会促进高海拔区域树木生长,将导致刺槐林林线上移[15]。
4.2 不同海拔刺槐径向生长对气候因素的响应差异
随海拔的升高,刺槐径向生长与温度的显著负相关转变为显著正相关(图5)。上一年生长季和当年生长季温度过高会限制中低海拔刺槐的生长,与韦景树等[15]对黄土高原羊圈沟流域刺槐的研究结果一致。温度对刺槐的径向生长存在“滞后效应”[30],上一年9月是刺槐生长季末期,温度过高会导致植物蒸腾作用加强,土壤水分的蒸发增大,影响刺槐碳水化合物的合成,并且有可能消耗本身的营养物质,导致下一年径向生长减缓[31]。当年生长季高温会导致土壤干旱,形成的水分胁迫降低了树木水势,增大叶肉细胞对水分的吸附力,限制叶内水分运动,从而降低植物的光合作用[32-33]。上一年生长季、休眠期和当年生长季温度对高海拔刺槐有促进作用,与玉龙雪山高海拔丽江云杉和长苞冷杉、关帝山林区华北落叶松与温度呈正相关的结果一致[7,34-36]。休眠期最低温度升高对高海拔刺槐生长有促进作用,休眠期温度升高可以有效防止低温对树木根系的伤害,冬季降雪有利于使土壤保持较好墒情,可以增加树木生长前期的水分储备,生长季前有效积温足够并且水分充足以便刺槐进入萌芽期,生长季时间增长有助于刺槐形成宽轮[34]。夏季高温对高海拔树木生长具有重要促进作用,夏季是树木生长旺盛时期,降水充足情况下适宜的高温有助于树木的光合作用,有效积累碳水化合物促进树木生长[37]。
罗玉沟流域年降水量存在明显空间差异,流域沟口到沟头,随着海拔上升多年平均降水量从460 mm上升至656 mm,降水量增加趋势明显,增加了42.6%(图8)[38]。流域中下游地区降水较上游明显偏少,因此,流域中低海拔刺槐生长受干旱胁迫限制,其中低海拔受干旱胁迫尤为显著(图8),而流域上游地区降水为600 mm,降水不是流域上游地区刺槐林生长的主导限制性因素(图8),而高海拔温度低,更多受温度影响(图5)。当年生长季降水促进中低海拔刺槐的生长,树木进入生长季后,生长旺盛需水量大,有效的降水可以促进光合作用产物的积累加速植物后期生长[39-40]。高海拔刺槐生长与降水相关性不高,该地区海拔升高降水量充足,温度是主要的限制因素。

图8 罗玉沟流域降水随海拔的变化趋势(1986-2004)[38]Fig.8 The variation trend of precipitation with elevation of Luoyugou Watershed, China (1986-2004)
PDSI反映土壤中水分的可利用程度,它直接影响树木形成层活动,影响木质部的生成[41]。中低海拔刺槐与多月份PDSI呈显著正相关,尤其是上一年生长季和当年生长季,说明中低海拔刺槐受水分胁迫的深刻影响,该区域刺槐遭受高温导致蒸发量增大,而降水量较少,极易遭遇干旱胁迫。在高海拔地区温度偏低,但降水量大(图7),较适宜刺槐生长。
4.3 不同海拔刺槐径向生长对气候变化的响应稳定性
由滑动相关分析发现,中低海拔地区刺槐对低温响应的敏感性明显降低,低海拔刺槐对水文要素(降水、相对湿度和PDSI)的敏感性维持稳定的显著正相关水平,而中海拔刺槐对水文要素的敏感性略有降低。高海拔刺槐对温度因素响应敏感性较为稳定,对降水和相对湿度的响应敏感性减弱。20世纪80年代以来,罗玉沟流域经历了快速升温降水减少,高温低降水导致树木受干旱胁迫严重,低海拔地区与6-9月PDSI维持稳定显著正相关,水分是限制低海拔刺槐未来生长的主要气候因素。此结果与Keyimu等报道的黄土高原半干旱地区刺槐对温度要素的敏感性下降,与水文要素的敏感性上升结果一致[16]。祁连山东部半干旱地区青海云杉与温度要素的负相关性逐渐减弱,与水文要素的正相关性逐渐增强,土壤水分条件对青海云杉的限制作用逐渐增强,本研究结果与其一致[42]。中低海拔刺槐对最低温度的响应敏感性减弱,与流域内气温迅速上升变化一致,生长季中最低温度对刺槐生长的限制作用逐渐消失。高海拔地区对温度、降水和相对湿度的敏感性均降低,说明高海拔地区温度升高对高海拔地区刺槐生长是有利的[43]。
5 结论
本研究表明,位于黄土高原丘陵区第三副区的甘肃天水罗玉沟流域,中、低海拔刺槐生长受上一年生长季和当年生长降水的限制,低海拔刺槐受水分限制作用更强;高海拔刺槐受上一年生长季、休眠期和当年生长季温度的限制,其中最低温对其限制作用更强,最低温度升高对高海拔刺槐有促进作用。中、低海拔刺槐受干旱胁迫影响严重,尤其低海拔地区与上一年生长季、休眠期和当年生长季PDSI均呈显著正相关,低海拔刺槐受干旱胁迫较中海拔地区高。在全球气候变暖的趋势下,罗玉沟流域中、低海拔刺槐林径向生长受干旱胁迫将更加严重,遇到严重干旱时将出现生长缓慢,甚至枯树梢、枯死等衰退现象。
